Аммониотелические животные
В книге проанализированы различные подходы и взгляды на структуру биосферы преимущественно на примере биогеографии и экологии морского бентоса.
Рассмотрены основные аспекты биологии обитателей дна водоемов: факторы абиотичеокой среды и их влияние на гидробионтов, особенности аутоэкологии (питания, размножения и др.), донные сообщества и трофическая зональность, синэкология и биогеография. Вопросы биогеографии и экологии рассматривают в литературе с самых разных позиций. В данной монографии они рассмотрены с позиций биологии, и, чтобы подчеркнуть это, оба слова написаны через дефис: био-география и био-экология.
Детально разобраны некоторые сообщества (макрофитов, коралловых рифов, глубоководных гидротерм). Подробно представлена структура, динамика, взаимодействие и эволюция экосистем — вопросы обычно слабо или вовсе не затрагиваемые в монографиях и учебниках по экологии и биогеографии. На множестве примеров показана общность экологических законов для экосистем суши, пресных и морских водоемов.
ну из основных своих задач автор видит в популяризации идей о существовании минимальной структурно-функциональной самовоспроизводящейся единицы биосферы — ценотической системы. Ценотическая система обладает своей собственной историей, судьбой, эволюцией. Отдельные сообщества, образующие конкретную ценотическую систему, есть лишь ее разные пространственно-временные стадии. Наблюдающаяся в каждый конкретный момент на территории ценотической системы мозаика сообществ обусловлена (1) асинхронностью в разных местах сукцессии восстановления после разнообразных нарушений и (2) изначальной гетерогенностью среды, которая приводит к тому, что в разных местах ареала ценотической системы идут сукцессии разных сукцессионных рядов. При обсуждении вопросов континуальности-дискретности биосферы масштаб рассмотрения имеет принципиальное значение. Существенно влияют на результат и использованные методы и подходы, многие из которых даже теоретически не способны обнаружить реально существующую дискретность.
Немало внимания в книге уделено вопросам биогеографии. По мнению автора, так называют две принципиально разные науки, отличающиеся объектом, методами и теорией: географическую и биологическую биогеографии. Обоснование такого взгляда дано в главе 5.
Вопросы дискретности биосферы рассмотрены с двух диаметрально противоположных позиций: континуализма и структурализма, причем настоящая книга — первая в отечественной литературе краткая сводка по теории структурализма. Дан анализ методов биогеографии. Подробно разобраны причины эндемизма, скорость образования новых таксонов, образование, развитие и взаимодействие ценотических систем.
Книга снабжена предметным указателем и словарем.
Предназначена для студентов, аспирантов, преподавателей вузов биологических специальностей и научных сотрудников, изучающих жизнь в морях и океанах, а также для широкого круга лиц, интересующихся вопросами биогеографии и экологии….
Источник: www.educationspb.ru
Виды конечных продуктов азотистого обмена у животных разных групп. В чем причины, определяющие их различия При расщеплении белков, нуклеиновых кислот и других азотсодержащих соединений образуются токсичные вещества — аммиак, мочевина и мочевая кислота, токсический эффект которых соответственно снижается в приведенном ряду. В зависимости от того, в какой из этих трех форм преимущественно выделяется азот, животных подразделяют на три группы: аммониотелические урикотелические (выделяющие мочевую кислоту).
Форма выделения продуктов азотистого обмена тесно связана с условиями жизни животного и обеспеченностью водой. Аммиак весьма токсичен даже в малых концентрациях. Благодаря хорошей растворимости и небольшому молекулярному весу, он легко диффундирует через любую поверхность, соприкасающуюся с водой. Аммиак является конечным продуктом азотистого обмена у водных беспозвоночных, костных рыб, личинок и постоянно живущих в воде земноводных.
Наземные животные ограничены в воде: чтобы избежать накопления аммиака в тканях и жидкостях тела, они должны преобразовать его в конечные продукты, нетоксичные для организма. Наземные ресничные черви, земноводные, млекопитающие выделяют мочевину.
