Преимущественно бесполым путем размножается
Размножение, или самовоспроизведение, — способность организмов воспроизводить себе подобных, одно из основных свойств живых существ. Различают половое и бесполое размножение.
Размножение у животных. При бесполом размножении одна родительская особь может делиться или почковаться, образуя две или больше дочерних особей, генетически одинаковых с материнской. Этот тип размножения встречается в основном у беспозвоночных животных, иногда бесполое размножение чередуется с половым (см. Чередование поколений).
Одноклеточные животные (амебы, парамеции) размножаются путем деления — митоза. Микроорганизмы образуют споры, из которых затем развивается новый организм.
Наиболее распространенный способ вегетативного размножения — почкование. Так размножаются, например, гидры. Организм, образовавшийся из почки, может отделиться и начать развиваться самостоятельно (гидры) или остаться прикрепленным к родительской особи (например, колониальные коралловые полипы). При другом способе вегетативного размножения — фрагментации родительская особь распадается на несколько частей, а-затем каждая из них восстанавливает недостающие части и превращается в целый организм. Так размножаются некоторые плоские черви.
У некоторых видов животных потомки могут развиваться из неоплодотворенных яйцеклеток. Этот вид бесполого размножения называется партеногенезом.
Преимущества бесполого размножения в его простоте и эффективности — не нужно находить партнера, потомство может оставить любая особь в любом месте, удачные сочетания генов, однажды появившись в результате мутаций, не теряются. При переходе к половому размножению эти преимущества утрачиваются. Однако в результате комбинации признаков отца и матери обеспечивается генетическое разнообразие потомства. Возникает новый неисчерпаемый источник генетической изменчивости. Появление оплодотворения — важное событие в эволюции размножения. Оно придает эволюционную пластичность популяции, что особенно необходимо в изменчивой среде.
Большинство высших форм животных размножаются в основном половым способом. В зависимости от отсутствия или наличия дифференциации (разделения) полов различные формы полового размножения можно отнести к двум типам — гермафродитному (обоеполому) и раздельнополому (см. рис.).
В половом размножении всегда участвуют гаметы противоположного пола — мужская и женская, которые у некоторых низших животных производятся одной особью — это герма-фродитный тип размножения. У большинства животных каждая особь производит только один тип гамет — это раздельнополый тип размножения (см. Пол). При оплодотворении гаплоидные гаметы разных особей сливаются и образуется диплоидная зигота, которая, делясь, дает начало новому организму. Лишь у некоторых видов в размножении могут участвовать гаметы одной особи (самооплодотворение).
Наиболее просто происходит оплодотворение у многих водных организмов. Особи противоположного пола выделяют гаметы в воду, где те находят друг друга и сливаются. Это наружное оплодотворение. Так размножаются многие кольчатые черви, пластинчатожаберные моллюски, иглокожие, многие рыбы, амфибии. Для повышения вероятности оплодотворения эти организмы производят гаметы в очень больших количествах. Например, луна-рыба мечет до 300 млн. икринок!
Более прогрессивно в эволюционном плане внутреннее оплодотворение, при котором процесс слияния гамет происходит в теле материнской особи. У животных для этих целей служат специальные копулятивные органы. Вероятность встречи гамет при внутреннем оплодотворении увеличивается, поэтому такие организмы вырабатывают значительно меньше яйцеклеток. Количество сперматозоидов остается достаточно большим. Внутреннее оплодотворение характерно для многих групп водных беспозвоночных, живородящих рыб, некоторых земноводных и для всех пресмыкающихся, птиц и млекопитающих.
У сухопутных форм зародыш может развиваться в теле самки, как у большинства млекопитающих. Такие животные называются живородящими (см. Живорождение). Иногда развитие протекает во внешней среде — в яйцах, покрытых скорлупой (насекомые, пресмыкающиеся, птицы, некоторые млекопитающие). Это яйцекладущие животные. Внутреннее оплодотворение и появление заботы о потомстве резко снижают смертность потомства у высших животных и повышают надежность размножения.
Если особь имеет и мужские и женские половые органы, то она способна скрещиваться с любой другой особью либо имеет способность к самооплодотворению. Это гермафродитное размножение. Многие низшие животные — гермафродиты. У видов, которым свойствен гермафродитизм, дифференциация (разделение) полов на уровне целого организма отсутствует (см. рис.). У них в размножении иногда может участвовать одна особь (например, у ленточных червей). Однако у большинства гер-мафродитных форм самооплодотворения не происходит.
