Низшие позвоночные
ОСНОВЫ ПАЛЕОНТОЛОГИИ
Позвоночные
Учебно-методическое пособие
Красноярск
СФУ
УДК 566
О753
Составитель: О.В.Сосновская
О753 Основы палеонтологии. Позвоночные: учебно-методическое пособие [Текст] / сост. О.В.Сосновская. – Красноярск: Сиб. федер. ун-т, 2012. — 56 с.
Приведены краткие сведения по строению, систематике, образу жизни и стратиграфическому значению животных подтипа Позвоночные.
Для студентов, обучающихся по специальности 130101.65 «Прикладная геология».
УДК 566
Ó Сибирский
федеральный
университет, 2012
ВВЕДЕНИЕ
Позвоночные животные, ископаемые и современные, объединяются в подтип Vertebrata (Позвоночные или Черепные). Вместе с подтипами Tunicata (Оболочники), Conodontochordata (Конодонты) и Acraniata (Бесчерепные) они входят в состав типа Chordata (Хордовые). К этому типу относятся вторичноротые двусторонне (билатерально) симметричные животные с вторичной полостью тела и посегментным расположением главнейших систем органов (спинномозговых нервов, мускулатуры, осевого скелета и пр.).
Характерные особенности хордовых:
1. Наличие внутреннего осевого скелета – хорды, составляющей опору тела. У одних животных хорда сохраняется в течение всей жизни, у других – присутствует только на ранней стадии развития и замещается во взрослом состоянии позвонками.
2. Центральная нервная система имеет вид трубки и расположена на спинной стороне над хордой.
3. Наличие жаберных отверстий (от округлых до щелевидных). У наземных позвоночных они только закладываются у зародышей.
4. Дыхательная система — производной пищеварительной системы.
Хордовые имеют пищеварительный канал, который расположен под осевым скелетом. Канал начинается ротовым отверстием и заканчивается анальным. В брюшной полости расположены органы пищеварения, выделения и размножения. Газообмен и пищеварение непосредственно связаны друг с другом (легкие развиваются как полость, которая отшнуровывается от пищевода). Центральный орган кровообращения — сердце.
Хордовые животные обладают хорошей приспособляемостью: они обитают на земле, в воде и освоили воздушное пространство.
Наиболее вероятными предками хордовых считают червей и личинки древних иглокожих. Самые древние хордовые обнаружены в кембрии. К ним относятся юннаньзоон (Yunnanozoon) и хайкоуэлла (Haikouella) из нижнего кембрия Китая, пикайя (Pikaia) (рис.1) из среднего кембрия Канады и др.

Рис. 1. Пикайя (Pikaia) – хордовое животное из среднего кембрия [46]
Позвоночные – самая высокоорганизованная и сложно построенная группа среди хордовых животных. В ее истории большое значение имело прогрессивное развитие головного мозга и органов чувств, благодаря чему эти животные заняли господствующее положение на Земле.
Среди позвоночных нет видов, ведущих сидячий (прикрепленный) образ жизни. Они перемещаются в широких пределах, активно разыскивая и захватывая пищу, находя для размножения особей другого пола, спасаясь от преследования врагов. В связи с активным образом, у беспозвоночных хорошо развиты органы передвижения. Органы дыхания – жабры или легкие, у значительной части позвоночных существенное значение в дыхании имеет кожа. Жаберный аппарат представляет собой систему парных, обычно симметрично расположенных, щелей, служащих для сообщения глотки с наружной средой. Легкие в схеме представляют собой пару мешков, открывающихся в глотку через гортанную щель. Орган слуха объединен с органом равновесия. Кровеносная система замкнутая. Они относятся к раздельнополым животным.
Тело позвоночных покрыто кожей, состоящей из эпидермиса и собственно кожи. Из эпидермиса развиваются роговая чешуя пресмыкающихся, перья птиц, волосы, ногти, копыта и роговые чехлы млекопитающих. Собственно кожа дает начало чешуе, кожным зубам, кожным костям.
Скелет позвоночных может быть костным или хрящевым, внутренним и наружным. Внутренний скелет состоит из осевого скелета (хорда, позвоночник), который тянется от хвоста и заходит в голову, черепа и конечностей. Внешний скелет состоит из кожных или накладных костей на голове и плечевом поясе или имеет вид чешуйчатого покрова на остальном теле.
Скелет позвоночных достаточно полно отражает строение мягкого тела и всего организма в целом, поэтому по его остаткам можно изучить и представить не только внешний вид всего ископаемого животного, но и функции отдельных частей тела. Кроме того, развитие подтипа протекало непрерывно от начала палеозоя до наших дней, благодаря чему родственные связи между отдельными классами беспозвоночных устанавливаются более точно, чем в других группах животных. Все это явилось причиной того, что ископаемые представители подтипа использовались для изучения путей и закономерностей эволюционного развития всех животных.
Среди позвоночных выделяют две группы (раздела): анамнии (низшие позвоночные) – без зародышевых оболочек, и амниоты (высшие позвоночные) – с зародышевыми оболочками. В первую группу объединены инфратип Бесчелюстные, надкласс Рыбы (4 класса) и один класс Амфибии надкласса Четвероногие. В амниоты включены остальные классы.
По наличию или отсутствию челюстей (а также другим признакам) позвоночные подразделяют на два инфратипа: Бесчелюстные – Agnata (?-Q) и Челюстноротые – Gnathostomata (S-Q). У бесчелюстных рот работает как сосуще-всасывающий аппарат, у вторых как капкан.
Классификация позвоночных приведена ниже.
Тип Chordata Хордовые.
Подтип Vertebrata. Позвоночные (черепные).
Раздел Anamnia. Низшие позвоночные.
Инфратип Agnatha. Бесчелюстные.
Инфратип Gnatostomata. Челюстноротые.
Надкласс Pisces. Рыбы.
Класс Placodermi. Пластинокожие.
Класс Acanthodei. Акантоды
Класс Chondrichthyes. Хрящевые.
Класс Osteichthyes. Костные.
Надкласс Tetrapoda. Четвероногие.
Класс Amphibia. Земноводные (амфибии).
Раздел Amniota. Высшие позвоночные.
Класс Reptilia. Пресмыкающиеся (рептилии).
Класс Aves. Птицы.
Класс Mammalia. Млекопитающие (звери).
Предками позвоночных считаются низшие хордовые (оболочники, бесчерепные). Древнейшие позвоночные известны из отложений кембрия. Одним из них считается, например, хайкуихтис (рис. 2). В юре существовали все известные ныне классы позвоночных. Общее число современных видов около 40 тыс.
Рис. 2. Реконструкция представителя хордовых хайкуихтиса (Haikouichthys, ?2) [25].