Низкая растворимость мочевой кислоты, выпадение ее в осадок делает ее осмотически неактивной. Для ее выведения из организма вода практически не нужна. Урикотелия в основном характерна для животных, освоивших наземную, в том числе и засушливую, среду (наземные насекомые, чешуйчатые пресмыкающиеся, птицы).
Водно-солевой обмен рыб
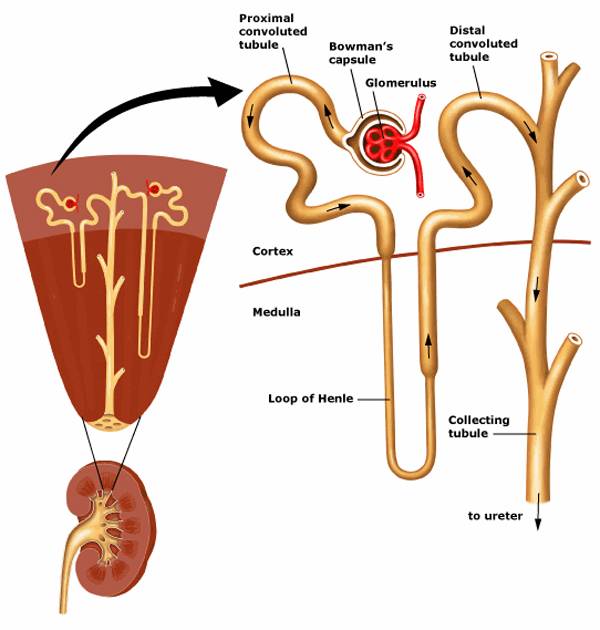
Почки рыб выводят аммиак, соли, воду; почки наземных позвоночных — мочевину, мочевую кислоту, соли, воду.Выделительная система рыб служит для выведения из организма продуктов обмена и обеспечения его водно-солевого состава. Она включает:
Основную массу туловищной почки заполняют нефроны. Нефрон состоит из:
1) мальпигиева тельца (клубочек капиллярных сосудов, заключенный в боуменову капсулу);
2) выделительного канальца.
Артериальная кровь по почечным артериям поступает в сосудистые клубочки, где фильтруется и образуется первичная моча. В средней части выделительных канальцев происходит обратное всасывание полезных для организма веществ (сахара, витамины, аминокислоты, вода) и образуется вторичная, или окончательная, моча. У хрящевых рыб основным компонентом мочи является мочевина, у костистых — аммиак (аммиак намного токсичнее мочевины).
Выделение продуктов распада тесно связано с водно-солевым обменом рыб. У морских и пресноводных рыб эти процессы протекают различно.
When a fish eats protein, like the peel of an orange part of it goes unused and becomes waste.
Dave McShaffrey, professor of biology at Marietta College in Ohio, explains on the college website, “When proteins are converted to carbohydrates to provide energy, the amino group is removed and must be dealt with.” In saltwater fish, this nitrogen-rich waste is usually converted to either ammonia or urea, which is one of the main excretory products of saltwater fish. Ammonia is easier to produce, but urea is less toxic, requires less water and gets rid of twice as much nitrogen. Theword “urine” is related to “urea.”
Морские хрящевые рыбы живут в изотоничной среде (т.е. осмотическое давление крови и тканевых жидкостей равно давлению окружающей среды). У них изотоничность внутренней и внешней среды обеспечивается за счет удержания в крови и тканевых жидкостях мочевины и солей (концентрация мочевины в крови у них достигает 2,6%). Через почки наружу у них выводятся лишь излишки мочевины, солей и воды, количество выделяемой мочи невелико (2-50 мл на 1 кг массы тела в сутки). У морских хрящевых рыб для выведения избытка солей сформировалась особая ректальная железа, открывающаяся в прямую кишку.