Но в основном эволюция размножения шла в направлении раздельнополости. При раздельнополом способе размножения наряду со скрещиванием имеет место и разделение на два пола, причем скрещиваться между собой могут только особи противоположного пола. Большинство эволюционно прогрессивных форм (млекопитающие, птицы, насекомые, двудомные растения и др.) раздельнополы.
Некоторым животным, например кишечнополостным, свойственно чередование бесполого и полового поколений (см. рис. на с. 326), но большинство животных размножаются только половым способом и имеют специальные половые органы (см. Половая система).
Эволюция размножения шла, как правило, в направлении от бесполых форм к половым, от изогамии к анизогамии (см.
меты), от участия всех клеток в размножении к разделению клеток на соматические и половые, от наружного оплодотворения к внутреннему с внутриутробным развитием (см. Половая система) и заботой о потомстве. Темп размножения, численность потомства, частота смены поколений наряду с другими факторами определяют скорость приспособления вида к условиям среды. Например, высокие темпы размножения и частая смена поколений позволяют насекомым в короткий срок вырабатывать устойчивость к ядохимикатам. В эволюции позвоночных — от рыб до теплокровных — наблюдается тенденция к уменьшению численности потомства и увеличению его выживаемости.
Размножение у растений. Как и у животных, у растений имеются обе формы размножения: бесполое и половое. Несмотря на многообразие способов бесполого размножения, среди них можно выделить вегетативное и собственно бесполое.
Вегетативное размножение растений непосредственно связано со способностью их к регенерации. Вегетативно размножаются одноклеточные и многоклеточные, как гаплоидные, так и диплоидные растения. Многие одноклеточные водоросли вегетативно размножаются митотическим делением клетки на две. У одноклеточных водорослей с плотными клеточными оболочками (например, хламидомонад) новые особи образуются внутри делящейся клетки и выходят наружу после разрушения ее оболочки. Подобным образом внутри клеток нитчатых зеленых водорослей (например, улотрикса) формируются и выходят в воду специальные клетки, предназначенные для вегетативного размножения. По происхождению это митоспо-ры (см. Споры). Они прикрепляются к подводному субстрату, теряют жгутики и прорастают в новые нити водоросли. Другие нитчатые водоросли (например, свободно плавающая в толще воды спирогира) спор не образуют, а размножаются вегетативно при разрыве нитей на части, способные делиться и расти.
Многие многоклеточные низшие растения и талломные (слоевищные) высшие (некоторые печеночные мхи) размножаются вегетативно — делением таллома на участки. Есть у них и иные способы вегетативного размножения (например, выводковыми тельцами).
Многие папоротникообразные и покрытосеменные растения размножаются корневищами (см. Органы растений), старые участки которых отмирают, а молодые ответвления обособляются. У некоторых высших растений могут опадать с материнской особи и самостоятельно укореняться пазушные или придаточные (развивающиеся на листьях) почки. Есть покрытосеменные растения, которые размножаются луковицами (например, многие лилейные) .
На способности растений к регенерации основано искусственное вегетативное их размножение делением корневищ, луковицами, отводками, черенками, используемое в садоводстве. При вегетативном размножении развивающиеся самостоятельно (или привитые) отделившиеся или отделенные части растений сохраняют признаки материнской особи. Поэтому вегетативное размножение многих возделываемых растений имеет большое народнохозяйственное значение.
Собственно бесполое (мейоспорами) и половое размножение растений следует рассматривать вместе. При половом размножении теоретически возможное максимальное число потомков может быть равным лишь числу зигот, т. е. половине числа гамет (у изогамных гомо-талличных организмов), или числу тех гамет, которых меньше (у изогамных гетероталлич-ных, а также гетеро- и оогамных организмов; в двух последних случаях это женские гаметы). А максимальное число потомков при бесполом размножении определяется общим числом жизнеспособных мейоспор (см. также Пол, Оплодотворение).
Водоросли (например, диатомовые, некоторые бурые), в циклах развития которых гаплоидны лишь гаметы, размножаются только половым путем. Водоросли, у которых диплоидны лишь зиготы (например,улотрикс), размножаются как половым, так и бесполым путем. У немногих водорослей спорофит и гаметофит (бесполое и половое поколения, сменяющие друг друга) представлены одинаково; здесь последовательно чередуются оба способа размножения — бесполое и половое (см. Чередование поколений).