Первоначально предки позвоночных жили в море, впоследствии некоторые из них заходили в пресные воды. Собственно позвоночные возникли в пресных водах и прошли здесь первые этапы эволюции. Благодаря развитой двигательной системе, водные позвоночные, главным образом, рыбы, широко расселились по водоёмам и проникли в море. Обитание в водоёмах, особенно с недостатком кислорода, подготовило их выход на сушу в девоне.
По сравнению с беспозвоночными, позвоночные имеют меньшее стратиграфическое значение. Однако с успехом используются остатки рыб для разработки стратиграфии девона, амфибий – континентальных отложений перми и триаса, млекопитающих – кайнозойских отложений и т.д. Ископаемые позвоночные широко применяются для палеогеографических реконструкций.
Надкласс Pisces. Рыбы
В надкласс входят животные, постоянно живущие в воде, дышащие жабрами и передвигающиеся с помощью плавников. Грудные и брюшные плавники являются парными, хвостовые, спинные и анальные – непарными. Последние поддерживаются специальным скелетом.
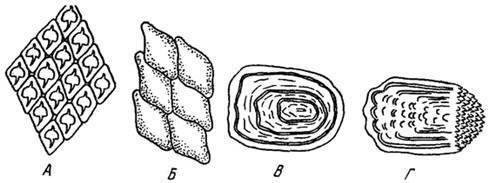
Осевая часть скелета рыб представлена позвоночником с двояковогнутыми позвонками. Кожа покрыта чешуей. Различают плакоидную, ганоидную, космоидную и костную чешуи (рис. 5).
Рис. 5. Типы чешуй рыб: А — плакоидная; Б — ганоидная; В — циклоидная;
Г — ктеноидная [48]
Плакоидная чешуя (рис. 5а) характерна для хрящевых рыб. Она создается кожными зубами, рассеянными по поверхности кожи. Чешуя имеет вид толстых пластин, часто округлого или овального очертания, расположенных в теле животного, и у многих рыб заканчивающимися шипами, выходящими наружу. В процессе жизни животного чешуя периодически сбрасывается.
Плакоидная чешуя дала начало остальным типам чешуй, а также зубам позвоночных.
Ганоидная чешуя (рис. 5б) – большие окостеневшие щитки обычно ромбической или круглой формы со скульптированной наружной поверхностью, образовавшиеся в результате срастания первичных плакоидных чешуй. На теле животного она располагается кольцами, образуя панцирь (верхний край каждой чешуи имеет шип, входящий в выемку чешуи, лежащей выше). Чешуя покрыта снаружи твердым блестящим слоем эмалеподобного вещества – ганоина. Помимо защитной функции, эта чешуя даёт опору мускулатуре и обеспечивает необходимую для движения упругость тела. Встречается у низших лучеперых рыб. В течение жизни организма не сменяется.
Примитивный вид ганоидной чешуи – космоидная чешуя (рис. 5в), характерная для древних кистеперых и двоякодышаших рыб. Ее наружная поверхность образована слоем космина (отсюда название) — видоизмененного дентина, но более плотного. Сверху чешуя покрыта твёрдым эмалеподобным дентином (эмаль), придающим ей характерный блеск. Космоидная чешуя имеет ромбическую форму и поверхность без скульптуры.
Костная чешуя (рис. 5г) представляет собой полупрозрачные тонкие костные пластинки разнообразной формы, но обычно округлых или почти четырехугольных очертаний. Она образовалась из ганоидной чешуи, у которой исчез поверхностный слой ганоина. Костные чешуи, как правило, черепицеобразно накладываются друг на друга. У большинства рыб чешуи — циклоидные, часто — с гладким наружным краем. У некоторых рыб, например у окунеобразных, чешуя — ктеноидная, имеющая зубчики по наружному краю. Чешуи растут в течение всей жизни рыбы.
Рыбы относятся к животным, имеющим непостоянную температуру тела. Они активны, ведут различный образ жизни. Размножаются икринками или являются живородящими.
Первые рыбы появились в силуре, а в девоне они уже — господствующая группа. В девоне кистеперые рыбы дали начало земноводным.
В надклассе рыб выделены 4 класса: Пластинокожие – Placodermi (S-D), Акантоды – Acanthodei (S2-P), Хрящевые рыбы – Chondrichthyes (D-ныне), Костные рыбы – Osteichthyes (D-ныне).
1.2.1.1 Класс Placodermi. Пластинокожие
Эти рыбы известны также под названием «панцирные рыбы» (рис. 6,7). Они были первыми позвоночными, у которых развились челюсти. Считается, что эти челюсти развились из жаберных дуг бесчелюстных позвоночных. Панцирные рыбы имели наружный скелет — панцирь, который покрывал голову и переднюю часть туловища.
Рис. 6. Панцирная рыба ботриолепис (Bothriolepis, D3) [1]
Панцирь слагался крупными костными пластинками, поверхность которых была покрыта бугорками и валиками. Настоящих зубов у панцирных рыб не было, а для разрушения добычи использовались или скошенные края пластин, образовывавших панцирь головы, или отдельные пластины. Парные плавники были хорошо развиты и у некоторых из них тоже защищены костными щитками. Внутренний скелет хрящевой, частично обызвествленный. Размеры тел различны: от небольших до 6, иногда 8-10 м в длину (дунклеостей).
Панцирные жили в пресной, преимущественно, и морской воде. Это обитатели мелководья. Образ жизни – придонные хищники.
Рыбы этого класса появились в силуре и полностью вымерли к концу девона. Благодаря массивности скелета, их представители сохранялись в ископаемом состоянии хорошо. Встречаются целые черепа, плечевые пояса и различные фрагменты скелета рыб. Есть находки цельных скелетов.
Рис. 7 Панцирная рыба дунклеостей (Dunkleosteus, D) [7]
Класс имеет важное стратиграфическое значение. Во-первых, пластинокожие быстро эволюционировали, что позволяет использовать их для дробного расчленения девонских отложений и достаточно точного определения относительного возраста, во-вторых, их существование и в морских, и в континентальных условиях дает возможность сопоставить разнофациальные отложения.
Класс не оставил потомков.
Характерные представители класса: ботриолепис — Bothriolepis (D3), дунклеостей — Dunkleosteus (D).
1.2.1.2 Класс Acanthodei. Акантоды
Акантоды («колючезубые рыбы») (рис. 8,9) относятся к древнейшей группе, сочетающей в себе черты панцирных, хрящевых и костных рыб. Они имели небольшие (длина 18-26, реже до 50 см) веретенообразные тела, покрытые плотно расположенными косыми рядами мелких четырехугольных чешуй ганоидного типа. Отдельные представители достигали 2 метров. Развитые челюсти акантод несли зубы, не имевшие эмалевого слоя (в отличие от зубов как хрящевых, так и костистых рыб). Акантоды были неплохими пловцами, питались придонными или медленно плавающими мелкими животными.