Все пресноводные рыбы живут в гипотонической среде (т.е. осмотическое давление крови и тканевых жидкостей выше, чем в окружающей среде), поэтому вода постоянно проникает в организм через кожу, жабры, с пищей. Чтобы избежать обводнения пресноводные рыбы имеют хорошо развитый фильтрационный аппарат почек, что позволяет им выделять большое количество мочи (50-300 л на 1 кг массы тела в сутки). Потеря солей с мочой компенсируется активной их реабсорбцией в почечных канальцах и поглощением солей жабрами из воды, часть солей поступает с пищей.
Морские костистые рыбы живут в гипертонической среде (т.е. осмотическое давление крови и тканевых жидкостей ниже, чем в окружающей среде), поэтому вода выходит из организма через кожу, жабры, с мочой и фекалиями. Во избежание иссушения они пьют соленую воду (от 40 до 200 мл на 1 кг массы в сутки), которая из кишечника всасывается в кровь. У морских костистых рыб уменьшается число клубочков в почках, а у некоторых рыб исчезают совсем (морская игла, морской черт). Таким образом, почки выводят небольшое количество мочи (0,5—20 мл на 1 кг массы тела в сутки).
Проходные рыбы при переходе из одной среды в другую могут изменять способ осморегуляции: в морской среде она осуществляется как у морских рыб, а в пресной — как у пресноводных. Такие адаптации водно-солевого обмена позволили костистым рыбам широко освоить пресные и соленые водоемы.
Адаптации наземных животных к выделению веществ
According to New World Encyclopedia, reptiles use two small kidneys as tools for excretion. The kidneys serve to filter the nitrogen from the animal’s bloodstream, then turn it into waste. The nitrogen then exits the body in dry form as uric acid crystals along with the feces. According to Stanford University, the kidneys in a bird also function as a means to remove nitrogen from the blood. The white substance found in bird droppings is actually uric acid, which is not water soluble. In both reptiles and birds, eliminating the nitrogen requires that the body exerts a great deal of energy. Both species are able to efficiently remove the nitrogen while losing very little water in the waste product.
Адаптации пустынных животных к выделению веществ
Обитатели полупустынных биотопов большую часть воды получают, поедая сочные части суккулентных растений.
кожно-легочные потери воды минимальны. Так, при температуре 20°С они достигают у относительно влаголюбивого вида — гребенщиковой песчанки 170 см3, а у сухолюбивой большой песчанки — только 50 см3 на 1 кг массы за 1 ч. Настоящие пустынные млекопитающие способны поедать почти сухие корма и практически не пить в течение всей жизни, удовлетворяя свои потребности лишь за счет образующейся в организме метаболической воды. Верблюды в кормные и влажные сезоны запасают жир, расходуемый в малокормное и сухое время — при этом образуется некоторое количество воды; наконец, во время отдыха и сна они снижают температуру тела, что также сокращает расход воды.
Desert Animals
Coping with water loss is a particular problem for animals that live in dry conditions. Some, like the camel, have developed great tolerance for dehydration. For example, under some conditions, camels can withstand the loss of one third of their body mass as water.
ey can also survive wide daily changes in temperature. This means they do not have to use large quantities of water in sweat to cool the body by evaporation.Smaller animals are more able than large ones to avoid extremes of temperature or dry conditions by resting in sheltered more humid situations during the day and being active only at night.The kangaroo rat is able to survive without access to any drinking water at all because it does not sweat and produces extremely concentrated urine. Water from its food and from chemical processes is sufficient to supply all its requirements.

Какой из нефронов принадлежит верблюду, а какой – рептилии? Почему вы сделали такой выбор?
Fresh Water Fish
Although the skin of fish is more or less waterproof, the gills are very porous. The body fluids of fish that live in fresh water have a higher concentration of dissolved substances than the water in which they swim. In other words the body fluids of fresh water fish are hypertonic to the water (see chapter 3). Water therefore flows into the body by osmosis. To stop the body fluids being constantly diluted fresh water fish produce large quantities of dilute urine.