У большинства растений, в циклах развития которых выражено чередование поколений, обычно одно из них преобладает над другим. Например, у мхов преобладает гаметофит, а спорофит (спорогоний) развивается из зиготы на спорофите. Но главную роль в размножении играют мейоспоры, которых в коробочке спорогония образуется гораздо больше, чем яйцеклеток (см. Гаметы) в женских гаметангиях одной особи.
У папоротников спорофит преобладает над гаметофитом (заростком).
обоеполом заростке равноспорового папоротника образуется несколько женских гаметангиев с одной яйцеклеткой в каждом. Поскольку заросток — очень маленькое растение (2—3 мм в поперечнике), на нем обычно развивается лишь один спорофит, т. е. полового размножения практически не бывает. Среди разноспоровых папоротников есть такие, у которых на женском заростке развивается лишь один гаметангий с единственной яйцеклеткой. Здесь полового размножения быть не может, так как на одном гаметофите развивается тоже лишь один спорофит. Оплодотворение, которое происходит на заростке и не приводит к увеличению числа особей, называется в ботанике половым воспроизведением. Таким образом, для папоротников характерно бесполое размножение.
Отсутствие полового размножения (при сохранении полового воспроизведения) еще более четко выражено у семенных растений, га-метофиты которых развиваются на спорофитах. Это привело к возникновению самого совершенного у растений способа размножения — семенами. При семенном размножении максимальное число потомков определяется числом жизнеспособных мегаспор, а в конечном итоге — числом семязачатков, так как нуцеллус в семязачатке один, а в нем одна мегаспора.
Смена ядерных фаз в циклах развития организмов имеет огромное биологическое значение, поскольку и при мейозе, и при оплодотворении происходит перекомбинация аллелей. Для обеспечения оплодотворения гетеро- и оогамные организмы в общем всегда образуют значительно больше мужских гамет, чем женских. У разноспоровых растений (особенно у семенных) каждый из сильно редуцированных мужских заростков образует ограниченное число мужских гамет. Но общее число этих гамет огромно, поскольку каждый из спорофитов, развивающий микроспорангии, образует множество микроспор, дающих начало мужским заросткам.
Однако у некоторых растений обнаруживается так называемый апомиксис — нарушение чередования ядерных фаз, проявляющееся в выпадении из циклов развития либо оплодотворения и мейоза, либо только оплодотворения.
У бактерий также может происходить процесс, напоминающий половое размножение. При этом две разные клетки обмениваются генетическим материалом.
Источник: yunc.org
Большинство организмов, обычно размножающихся бесполым путем, способно к половому размножению. При этом ряд поколений с бесполым размножением сменяется поколением особей, размножающихся с помощью гамет или же осуществляющих половой процесс. Вслед за этим вновь наблюдается бесполое размножение. Смена (чередование) половых и бесполых поколений происходит у разных видов с разной периодичностью, регулярно или через неодинаковые промежутки времени.
Первичное чередование поколений заключается в смене полового размножения спорообразованием. Оно наблюдается у представителей классов споровиков, жгутиконосцев, некоторых растений и отражает сохранение в филогенезе соответствующих организмов как более древней (бесполой), так и более прогрессивной (половой) форм размножения. Вторичное чередование поколений заключается в переходе на некоторых стадиях жизненного цикла к бесполому или партеногенетическому размножению животных, освоивших половое размножение. Оно распространено у кишечнополостных, членистоногих.
Включение в цикл развития организмов, размножающихся преимущественно бесполым путем, полового поколения время от времени активизирует комбинативную изменчивость и этим способствует преодолению генетического однообразия потомков, расширяя эволюционные и экологические перспективы группы.
ПОЛОВЫЕ КЛЕТКИ
По сравнению с другими клетками функция гамет уникальна. Они обеспечивают передачу наследственной информации между особями разных поколений, чем сохраняют жизнь во времени. Был период в биологии, когда половые и соматические клетки противопоставляли друг другу, наделяя лишь первые всей полнотой свойств жизни, проносимых ими через поколения. В настоящее время экспериментально доказана возможность развития полноценного организма на основе наследственной информации ядра дифференцированной соматической клетки, например кишечного эпителия (рис. 5.3).
В 1990-х годах группе английских ученых, использовавших принципиально аналогичный подход, удалось вырастить (клонировать) овцуДолли. Это был пример успешного решения задачи получения взрослой особи на основе генетического материала соматической клетки у млекопитающих.