Рис. 8. Акантоды: а–б – эвтакантус (Euthacanthus, D)
(а – вид сбоку, б – вид с брюшной стороны), в – чешуя [1]
Внутренний скелет хрящевой, иногда с частичным окостенением. Все плавники, кроме хвостового, обладали сильным шипом.
Акантоды – в значительной степени представители пресноводных водоемов. Лишь в карбоне небольшое количество этих рыб встречено в морских отложениях.
Первые представители акантод появились в позднем силуре, в девоне они широко распространены, а в перми вымирают.
В ископаемом состоянии в речных и морских отложениях встречаются обычно разрозненные чешуйки и шипы акантод, а в озерных и лагунных отложениях могут быть обнаружены и цельные экземпляры.
Рис. 9. Ископаемая рыба акантода [23]
Акантоды используются для расчленения и определения возраста девонских отложений. Кроме того, они представляют большой интерес для познания эволюции рыб, т.к. являются одной из древнейших групп челюстноротых, сочетающих в себе черты основных классов рыб, и родоначальной для лучеперых костных рыб.
1.2.1.3 Класс Chondrichthyes. Хрящевые рыбы
К классу относятся рыбы с внутренним хрящевым скелетом (рис. 10). Современные представители — акулы, скаты и химеры.
Наружный скелет состоит из плакоидных чешуй (кожных зубов), иногда превращенных в головные или плавниковые колючки. Кожа может быть голой. Наружного костного панциря нет. Зубы у представителей класса разнообразны. Так, у пермской рыбы Helicoprion зубы были свернуты в плоскую спираль, состоящую из 2-3 оборотов. Скаты обладали тупыми, дробящими зубами, у акул они были острые, режущие.
| |
|
| |
Рис. 10. Хрящевые рыбы: а – скелет акулы [1], б — зубная спираль акулы геликоприон (Helicoprion, P1) [1], в-г – зубы разных хрящевых рыб
У хрящевых рыб, в отличие от костных, нет плавательного пузыря. В связи с этим, чтобы не опуститься на дно, они должны находиться в движении. Хрящевые рыбы – хищники. Среди древних их представителей преобладают пресноводные формы, среди современных – морские пелагические. Несколько видов встречаются в тропических реках.
Хрящевые рыбы появились в девоне, настоящие акулы – в карбоне, скаты – в юре.
В ископаемом состоянии от этих рыб остались главным образом только плакоидная чешуя, зубы и иглы плавников. Находки полных скелетов редки. Вместе с рыбами встречаются обильные копролиты различного облика и замещенные различными минералами. Зубы акул могут захороняться в массовом количестве. Горняки называли их «птичьими языками». Они широко используются в стратиграфии для дробного расчленения фанерозойских (с девона по четвертичную систему) отложений. Кроме того, акуловые имеют и биологическое значение. Они развивались быстро и, разделяясь на многочисленные группы, дали богатый материал для исследований общебиологического значения.
Характерная форма: геликоприон — Helicoprion (P1).
1.2.1.4 Класс Osteichthyes. Костные рыбы
Костные рыбы имеют окостеневший внутренний скелет и окостенения в наружном скелете. У некоторых рыб скелет может быть хрящевым (осетровые) или более или менее окостеневшим. Окостенение происходит за счет прижизненной минерализации межклеточного вещества карбонатами и сульфатами кальция и кристалликами апатита. На голове костных рыб присутствуют кожные кости, выполняющие покровную и защитную функцию. Тело покрыто космоидной, ганоидной или костной чешуей. Кожа может быть голой. Есть плавательный пузырь или легкое. Плавники поддерживаются костными лучами.
Форма тела костных рыб разнообразная: уплощенная (скаты), змееобразная, веретенообразная и торпедообразная. Длина тела колеблется от 0,7 см до 9 м.
Класс объединяет ископаемых и современных рыб, средой обитания которых являются и пресные и морские (всех глубин) бассейны. В современной ихтиофауне они составляют 95%. Кроме плавающих их представителей встречаются лазающие с помощью плавников рыбы (современные лучеперые в мангровых зарослях), ползающие рыбы (двоякодышащие и кистеперые) мелководья и пересыхающих водоемов, а также летучие рыбы тропиков, которые могут преодолевать расстояния над водой до 400 м.
Первые представители костных рыб, как предполагают, появились в позднем силуре, т.к. в девоне они уже представлены двумя разными группами. К концу палеозоя эти животные стали господствующей группой в пресноводных бассейнах и заселили моря.
В ископаемом состоянии костные рыбы встречаются обычно в виде скелетов или отпечатков, рыбы с незначительно развитым окостенением – в виде отдельных костей и зубов. Нередко скелеты рыб находят в известковых конкрециях. В мире имеется много местонахождений костных рыб, часто хорошей сохранности. Примером могут быть древнейшие костистые (одна из групп костных) рыбы в медистых сланцах цехштейна (Р2) Германии. С давних времен сохранилось их местное название «селедки медистых сланцев».
Костные рыбы разделяются на три подкласса: кистеперые – Crossopterygii (D-Q), двоякодышащие — Dipnoi (D2-Q) и лучеперые – (D-Q).
Кистеперые рыбы. Crossopterygii (рис. 11) имели веретеновидное тело, покрытое у древних видов космоидными чешуями – щитками, у более поздних видов – костными чешуями. Они были хищниками и имели хорошо развитые и многочисленные зубы. Зубы располагались вдоль краев челюстей и на небе. У рипидистий (предков амфибий) они характеризовались наличием складчатого дентина (лабиринтные зубы).
Рис. 11. Кистеперые рыбы девонского периода: а – пандерихтис (Panderichthys) (представитель группы кистеперых рыб рипидистий, предковых для тетрапод) [38], б – диптерус (Dipterus), в – голоптихиус (Holoptychius) [1]
Наиболее характерная особенность кистепёрых – парные плавники с мускулистой лопастью, включающей скелетную ось из нескольких кистеобразно разветвлённых сегментов (отсюда название), служащих для опоры о дно и имеющих отдаленное сходство с пятипалой конечностью четвероногих (рис. 12).
Интересно, что способность передвигать по грунту с помощью плавников имееся у некоторых более прогрессивных рыб и по настоящее время (илистый прыгун, бентозавр, рыбы-лягушки и др.).
Функцию дыхания у кистеперых рыб, кроме жабр, выполнял плавательный пузырь. Он периодически наполнялся атмосферным воздухом, а его стенки были пронизаны многочисленными кровеносными сосудами. Прямо из пузыря кислород попадал в кровь. Для дыхания атмосферным воздухом у ряда кистеперых рыб имелись внутренние носовые отверстия (хоаны).