Marine Fish
Marine fish like the sharks and dogfish have body fluids that have the same concentration of dissolved substances as the water (isotonic) have little problem with water balance. However, marine bony fish like red cod, snapper and sole, have body fluids with a lower concentration of dissolved substances than seawater (they are hypotonic to seawater). This means that water tends to flow out of their bodies by osmosis. To make up this fluid loss they drink seawater and get rid of the excess salt by excreting it from the gills.
Marine Birds
Marine birds that eat marine fish take in large quantities of salt and some only have access to seawater for drinking. Bird’s kidneys are unable to produce very concentrated urine, so they have developed a salt gland. This excretes a concentrated salt solution into the nose to get rid of the excess salt.
2. Using the words/phrases in the list below fill in the blanks in the following statements.
| cortex | amino acids | renal | glucose | water reabsorption | large proteins |
| bowman’s capsule | diabetes mellitus | secreted | antidiuretic hormone (ADH) | blood cells |
| glomerulus | concentration of the urine | medulla | nephron |
a) Blood enters the kidney via the ……………………. artery.
b) When cut across the kidney is seen to consist of two regions, the outer………….. and the inner…………..
c) Another word for the kidney tubule is the………………………….
d) Filtration of the blood occurs in the…………………………
e) The filtered fluid (filtrate) enters the………………………..
f) The filtrate entering the e) above is similar to blood but does not contain……………… or………………..
g) As the fluid passes along the first coiled part of the kidney tubule……………… and……………….. are removed.
h) The main function of the loop of Henle is…………………………………………………….
i) Hydrogen and potassium ions are………………………… into the second coiled part of the tubule.
j) The main function of the collecting tube is…………………………………………..
k) The hormone……………………………….. is responsible for controlling water reabsorption in the collecting tube.
l) When the pancreas secretes inadequate amounts of the hormone insulin the condition known as…………………………. results. This is most easily diagnosed by testing for………………………….. in the urine.
Источник: infourok.ru
|
1 Общие представления о процессе выделения. Секреция. Экскреция. Выделение (экскреция) – освобождение о-ма от конечных продуктов обмена в-в, избытка воды, солей, БАВ, чужеродных и токсичных соединений (поступивших с пищей или обр-ся при метаболизме). Секреция – образование и выведение в-в из клетки во внешнюю среду. Экскреция – частный случай секреции. Ф-ии выделит системы (значение процесса экскреции): 1 – выведение продуктов хим р-ий, которые могут сдвигать хим равновесие (D): А+В=С+D. 2 – выведение токсических продуктов, которые образуются в процессе метаболических реакций (напр продукт дезаминирования – аммиак). 3 – выведение излишков солей и ионов (Na, K, Ca, Mg, Fe, Cl, фосфаты, карбонаты). 4 – регуляция содержания воды в о-ме. 5 – поддержание водно-солевого равновесия в о-ме (осморегуляция). 6 – поддержание КЩР (важную роль играют бикарбонаты). Органы выделения позвоночных: почки, кожа (вода, соли, мочевина), легкие (вода, СО2), печень (желчные пигменты, образуется аммиак, из которого образуются мочевина).
Биология
1 Общие представления о процессе выделения. Секреция. Экскреция 2 Взаимосвязь процесса экскреции и осморегуляции 3 Продукты, подлежащие экскреции 4 Аммонио-, урео-, урекотелия 5 Филогенетический обзор органов выделения беспозвоночных 6 Роль кожи, жабр, легких и печени в процессах выделения и осморегуляции у позвоночных 7 Типы почек у позвоночных. Взаимо-связь выделит-х и половых протоков 8 Особенности выделения и осморегуля-ции в разных гр позвоночных |
2 Взаимосвязь процесса экскреции и осморегуляции. Выделение (экскреция) – освобождение о-ма от конечных продуктов обмена в-в, избытка воды, солей, БАВ, чужеродных и токсичных соединений (поступивших с пищей или обр-ся при метаболизме). Осморегуляция – поддержание водно-солевого равновесия. Связь процесса экскреции с процессом осморегуляции заключается в обеспечении постоянства ионного сотава внутренней среды о-ма и поддержание КЩР. В зависимости от способности поддерживать постоянным свое осмотическое давление при / осмотического давления окружающей среды, живые организмы делятся на осморегуляторов (способны) и осмоконформеров (ОД меняется вслед за / ОД внешней среды). Осмоконформеры – при / осмоляльности внешней среды осмотическое давление крови и внеклеточных жидкостей меняется. Вследствие того что плазматическая мембрана клеток проницаема для воды, происходит / объема клеток / их функций – выработался ряд мех-мов противодействия этому процессу: При / осмоляльности морской воды, а и гемолимфы, клетки сморщиваются, в них / концентрация аминокислот, ионов Na+ и Сl–, из внеклеточной среды входит вода и восстанавливается исходный объем. При осмоляльности морской воды клетки набухают, в их цитоплазме концентрация аминокислот, натриевый насос удаляет избыток ионов Na+ — исходный объем клетки восстанавливается. Осморегуляторы могут иметь меньшее осмотическое давление крови, чем в окружающей водной среде (миноги и костистые рыбы в морской воде), или большее (все пресноводные организмы и хрящевые рыбы в морской воде). Гипоосмотическая регуляция (на примере морских костистых рыб): В море они теряют воду через жаберную поверхность (обезвоживаются). Потеря воды происходит по осмотическому градиенту, т.к морская вода имеет большее осмотическое давление, чем кровь. Для возмещения дефицита воды рыбы пьют морскую воду, она всасывается вместе с солями Na+ и К+ в кровь. В жабрах имеются хлоридные клетки, которые удаляют из крови в морскую воду соли Na+ без воды – опреснение жидкостей внутренней среды. Часть солей Са2+ и Mg2+ удаляется почками и кишечником. Гиперосмотическая регуляция: У пресноводных животных часть солей сорбируется из внешней среды специальными клетками в жабрах или покровах тела, часть поступает с пищей, а избыток воды удаляется почкой. У наземных организмов потребности в воде и солях обеспечиваются пищей и потреблением воды. |
3 Продукты, подлежащие экскреции. Выделение (экскреция) – освобождение о-ма от конечных продуктов обмена в-в, избытка воды, солей, БАВ, чужеродных и токсичных соединений (поступивших с пищей или обр-ся при метаболизме). Продукты, выводимые из о-ма: 1 – О2 (у растений), 2 – СО2 – образуется в о-ме в результате процессов энергетического обмена, выводится путем диффузии и через легкие. 3 – соли, 4 – соли желчных кислот – образуются в печени, являются составной частью желчи. 5 – желчные пигменты – продукты расщепления Hb (в о-ме не используются). (1). 6 – азотистые продукты обмена – образуются в результате расщепления амино- и нуклеиновых кислот. В зависимости от того в каком виде выделяются продукты азотистого обмена, животных можно разделить на: 1 – аммониотелические (пресноводные и морские беспозвоночные, костистые рыбы, личинки и постоянно живущие в воде пресноводные) – продукты азотистого обмена выводятся в виде аммиака. 2 – уреотелические (наземная планария, хрящевые рыбы, взрослые земноводные, млекопитающие) – мочевина. 3 – урикотелические (наземные брюхоногие моллюски, наземные насекомые, пресмыкающиеся, птицы) – мочевая кислота. 4 – гуанотелические (скорпионы, пауки) – гуанин. У форм животных, нуждающихся в экономном расходовании воды выделяется мочевая кислота и ее слабо растворимые соли. Токсический эффект продуктов азотистого обмена падает в ряду: аммиак > мочевина > мочевая кислота.