Рис. 12. Строение кисти земноводного (а) и кистеперой рыбы (б) [49]
У кистеперых рыб было по три глаза: два больших — по бокам головы, а третий поменьше — на темени. Он в небо смотрел. Собственно, это был не совсем глаз, а теменное отверстие, связанное со светочувствительным органом. Потом этот глаз атрофировался, омертвел и исчез. У людей сохранился еще в мозгу остаток «третьего глаза» — особая шишковидная железка.
Кистеперые рыбы обитали как в морских, так и пресноводных бассейнах. Одна группа этих рыб (целаканты), не имевшая легочного дыхания (без хоан), в палеозое были пресноводными, в мезозое перешли в море, где на значительных глубинах (400-1000 м) у Коморских островов (близ юго-восточного побережья Африки) сохранились до наших дней. Впервые представитель целакантов – Latimeria, был обнаружен в 1938 году и назван по имени г-жи Латимер, обнаружившей эту рыбу в улове рыболовецкого судна и сохранившую её для науки. Другая группа кистеперых, обладавшая внутренними ноздрями (хоаны), считают, явилась предковой для наземных позвоночных – земноводных. Эти рыбы первыми «вышли на сушу».
Одной из наиболее «продвинутых» известных в настоящее время кистепёрых рыб являлся тиктаалик (Tiktaalik) (рис. 13), имевший ряд переходных признаков, сближающих его с земноводными.
Кистеперые рыбы сохраняются, как правило, в виде чешуй и зубов и встречаются довольно часто, особенно в континентальных отложениях палеозоя. Они используются для их расчленения и корреляции.
Характерные представители: пандерихтис – Panderichthys (D), целаканты.
Рис. 13. Кистеперая рыба тиктаалик (Tiktaalik) из позднего девона – переходная форма между рыбами и земноводными [26]
Двоякодышащие рыбы (Dipnoi) (рис. 14) обладают также способностью дышать в воде и на суше. Для этого они, как и кистеперые, используют жабры и плавательный пузырь, который может играть роль легких. Двоякодышащие имеют много общих черт с кистеперыми, поэтому их нередко объединяют в один подкласс Лопастеперые – Sarcopterigii. Считается, что и те, и другие произошли от одного предка.
Рис. 14. Протоптерус (Protopterus) –
современная двоякодышащаярыба [14]
Отличие этих рыб состоит в некоторых особенностях строения тела. Например, у них отсутствует «третий глаз». Длина до 2 м.
Современные двоякодышащие являются обитателями пресноводных водоемов. При их пересыхании перебираются в другой водоем или впадают в спячку. Когда во время шестимесячной засухи озеро Чад в Африке уменьшает свою площадь почти на одну треть и обнажается илистое дно, местные жители отправляются на рыбалку, прихватывая с собой мотыги. Они отыскивают на обсохшем дне холмики и выкапывают из каждого глиняную капсулу с рыбой, сложившейся вдвое. В таком состоянии двоякодышащая рыба протоптер ожидает следующего периода дождей, который наступает через 6–9 месяцев.
Двоякодышащие рыбы распространены со среднего девона по ныне. Характерный представитель: протоптерус – Protopterus (современный).
Лучеперые рыбы. Actinopterygii в отличие от кистеперых и двоякодышащих рыб, как правило, обладают костным позвоночником, и только у немногих сохраняется хорда или ее остатки. У них нет мясистых плавников (лопастей), хоан и «третьего глаза». Эти рыбы имеют уплощенные плавники с лучевидными шипами хрящевого и костного состава. Длина тела – до 9 м.
Наиболее известные их древние представители – это пресноводные и морские палеониски (рис. 15, 16). Эти рыбы имели небольшие размеры – до 40 см и могли сохраняться целиком. Чешуя ганоидная, дифференцированная по положению и морфологии на теле. Жили со среднего девона по ранний мел.
Рис. 15. Ископаемая рыба палеониска из девона [11]
Рис. 16. Верхнедевонская палеонискообразная рыба Cheirolepis (реконструкция) [11]
Лучеперые рыбы в настоящее время составляют более 90 %. Первые их представители известны из среднего девона, однако их предки, имеющие ряд признаков лучеперых рыб, находят уже в силуре. Например, рыба Psarolepis. Ископаемые рыбы используются в стратиграфии.
ОСНОВЫ ПАЛЕОНТОЛОГИИ
Позвоночные
Составитель:
ОСНОВЫ ПАЛЕОНТОЛОГИИ
Позвоночные
Учебно-методическое пособие
Красноярск
СФУ
УДК 566
О753
Составитель: О.В.Сосновская
О753 Основы палеонтологии. Позвоночные: учебно-методическое пособие [Текст] / сост. О.В.Сосновская. – Красноярск: Сиб. федер. ун-т, 2012. — 56 с.
Приведены краткие сведения по строению, систематике, образу жизни и стратиграфическому значению животных подтипа Позвоночные.
Для студентов, обучающихся по специальности 130101.65 «Прикладная геология».
УДК 566
Ó Сибирский
федеральный
университет, 2012
ВВЕДЕНИЕ
Позвоночные животные, ископаемые и современные, объединяются в подтип Vertebrata (Позвоночные или Черепные). Вместе с подтипами Tunicata (Оболочники), Conodontochordata (Конодонты) и Acraniata (Бесчерепные) они входят в состав типа Chordata (Хордовые). К этому типу относятся вторичноротые двусторонне (билатерально) симметричные животные с вторичной полостью тела и посегментным расположением главнейших систем органов (спинномозговых нервов, мускулатуры, осевого скелета и пр.).
Характерные особенности хордовых:
1. Наличие внутреннего осевого скелета – хорды, составляющей опору тела. У одних животных хорда сохраняется в течение всей жизни, у других – присутствует только на ранней стадии развития и замещается во взрослом состоянии позвонками.
2. Центральная нервная система имеет вид трубки и расположена на спинной стороне над хордой.
3. Наличие жаберных отверстий (от округлых до щелевидных). У наземных позвоночных они только закладываются у зародышей.
4. Дыхательная система — производной пищеварительной системы.
Хордовые имеют пищеварительный канал, который расположен под осевым скелетом. Канал начинается ротовым отверстием и заканчивается анальным. В брюшной полости расположены органы пищеварения, выделения и размножения. Газообмен и пищеварение непосредственно связаны друг с другом (легкие развиваются как полость, которая отшнуровывается от пищевода). Центральный орган кровообращения — сердце.
Хордовые животные обладают хорошей приспособляемостью: они обитают на земле, в воде и освоили воздушное пространство.
Наиболее вероятными предками хордовых считают червей и личинки древних иглокожих. Самые древние хордовые обнаружены в кембрии. К ним относятся юннаньзоон (Yunnanozoon) и хайкоуэлла (Haikouella) из нижнего кембрия Китая, пикайя (Pikaia) (рис.1) из среднего кембрия Канады и др.
Рис. 1. Пикайя (Pikaia) – хордовое животное из среднего кембрия [46]
Позвоночные – самая высокоорганизованная и сложно построенная группа среди хордовых животных. В ее истории большое значение имело прогрессивное развитие головного мозга и органов чувств, благодаря чему эти животные заняли господствующее положение на Земле.
Среди позвоночных нет видов, ведущих сидячий (прикрепленный) образ жизни. Они перемещаются в широких пределах, активно разыскивая и захватывая пищу, находя для размножения особей другого пола, спасаясь от преследования врагов. В связи с активным образом, у беспозвоночных хорошо развиты органы передвижения. Органы дыхания – жабры или легкие, у значительной части позвоночных существенное значение в дыхании имеет кожа. Жаберный аппарат представляет собой систему парных, обычно симметрично расположенных, щелей, служащих для сообщения глотки с наружной средой. Легкие в схеме представляют собой пару мешков, открывающихся в глотку через гортанную щель. Орган слуха объединен с органом равновесия. Кровеносная система замкнутая. Они относятся к раздельнополым животным.
Тело позвоночных покрыто кожей, состоящей из эпидермиса и собственно кожи. Из эпидермиса развиваются роговая чешуя пресмыкающихся, перья птиц, волосы, ногти, копыта и роговые чехлы млекопитающих. Собственно кожа дает начало чешуе, кожным зубам, кожным костям.
Скелет позвоночных может быть костным или хрящевым, внутренним и наружным. Внутренний скелет состоит из осевого скелета (хорда, позвоночник), который тянется от хвоста и заходит в голову, черепа и конечностей. Внешний скелет состоит из кожных или накладных костей на голове и плечевом поясе или имеет вид чешуйчатого покрова на остальном теле.
Скелет позвоночных достаточно полно отражает строение мягкого тела и всего организма в целом, поэтому по его остаткам можно изучить и представить не только внешний вид всего ископаемого животного, но и функции отдельных частей тела. Кроме того, развитие подтипа протекало непрерывно от начала палеозоя до наших дней, благодаря чему родственные связи между отдельными классами беспозвоночных устанавливаются более точно, чем в других группах животных. Все это явилось причиной того, что ископаемые представители подтипа использовались для изучения путей и закономерностей эволюционного развития всех животных.
Среди позвоночных выделяют две группы (раздела): анамнии (низшие позвоночные) – без зародышевых оболочек, и амниоты (высшие позвоночные) – с зародышевыми оболочками. В первую группу объединены инфратип Бесчелюстные, надкласс Рыбы (4 класса) и один класс Амфибии надкласса Четвероногие. В амниоты включены остальные классы.
По наличию или отсутствию челюстей (а также другим признакам) позвоночные подразделяют на два инфратипа: Бесчелюстные – Agnata (?-Q) и Челюстноротые – Gnathostomata (S-Q). У бесчелюстных рот работает как сосуще-всасывающий аппарат, у вторых как капкан.
Классификация позвоночных приведена ниже.
Тип Chordata Хордовые.
Подтип Vertebrata. Позвоночные (черепные).
Раздел Anamnia. Низшие позвоночные.
Инфратип Agnatha. Бесчелюстные.
Инфратип Gnatostomata. Челюстноротые.
Надкласс Pisces. Рыбы.
Класс Placodermi. Пластинокожие.
Класс Acanthodei. Акантоды
Класс Chondrichthyes. Хрящевые.
Класс Osteichthyes. Костные.
Надкласс Tetrapoda. Четвероногие.
Класс Amphibia. Земноводные (амфибии).
Раздел Amniota. Высшие позвоночные.
Класс Reptilia. Пресмыкающиеся (рептилии).
Класс Aves. Птицы.
Класс Mammalia. Млекопитающие (звери).
Предками позвоночных считаются низшие хордовые (оболочники, бесчерепные). Древнейшие позвоночные известны из отложений кембрия. Одним из них считается, например, хайкуихтис (рис. 2). В юре существовали все известные ныне классы позвоночных. Общее число современных видов около 40 тыс.
Рис. 2. Реконструкция представителя хордовых хайкуихтиса (Haikouichthys, ?2) [25].
Первоначально предки позвоночных жили в море, впоследствии некоторые из них заходили в пресные воды. Собственно позвоночные возникли в пресных водах и прошли здесь первые этапы эволюции. Благодаря развитой двигательной системе, водные позвоночные, главным образом, рыбы, широко расселились по водоёмам и проникли в море. Обитание в водоёмах, особенно с недостатком кислорода, подготовило их выход на сушу в девоне.
По сравнению с беспозвоночными, позвоночные имеют меньшее стратиграфическое значение. Однако с успехом используются остатки рыб для разработки стратиграфии девона, амфибий – континентальных отложений перми и триаса, млекопитающих – кайнозойских отложений и т.д. Ископаемые позвоночные широко применяются для палеогеографических реконструкций.
РАЗДЕЛ ANAMNIA. НИЗШИЕ ПОЗВОНОЧНЫЕ
1. 1 ИНФРАТИП AGNATA. БЕСЧЕЛЮСТНЫЕ
Это рыбообразные позвоночные животные (рис. 3,4) без челюстей и парных плавников, имеющие хорду. В отличие от бесчерепных – более примитивной группы хордовых, у них имеется череп. Внутренний скелет хрящевой. Внешний скелет состоит из разнообразных кожных костных образований (зубы, чешуи, пластины, щиты и панцири). Кожа современных представителей покрыта только слизью.
Длина от нескольких см (веретенообразные животные) до 1-2 м (уплощенные донные формы). Рот круглый, иногда щелевидный. У многих видов образовались в замену настоящим челюстям зубы, свободно сидящие в ротовой полости.
Рис. 3. Современные бесчелюстные: а–б – минога (а – общий вид сбоку, б – ротовая воронка), в – миксина; древние бесчелюстные: г – цефаласпис(Cephalaspis, D1), д – дрепанаспис (Drepanaspis, D1), ж – гемицикласпис (Hemicyclaspis, D1), e – телодус (Thelodus, O-S) [1].
Бесчелюстные – это обитатели пресных и солоноватых вод прибрежной зоны морей, они были приспособлены к пассивному питанию (всасывали или подхватывали мелкие частицы пищи вместе с водой).
Бесчелюстные являются первой по времени появления группой позвоночных. Самые древние представители их известны в кембрийских отложениях (см. рис. 2). Считается, что они являются предками рыб. Современные представители инфратипа – миноги и миксины. В некоторых классификациях в состав бесчелюстных включают конодонтофораты.