|
4 Аммонио-, урео-, урекотелия. Выделение (экскреция) – освобождение о-ма от конечных продуктов обмена в-в, избытка воды, солей, БАВ, чужеродных и токсичных соединений (поступивших с пищей или обр-ся при метаболизме). Азотистые продукты обмена образуются в результате расщепления амино- и нуклеиновых кислот. В зависимости от того в каком виде выделяются продукты азотистого обмена, животных можно разделить на: 1 – аммониотелические (пресноводные и морские беспозвоночные, костистые рыбы, личинки и постоянно живущие в воде пресноводные) – продукты азотистого обмена выводятся в виде аммиака. 2 – уреотелические (наземная планария, хрящевые рыбы, взрослые земноводные, млекопитающие) – мочевина. 3 – урикотелические (наземные брюхоногие моллюски, наземные насекомые, пресмыкающиеся, птицы) – мочевая кислота. 4 – гуанотелические (скорпионы, пауки) – гуанин. Характер и соотношение конечных продуктов азотистого обмена имеют приспособительное значение – у форм животных, нуждающихся в экономном расходовании воды выделяется мочевая кислота и ее слабо растворимые соли. Токсический эффект продуктов азотистого обмена падает в ряду: аммиак > мочевина > мочевая кислота. У человека и большинства позвоночных образующийся при распаде белков азот экскретируется в основном в виде мочевины. Она свободно фильтруется, но легко диффундирует (в проксимальном канальце около 1/3 мочевины возвращается в кровь). В дистальной части нефрона и в концевом отделе собирательной трубочки диффузионная проницаемость для мочевины низка, но здесь она может реабсорбироваться по механизму следования за растворителем. Поэтому, если в дистальных отделах всасывается большое количество воды и образуется моча с высокой осмотической концентрацией, вместе с водой возвращается в кровь еще до трети отфильтрованной мочевины. В случае водного диуреза всасывание воды в дистальном канальце прекращается, и мочевины соответственно выводится больше. Таким образом, ее экскреция зависит от диуреза.
|
|
5 Филогенетический обзор органов выделения беспозвоночных. Осморегуляция, экскреция воды у пресноводных простейших, а также выделение жидкости у некоторых морских форм обеспечиваются сократительной вакуолью. У амебы она окружена мембраной, снаружи от которой располагается зона мелких вакуолей и далее находятся митохондрии. В пресной воде содержимое сократительной вакуоли у амебы имеет меньшее осмотическое давление, чем цитоплазмы. Эта вакуоль обеспечивает экскрецию воды. Сократительная вакуоль имеется обычно у пресноводных форм, а у эндопаразитических может отсутствовать. Губки и кишечнополостные лишены специальных выделительных органов. Выделение с помощью протонефридия осуществляется у плоских червей, лишенных целома. Протонефридий представляет собой канадец, проксимальная часть которого состоит из терминальной клетки с ресничками. Биение ресничек создает отрицательное давление в просвете нефридиального канальца, благодаря чему в него засасывается жидкость. В просвете канальца реабсорбируются ионы натрия и вода. При наличии целомической полости выделительная функция осуществляется метанефридиями и целомодуктами. Метанефридии функционируют у аннелид, измененные целомодукты имеются у моллюсков, членистоногих. Важную роль в экскреции у членистоногих выполняют выросты кишечника из которых хорошо исследованы мальпигиевы канальцы. Кольчатые черви обладают полостью тела, в которой находится целомическая жидкость, и замкнутой системой кровообращения. Нефридиостом метанефридия открывается в целомическую полость, из которой жидкость поступает в метанефридий, движется по канальцу (он состоит из нескольких отделов — узкого, ресничного и широкого) и поступает в мочевой пузырь, из которого содержимое через нефридиопору выделяется наружу. В канальцах реабсорбируются соли, в результате образуется гипотоническая моча. Перикардиальная полость у моллюсков является остатком целома. В нее через стенку желудочка сердца фильтруется жидкость, которая по реноперикардиальному каналу поступает в почечный мешок, движется по первичному и вторичному мочеточникам и выделяется наружу. Эпителиальные клетки почечного мешка имеют многочисленные микроворсинки на поверхности, обращенной в просвет мешка. Конечная моча морских моллюсков изоосмотична гемолимфе, у пресноводных — гипотонична ей.