Бесчелюстные используются для определения возраста отложений ордовика, силура и нижнего девона. Характерные древние представители инфратипа: телодус — Thelodus (O-S), цефаласпис — Cephalaspis (D1), гемицикласпис — Hemicyclaspis (D1).
Рис. 4. Бесчелюстное животное цефаласпис (Cephalaspis, D1)
1.2 ИНФРАТИП GNATOSTOMATA. ЧЕЛЮСТНОРОТЫЕ
В эту ветвь объединяют животных, имеющих челюсти, которые образованы из передней пары жаберных дуг. Они служат для захвата и измельчения пищи, защиты и нападения. Хорда сохраняется или заменена позвонками. Внутренний скелет хрящевой или костный. Парные конечности (в виде парных плавников, пятипалых конечностей или крыльев) присутствуют, но у некоторых представителей вторично утрачены. Органы дыхания – жабры или легкие (у наземных животных).
В водной среде челюстноротые представлены рыбами, на суше – тетраподами, или четвероногими (земноводные, пресмыкающиеся, птицы и млекопитающие). Встречаются с позднего силура.
В настоящее время челюстноротые — преобладающая на нашей планете группа позвоночных животных. Они составляют 99,8 % всего числа видов типа хордовых.
Надкласс Pisces. Рыбы
В надкласс входят животные, постоянно живущие в воде, дышащие жабрами и передвигающиеся с помощью плавников. Грудные и брюшные плавники являются парными, хвостовые, спинные и анальные – непарными. Последние поддерживаются специальным скелетом.
Осевая часть скелета рыб представлена позвоночником с двояковогнутыми позвонками. Кожа покрыта чешуей. Различают плакоидную, ганоидную, космоидную и костную чешуи (рис. 5).
Рис. 5. Типы чешуй рыб: А — плакоидная; Б — ганоидная; В — циклоидная;
Г — ктеноидная [48]
Плакоидная чешуя (рис. 5а) характерна для хрящевых рыб. Она создается кожными зубами, рассеянными по поверхности кожи. Чешуя имеет вид толстых пластин, часто округлого или овального очертания, расположенных в теле животного, и у многих рыб заканчивающимися шипами, выходящими наружу. В процессе жизни животного чешуя периодически сбрасывается.
Плакоидная чешуя дала начало остальным типам чешуй, а также зубам позвоночных.
Ганоидная че
Источник: cyberpedia.su
10. Морфофизиологические и экологические особенности амниот. Принципиальные отличия от анамний.
Анамнии (лат. Anamnia) или низшие позвоночные — парафилетическая группа, включающая позвоночных животных, не имеющих зародышевых оболочек. В отличие от амниот, у анамний в процессе эмбрионального развития не возникает зародышевой оболочки — амниона и особого зародышевого органа — аллантоиса. Анамнии связаны в своём существовании с водной средой, в которой они проводят либо всю жизнь, либо начальные стадии (яйцевые и личиночные). К анамниям относятся все позвоночные, за исключением амниот, то есть бесчелюстные и различные группы рыб, а также амфибии. Оплодотворение у большинства анамний наружное, у хрящевых рыб и части земноводных появляется внутреннее оплодотворение. Яйца окружены студенистой оболочкой, которая обеспечивает сохранение формы яйца в воде. Желтка мало, вода поступает из внешней среды через проницаемые яйцевые оболочки. Яйцеклетка в процессе развития проходит полное дробление. Личинка, сформировавшись, разрывает оболочки и ведёт водный образ жизни. Дыхание у личинок жаберное. Земноводные для формирования взрослой особи должны пройти через метаморфоз. В ходе его образуются конечности для наземного перемещения, перестраивается корневая система и происходит переход к лёгочному дыханию. У некоторых земноводных возникли эволюционные приспособления для развития яиц вне водной среды.
Амнио́ты, или высшие позвоночные (лат. Amniota) — монофилетическая группа (клада) позвоночных животных, характеризующихся наличием зародышевых оболочек. Известны с раннего карбона. Группа входит в состав надкласса Tetrapoda (четвероногие); включает в себя пресмыкающихся, птиц, а также млекопитающих и прочих синапсид (остальные представители которых вымерли к концу нижнего мела). Все амниоты имеют внутреннее оплодотворение (в отличие от анамний, у которых оплодотворение внешнее). Размножение их происходит на суше; только немногие виды (как китообразные) вернулись к размножению в воде. При эмбриональном развитии амниот развиваются две зародышевые оболочки — амнион и сероза, формирующие амниотическую полость, от которой и происходит название таксона (Э. Г. Геккель 1866 г.). Амниотическая полость заполнена жидкостью, и зародыш находится как бы в маленьком водоёме, условия среды которого относительно постоянны. Эволюционно зародышевые оболочки возникли для обеспечения возможности развития эмбриона в воздушной среде. Параллельно с амнионом из вентральной стенки задней кишки эмбриона возникает зародышевый мочевой пузырь — аллантоис для сбора жидких продуктов обмена. Стенки аллантоиса богаты кровеносными сосудами, через которые идёт газообмен. У зародышей амниот формируется только одна пара жаберных щелей, превращающаяся в полость среднего уха. Большинство амниот откладывают характерные яйца. При этом студенистая оболочка яйцеклеток рыб и амфибий заменяется у амниот пергаментообразной или пропитанной известью скорлупой, через которую свободно проходит воздух; такие яйца богаты желтком (т. е. полилецитальны). Исключение составляют некоторые чешуйчатые, вымершие ихтиозавры, а среди млекопитающих — сумчатые и плацентарные; для этих групп характерно живорождение, и их яйцеклетки желтка не содержат (т. е. алецитальны). Соответственно, развитие амниот до типично наземной формы происходит либо в яйце, либо в матке под защитой зародышевых оболочек, а фаза водной личинки (головастика) полностью отсутствует.
Различия отдельно.
Яйца анамний окружены студенистой оболочкой, обеспечивающей сохранение формы яйца в воде; желтка относительно немного, а необходимая для развития зародыша вода поступает извне через проницаемые яйцевые оболочки. В яйцах амниот заметно возрастает количество желтка, содержащего необходимые для формирования зародыша пластические и энергетические вещества. Резко увеличивается белковая оболочка (белок яйца), содержащая достаточный для развития зародыша запас воды.
Различия взрослых особей. Особенности строения взрослых амниот, отличающие их от анамний, связаны с приспособлениями к жизни на суше и в той или иной степени затрагивают все системы органов. Кожа анамний проницаема для воды и газов и покрыта слизью, выделяемой многочисленными кожными железами. Она участвует в газовом и водном обмене и удалении продуктов распада. Защитные кожные образования — чешуи, покровные кости — развиваются в соединительнотканном слое кожи. У амниот резко сокращается количество кожных желез (кроме млекопитающих), а поверхностные слои эпидермиса ороговевают (в клетках накапливается кератогиалин), что делает кожу мало проницаемой для воды и газов. Это исключает ее участие в дыхании и выделении, но вместе с тем предохраняет организм от иссушения. Защитные роговые образования кожи амниот — чешуи, когти, перья, волосы — производные эпидермиса. Роговые чешуи улучшают защиту тела от механических и химических повреждений, а у птиц и млекопитающих перьевой и волосяной покров выполняет и теплоизолирующую функцию, обеспечивая теплокровность.
Совершенствование опорно-мышечной системы значительно увеличивает подвижность амниот по сравнению с анамниями (земноводными). Это выражается в полном окостенении скелета, в большей дифференцировке позвоночного столба, усилении поясов конечностей и укреплении их связи с осевым скелетом, в большем развитии и дифференцировке мускулатуры.
Усиление челюстей, развитие жевательной мускулатуры и дальнейшая дифференцировка пищеварительного тракта позволили расширить спектр используемых кормов и повысить степень их усвоения. Возрастание потребления кислорода обеспечивается увеличением поверхности легких и интенсификацией дыхания благодаря образованию грудной клетки. У пресмыкающихся, по сравнению с земноводными, усиливается разобщенность большого и малого кругов кровообращения, а у птиц и млекопитающих они полностью разделены. Число эритроцитов в единице объема крови увеличивается, а их размеры уменьшаются; поэтому возрастает общая поверхность эритроцитов и увеличивается кислородная емкость крови. Параллельно растет масса красного костного мозга.
Характерные для анамний мезонефрические почки функционируют только у зародышей амниот. Во второй половине эмбрионального развития формируются метанефрические, или тазовые, почки, одновременно развиваются их протоки — мочеточники. У самок амниот редуцируются мезонефрические почки и вольфовы каналы; сохраняются выполняющие функцию яйцеводов мюллеровы каналы. У самцов сохраняется лишь часть мезонефрической почки, становящейся придатком семенника; вольфовы каналы выполняют функцию семяпроводов. В метанефрической почке, по сравнению с мезонефрической, почечные канальцы (нефроны) заметно усложняются. Выделение продуктов распада идет не только путем фильтрации плазмы из капилляров клубочков в просвет боуменовых капсул, но и секрецией железистыми клетками стенок почечных канальцев. Этот фильтрат — первичная моча, проходя по почечному канальцу, существенно изменяется, так как через его стенки идет обратное всасывание воды и абсорбция ряда нужных орканизму веществ — солей, органических молекул и т. п. Благодаря этому метанефрическая почка амниот не только служит практически единственным органом выделения, но активно участвует в водном и солевом обмене, обеспечивая экономию воды.
У амниот возрастают относительные размеры головного мозга, особенно, и мозжечка. У пресмыкающихся и птиц в крыше переднего мозга увеличиваются скопления нервных клеток, а у млекопитающих они разрастаются в кору больших полушарий — неопаллиум, где образуются новые высшие мозговые центры. Изменения в деталях строения рецепторов повышают их эффективность
Источник: StudFiles.net
Беспозвоночные
Поведение зародышей беспозвоночных изучено сравнительно слабо, главным образом, кольчецов, моллюсков и членистоногих. Но очевидно, что двигательные реакции начинаются уже на ранних стадиях развития.
Примеры: зародыши некоторых головоногих моллюсков уже на ранних стадиях развития вращаются внутри яйца вокруг своей оси со скоростью один оборот в час. Другие перемещаются от одного полюса яйца к другому. При этом движения зародышей осуществляются с помощью ресничек (вспомним типичную для моллюсков ресничную личинку трохофору, стадия которой и происходит под оболочкой яйца).
У рачков-мизид (из подкласса Высших раков) первоначально наблюдаются эндогенные спонтанные беспорядочные вздрагивания и подергивания. Аналогичные, но ритмичные движения головы и других частей наблюдаются у эмбрионов морских козочек (рачки из отряда Бокоплавы).
К концу эмбриогенеза у беспозвоночных уже оказываются сформированными базовые инстинктивные реакции, необходимые для выживания.
Примеры: у тех же рачков-мизид к моменту вылупления развита реакция избегания. У эмбрионов дафний (подкласс Листоногие раки) антенны, служащие у взрослых особей для плавания, начинают двигаться на средних этапах эмбриогенеза, а к моменту рождения они поднимаются и принимают положение, необходимое для выполнения плавательных движений. За несколько часов до выхода из оболочек яйца антенны двигаются наиболее интенсивно.
На последних этапах эмбриогенеза формируются рефлекторные реакции, обусловленные стимулами из внешней среды.
Так, у мизид первоначально эндогенные спонтанные движения к концу эмбриогенеза начинают возникать в ответ на прикосновение к икринке. У морских козочек рефлекторный ответ на тактильное раздражение возникает в день вылупления. Таким образом, первоначальные спонтанные движения зародышей беспозвоночных постепенно заменяются рефлекторными. Это позволяет говорить об «эмбриональном научении», при котором эндогенно обусловленные движения связываются с внешними раздражителями, что сопряжено с глубокими морфофункциональными перестройками организма.
Низшие позвоночные (анамнии)
У рыб и амфибий первичная двигательная активность генерализованна и эндогенна, т.е. происходят беспорядочные (вначале) движения всего тела под влиянием не внешних раздражителей, а внутренних импульсов. При этом из первоначально генерализованных движений всего тела возникают все более дифференцированные движения отдельных органов по мере их развития. Они появляются в строгих последовательности и ритме в зависимости от созревания соответствующих нервных связей.
Примеры: у зародышей хвостатых амфибий (работы Когхилла) первоначально развиваются
– общие изгибания туловища, сначала С-образные, позже S-образные;
– затем к ним добавляются общие движения конечностей (слитые с движениями туловища);
– затем одиночные движения конечностей независимо от туловища;
– позже согласованные с туловищем и друг с другом движения всех четырех конечностей;
– наконец, формируются совершенные плавательные движения.
Сходным образом развиваются плавательные движения и у бесхвостых амфибий. Интересно, что если выход из яйцевых оболочек задерживается (у жабы Eleuterodactylus martinicensis с о-ва Ямайка стадия личинки проходит в яйце), то личинка (головастик) совершает все те движения, которые присущи и ее свободноживущим родственникам, но внутри икринки.