|
6 Роль кожи, жабр, легких и печени в процессах выделения и осморегуляции у позвоночных. Выделение (экскреция) – освобождение о-ма от конечных продуктов обмена в-в, избытка воды, солей, БАВ, чужеродных и токсичных соединений (поступивших с пищей или обр-ся при метаболизме). Секреция – образование и выведение в-в из клетки во внешнюю среду. Экскреция – частный случай секреции. Органы выделения позвоночных: почки, кожа (вода, соли, мочевина), легкие (вода, СО2), печень (желчные пигменты, образуется аммиак, из которого образуются мочевина). Связь процесса экскреции с процессом осморегуляции заключается в обеспечении постоянства ионного сотава внутренней среды о-ма и поддержание КЩР. В зависимости от способности поддерживать постоянным свое осмотическое давление при / осмотического давления окружающей среды, живые организмы делятся на осморегуляторов (способны) и осмоконформеров (ОД меняется вслед за / ОД внешней среды). Осмоконформеры – при / осмоляльности внешней среды осмотическое давление крови и внеклеточных жидкостей меняется. Осморегуляторы могут иметь меньшее осмотическое давление крови, чем в окружающей водной среде (миноги и костистые рыбы в морской воде), или большее (все пресноводные организмы и хрящевые рыбы в морской воде). Роль жабр в процессе осморегуляции: Гипоосмотическая регуляция (на примере морских костистых рыб): В море они теряют воду через жаберную поверхность (обезвоживаются). Потеря воды происходит по осмотическому градиенту, т.к морская вода имеет большее осмотическое давление, чем кровь. Для возмещения дефицита воды рыбы пьют морскую воду, она всасывается вместе с солями Na+ и К+ в кровь. В жабрах имеются хлоридные клетки, которые удаляют из крови в морскую воду соли Na+ без воды – опреснение жидкостей внутренней среды. Часть солей Са2+ и Mg2+ удаляется почками и кишечником. Гиперосмотическая регуляция: У пресноводных животных часть солей сорбируется из внешней среды специальными клетками в жабрах или покровах тела, часть поступает с пищей, а избыток воды удаляется почкой.
|
7 Типы почек у позвоночных. Взаимо-связь выделит-х и половых протоков. У всех позвоночных органами выделения слу-жат парные почки мезодермального проис-хождения. У позвоночных в процессе эво-люции и эмбриогенеза закладывается 3 по-коления почек: 1 – пронефрос (головная по-чка) – орган выделения у зародышей анам-ний. У амниот (птицы, пресмыкающиеся, млекопитающие) закладываются, но не функционируют. Состоит из 6-12 нефронов. Нефрон пронефроса состоит из мерцатель-ной воронки (нефростом), которая откры-вается в целом и короткого прямого выде-лительного канальца, впадающего в вывод-ной проток пронефроса. Выделительные ка-нальцы обычно имеют единый фильтриру-ющий аппарат (сосудистый клубочек), рас-положенный вблизи нефростома. Несовер-шенство нефрона пронефроса – в отсутст-вии прямой связи между кровеносной и выделительной системами + в постоянном присутствии в целомической жидкости про-дуктов выделения. Предпочка подвергается редукции. У зародыша человека она созра-няется 40 часов. Основная ее роль в эмбри-огенезе – образование Вольфова протока из мочеточника, который является индуктором развития первичной почки. 2 – мезонефрос (первичная почка) – ее нефрон состоит из: мерцательной воронки (нефростом) откры-вающейся в целом; почечного тельца, кот состоит из 2хстенной капсулы Шум-Боу и мальпигиевых каплляров; извитого выдели-тельного канальца. Продукты выделения из капиллярного клубочка отфильтровываются в полость капсулы > по извитому канальцу > в мочеточник > мочевой пузырь > выво-дятся через клоаку или мочевое отверстие. Прогрессивные изменения нефрона первич-ной почки: появляется прямая связь между кровеносной и выделительной системами; кол-во продуктов выделения в целоме; удлинение и изменение формы канальца – начинается реабсорбция нужных в-в и происходит концентрация мочи; / кол-во нефронов почки (30-34 шт). У круглоротых, рыб и земноводных мезонефрос функцио-нирует в течение всей жизни и играет важ-ную роль в удалении излишков воды. У амниот – только на ранних стадиях зароды-шевого развития, дальше сменяется мета-нефрососм. У самцов амниот бОльшая часть чезонефроса развивается в придаток семенника и вместе с фольфовым каналом образует семявыносящий канал. У самок мезонефрос редуцируется. 