Эти и другие исследования эмбрионального поведения амфибий показали, что для них справедлив принцип Когхилла: в процессе эмбрионального развития происходит вычленение локальных движений из целостных, генерализованных. Кроме того, очевидно, что некоторые важные двигательные навыки, прежде всего, плавательные движения и оптомоторные реакции (реакции слежения за движущимся объектом, ориентировочные) формируются в результате созревания соответствующих внутренних структур, особенно нервно-мышечного аппарата. Этот процесс получил название эмбрионального созревания.Поведение развивается в результате влияния эндогенных факторов и при экспериментальных нарушениях не может быть скорректировано.
Доказательства:
А. Опыты Кармайкла с хвостатыми амфибиями.
· При выращивании зародышей амбистомы в анестезирующем растворе ацетонхлороформа происходило обездвиживание, но морфогенез и рост протекали без нарушений. После завершения развития животное плавало нормально (не имея возможности упражняться в этом виде локомоций).
· У эмбрионов саламандр пересаживались зачатки конечностей таким образом, что стопы оказывались повернутыми в обратную сторону. Если бы развитие шагания определялось эмбриональным упражнением (афферентная обратная связь), то в ходе эмбриогенеза произошла бы функциональная коррекция и восстановление нормального поступательного движения. Этого не случилось, и после вылупления оперированные животные пятились от положительных раздражителей вместо того, чтобы приближаться к ним.
Б. Опыты Сперри с бесхвостыми амфибиями (лягушками). При перевертывании зачатков глазных яблок на 1800 у эмбрионов, выросшие лягушки демонстрировали инверсию оптокинетической реакции: при движении цели вверх лягушка поворачивала голову вниз и наоборот. Коррекции также не происходило.
Таким образом, у анамний формирование локомоторных и оптомоторных движений (а также некоторых других) происходит в результате эндогенно обусловленного созревания внутренних функциональных структур.
Под влиянием результатов исследования эмбрионального поведения амфибий американский ученый Л. Кармайкл счел процессы эмбрионального созревания единственным фактором развития поведения. Более того, он распространил этот механизм и на постнатальный период, считая, что все поведенческие реакции, которые проявляются в это время, также развиваются по типу созревания. Кармайкл допускал, что даже у человека поведение на «девять десятых» является врожденным.
Амниоты
Птицы
Эмбриональное поведение птиц изучалось очень подробно на зародышах домашней курицы. Период инкубации длится 21 день. Движения зародыша начинаются уже на 3,5 – 4-е сутки с переднего конца к заднему. Они генерализованны. Самостоятельные движения органов развиваются позднее.
Гамбургер разделил двигательные реакции куриного зародыша на три категории:
А. Спонтанная подвижность – результат автономных самоуправляемых нервных разрядов, генерируемых в ЦНС (без изменения внутренней среды).
В. Эндогенно стимулируемая подвижность, вызванная гуморальными агентами, уровнем накопления продуктов обмена веществ, снабжения тканей кислородом или иными изменениями внутренней среды. Изучена недостаточно.
С. Вызванная подвижность – реакция на внешние раздражители.
Спонтанные движения начинаются самыми первыми с 3,5-го дня в виде небольших нерегулярных наклонов головы. Достигают пика на 13-й день. Движутся голова, туловище, конечности, пальцы, глаза, клюв. Движения бесцельны, отрывисты, судорожны, каждый орган «работает» в своем ритме. Эти отдельные ритмы, генерируются в разных участках спинного мозга. К 17-му дню движения становятся высокоинтегрированными, создают удобную для вылупления позу. Это связано с развитием головного мозга, который начинает контролировать отдельные ритмы. В последние дни перед вылуплением зародыш издает звуки.
Следует отметить, что у позвоночных иннервация соматической мускулатуры туловища и конечностей предшествует замыканию рефлекторных дуг. Таким образом, спонтанные движения носят нерефлекторный характер. Эти движения являются ритмичными, так они обусловлены спонтанными нейрогенными ритмами. Эндогенная ритмика в нервно-мышечных структурах сохраняется на протяжении всей жизни животного и является важным фактором созревания элементов врожденного поведения.
Движения, вызванные внешними стимулами, начинаются на 7-й день. К этому моменту происходит замыкание рефлекторных дуг. Сначала включается тактильная чувствительность: зародыш начинает реагировать на прикосновение кисточкой (сначала около клюва). Проприорецептивные стимулы вызывают движения на 10-й день. Интересно, что у цыпленка в яйце происходит самостимуляция (когда он прикасается одной частью тела, например, ногой или крылом к другой части, например, к голове). Некоторые исследователи полагали самостимуляцию ведущим механизмом развития поведения эмбриона (см. концепцию эмбриональной тренировки Куо). Однако, по последним данным тактильно-проприорецептивная стимуляция не влияет на частоту и периодичность движений, производимых эмбрионом на протяжении 2–2,5 недель инкубации. Хотя изолированное крыло куриного эмбриона способно отвечать на тактильные раздражения еще до замыкания рефлекторных дуг, у зародыша в нормальных условиях двигательная активность проявляется лишь в общих спонтанных телодвижениях.
Реакции на зрительные и звуковые раздражители появляются в последние дни развития. Реакции на громкие звуки обнаруживаются после 14-19-го дня, оптические реакции – с 17-18-х суток инкубации. Однако, как и в случае с тактильной чувствительностью, можно вызвать эти реакции на ранних этапах эмбриогенеза – с 5-го дня инкубации, т.е. еще до замыкания рефлекторных дуг, при воздействии мощных звуковых и световых сигналов. Не следует забывать, что это экстремальные, биологически неадекватные воздействия, которые возможно, выступают не как носители акустической или оптической информации, а как физические агенты, воздействующие непосредственно на соответствующие ткани.
Таким образом, потенциальные способности реагировать на внешние раздражители эмбрион получает довольно рано, но использовать их начинает лишь на поздних этапах эмбриогенеза. Однако если воздействовать на эмбрион биологически адекватными звуками на такой стадии, когда он еще не реагирует на подобные стимулы, в дальнейшем это положительно сказывается на слуховых реакциях организма (данные американского исследователя Г. Готлиба). Практическое применение этого важного заключения к человеческому эмбриону состоит в том, что будущие родители могут формировать акустические реакции и предпочтения своего ребенка задолго до того, как он появится на свет.
КОНЦЕПЦИЯ ЭМБРИОНАЛЬНОЙ ТРЕНИРОВКИ КУО
Известный американский исследователь Цин Янг Куо в 20-30-х годах прошлого века, активно изучая поведение куриных эмбрионов, разработал и осуществил виртуозные операции: вставлял плексигласовые окошки в яйцо, перемещал зародыш внутри яйца и т.п. Проведя наблюдения на сотнях экземпляров, он пришел к идее, что двигательные реакции зародышей формируются не в результате эмбрионального созревания соответствующих нервно-мышечных структур, а при накоплении зародышем двигательного опыта под влиянием особых условий, складывающихся в яйце.
Источник: megaobuchalka.ru