3 – метанефрос – (вторичная туловищеая почка) – парный ор-ган выделения у амниот. Каждая поча соде-ржит более 1 млн нефронов. Нефрон состо-ит из: почечного канльца и выделительного канальца, который дифференцируется на проксимальный и дистальный канальцы и петлю Генле. Фильтрует продукты обмена только из крови. (см образование первич-ной и вторичной мочи). Эволюция проте-кает в 2х напрвленияз: 1 — / выделительной поверхности, 2 – создание механизма обрат-ного всасывания. |
8 Особенности выделения и осморегуля-ции в разных гр позвоночных. 1 – Ланцет-ник – над глоткой лежат около 100 пар неф-ридиев, которые открываются отверстием в атриальную полость и вдаются в полость целома (в виде 2х целомических мешков по бокам от хорды). Нефридии имеют особые клетки – соленоциты, в ножках которых имеется узкий канал с мерцательным волос-ком. Продукты распада фильтруются в целомическую полость > в соленоциты > в просвет нефридиальной трубки > через нефростомы наружу. 2 – Круглоротые: у эмрионов – парные предпочки, которые редуцируются, сменяясь туловищными. Почки круглоротых представляют первую фазу возникновения мальпигиевых телец. У низ нет объединенного фильтрующего клу-бочка и принимающей фильтрат капсулы. 3 – Хрящевые рыбы – мезонефрос. Основную массу почки занимают нефроны, состоящие из мальпигиевых телец. У некоторых сохра-няются и нефростомы. Сохраняя фильтра-ционный аппарат почек и биохимически обеспечивая почти полную изотоничность крови и тканевой жидкости внешней среде. Это было достигнуто удержанием моче-вины и солей в крови – непроницаемость для них жаберных лепестков и наличие особого сегмента почечного канальца для раебсорбции мочевины. 4 – Костные рыбы – органы выделния рыб не только выводят продукты распада, но и обеспечивают пос-тоянство осмотического давления. У них парные туловищные почки, которые у мно-гих сливаются вместе. Вольфовы каналы выполнят ф-ю мочеточников. У пресновод-ных рыб хорошо развиты капсулы Шум-Боу с клубочками; у морских рыб клубочки а размере или редуцированы; а – пресновод-ные рыбы – ч/з жабры в о-м поступает вода и || ч/з жабры выводятся соли, аммиак, т.к окружающая среда гипотонична. Почки вы-водят большое кол-во гипотоничной мочи; б – морские рыбы – см билет №2. 5 – Зем-новодные – мезонефрос. Покровы очень тонкие, и т.к среда гипотоническая, то вода ч/з покровы всасывается в о-м – встает про-блема выведения излишков овды, поэтому почки выделяют гипотоническую мочу. Со-ли поступают с пищей и всасываются всей поверхностью тела. Под кожей – запасы во-ды. большую роль в водном обмене играет приспособительное поведение – в тени и влаге. 6 – Пресмыкающиеся – тазовые почки, которые позволяют экономить воду. Мочи выделяется мало, а с пищей поступа-ет большое кол-во солей > необходимы об-разования, кот удаляли бы излишки солей. у пресмыкающихся продукты азотистого об-мена выводятся в виде мочевой кислоты – экономия воды по сравнению с экскрекцией мочевины – в 200 раз. 7 – Птицы – интенси-вное дыхание – нужно экономить воду – почки крупные (1-2% массы тела), мочевого пузыря нет. Размеры клубочков > интенсивность фильтрации и потеря воды небольшие. В нефроне появляется петля Генле. Выделяется мочевая кислота. |
Источник: StudFiles.net
Источник: chem21.info