помним, что клоны представлены генетически идентичными клетками, образовавшимися путем последовательных митотических делений исходно одной клетки-родоначальницы и ее потомков. Вытекающая из этого техническая возможность клонирования людей порождает серьезные морально-этические проблемы, так как допускает преднамеренное «тиражирование» избранного человека с присущим только ему набором физических психо-эмоциональных свойств и даже интеллектуального потенциала. В настоящее время общественностью, в том числе и научной, признается недопустимым клонирование людей. Вместе с тем, использование технологии клонирования для получения достаточного числа клеток определенного типа, генетически неотличимых от клеток данного организма и, следовательно, иммунологически с ним совместимых, рассматривается как перспективное направление в области трансплантации тканей.

Рис. 5.3. Опыт, показывающий функциональную полноценность наследственного материала дифференцированной клетки:
1 — яйцеклетка с убитым УФ-лучами ядром — источник цитоплазмы, 2 — эпителиальные клетки кишечника головастика — источник наследственного материала, 3 — ядро, 4 — пересадка ядра из эпителиальной клетки в яйцеклетку, 5 — головастик, 6 — лягушка
Гаметы представляют собой одно из многих направлений дифференцировки клеток многоклеточного организма. Они образуют «клеточную линию», специализированную к выполнению репродуктивной функции. Предполагают, что клетки указанной линии возникают из бластомеров, имеющих на вегетативном полюсе цитоплазму особого рода — так называемую зародышевую плазму1, богатую РНК.
В сравнении с другими линиями соматических клеток (эпителиальные, нервные, мышечные) гаметы характеризуются рядом отличий. Важнейшее из них — гаплоидный набор хромосом в ядрах, что обеспечивает воспроизведение в зиготе типичного для организмов данного вида диплоидного числа хромосом.
Действительно, оплодотворение сперматозоидом яйцеклетки, ядра которых содержат по 23 хромосомы, обусловливает формирование зиготы с 46 хромосомами, что типично для соматических клеток человека. Гаметы отличаются необычным для других клеток значением ядерно-цитоплазматического отношения. У яйцеклеток оно снижено благодаря увеличенному объему цитоплазмы, в которой размещен питательный материал (желток) для развития зародыша. У сперматозоидов благодаря малому количеству цитоплазмы ядерно-цитоплазматическое отношение высокое. Это находится в соответствии с главной функциональной задачей мужской гаметы — транспортировкой наследственного материала к яйцеклетке.
Половые клетки отличаются низким уровнем обменных процессов, близким к состоянию анабиоза. Мужские гаметы не вступают в митотический цикл. У яйцеклеток эта способность восстанавливается при оплодотворении или действии фактора, активирующего партеногенез.
По ряду признаков женские и мужские гаметы отличаются друг от друга, что связано с различными функциями яйцеклетки и сперматозоида в процессе размножения. Яйцеклетки имеют оболочки, которые выполняют защитную функцию, обеспечивают требуемый уровень обмена веществ, препятствуют проникновению в яйцеклетку более одного спермия, способствуют внедрению (имплантации) зародыша в стенку матки у плацентарных животных, поддерживают форму зародыша.
Для яйцеклетки характерна плазматическая сегрегация. После оплодотворения (у асцидий уже через 5 мин) в еще не дробящемся яйце происходит закономерное перераспределение цитоплазмы. В дальнейшем цитоплазма разного состава также закономерно распределяется по клеткам тканей разных зачатков. По-видимому, на ранних стадиях способность бластомеров развиваться в определенном направлении зависит от наследования ими веществ, концентрирующихся в разных участках цитоплазмы яйцеклетки.
Сперматозоид имеет аппарат движения в виде жгутика. В семенной жидкости мужская гамета человека развивает скорость до 5 см/ч. Поясним приводимую цифру следующим примером. Если учесть соотношение преодолеваемого расстояния и длины движущегося объекта, то при названной скорости сперматозоид человека перемещается в 1,5 раза быстрее, чем пловец олимпийского ранга. Яйцеклетка, лишенная аппарата активного движения, преодолевает расстояние до полости матки, равное примерно 10 см, за 4—7 сут. Сперматозоиды некоторых видов животных имеют акросомный аппарат, выбрасывающий при контакте с яйцеклеткой длинную нить. Он обеспечивает проникновение ядра мужской гаметы в цитоплазму яйцеклетки путем растворения особыми ферментами ее оболочек. Описаны и другие приспособления, способствующие оплодотворению.
Гаметогенез
Гаметогенез — процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) —подразделяется наряд стадий (рис. 5.4).
В стадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониями и овогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Размножение овогоний приурочено главным образом к периоду эмбриогенеза. У человека в женском организме этот процесс наиболее интенсивно протекает в яичниках между 2-м и 5-м месяцами внутриутробного развития. К 7-му месяцу большая часть овоцитов входит в профазу I мейоза.
Так как способом размножения клеток-предшественниц женских и мужских гамет является митоз, то овогоний и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. В ходе митотического цикла их хромосомы имеют либо однонитчатую (после митоза и до завершения синтетического периода интерфазы), либо двунитчатую (постсинтетический период, профаза и метафаза митоза) структуру в зависимости от количества биспиралей ДНК. Если в одинарном, гаплоидном наборе число хромосом обозначить как п, а количество ДНК — как с, то генетическая формула клеток в стадии размножения соответствует 2п2с до S-периода и 2n4c после него.
Рис. 5.4. Схема гаметогенеза:
1 — сперматогенез, 2 — овогенез, n — количество хромосомных наборов,
с — количество ДНК, РТ — редукционные тельца
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоциты и овоциты I порядка, причем последние достигают больших размеров, чем первые. Одна часть накапливаемых веществ представляет собой питательный материал (желток в овоцитах), другая — связана с последующими делениями. Важным событием этого периода является репликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и овоцитов I порядка приобретает вид 2n4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное, которые вместе составляют мейоз (см. разд. 5.3.2). После первого деления образуются сперматоциты и овоциты II порядка (формула n2с), а после второго — сперматиды и зрелая яйцеклетка (пс).
В результате делений на стадии созревания каждый сперматоцит I порядка дает четыре сперматиды, тогда как каждый овоцит I порядка — одну полноценную яйцеклетку и редукционные тельца, которые в размножении не участвуют. Благодаря этому в женской гамете концентрируется максимальное количество питательного материала — желтка.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра, образуя акросомный аппарат, играющий большую роль в оплодотворении. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена.
Мейоз
Центральным событием гаметогенеза является особая форма клеточного деления — мейоз. В отличие от широко распространенного митоза, сохраняющего в клетках постоянное диплоидное число хромосом, мейоз приводит к образованию из диплоидных клеток гаплоидных гамет. При последующем оплодотворении гаметы формируют организм нового поколения с диплоидным кариотипом (пс + пс == 2n2c). В этом заключается важнейшее биологическое значение мейоза, который возник и закрепился в процессе эволюции у всех видов, размножающихся половьм путем (см. разд. 3.6.2.2).
Мейоз состоит из двух быстро следующих одно за другим делений, происходящих в периоде созревания. Удвоение ДНК для этих делений осуществляется однократно в периоде роста. Второе деление мейоза следует за первым практически сразу так, что наследственный материал не синтезируется в промежутке между ними (рис. 5.5).
Первое мейотическое деление называют редукционным, так как оно приводит к образованию из диплоидных клеток (2п2с) гаплоидных клеток п2с. Такой результат обеспечивается благодаря особенностям профазы первого деления мейоза. В профазе I мейоза, так же как в обычном митозе, наблюдается компактная упаковка генетического материала (спирализация хромосом). Одновременно происходит событие, отсутствующее в митозе: гомологичные хромосомы конъюгируют друг с другом, т.е. тесно сближаются соответствующими участками.
В результате конъюгации образуются хромосомные пары, или биваленты, числом п. Так как каждая хромосома, вступающая в мейоз, состоит из двух хроматид, то бивалент содержит четыре хроматиды. Формула генетического материала в профазе I остается 2n4c. К концу профазы хромосомы в бивалентах, сильно спирализуясь, укорачиваются. Так же как в митозе, в профазе I мейоза начинается формирование веретена деления, с помощью которого хромосомный материал будет распределяться между дочерними клетками (рис. 5.5).
Рис. 5.5. Стадии мейоза
Отцовские хромосомы обозначены черным цветом, материнские —неокрашснные. На рисунке не изображена метафаза I, в которой биваленты располагаются в плоскости экватора веретена деления, и телофаза I, быстро переходящая в профазу II
Процессы, происходящие в профазе I мейоза и определяющие его результаты, обусловливают более продолжительное течение этой фазы деления по сравнению с митозом и дают возможность выделить несколько стадий в ее пределах (рис. 5.5).
Лептотена —наиболее ранняя стадия профазы I мейоза, в которой начинается спирализация хромосом, и они становятся видимыми в микроскоп как длинные и тонкие нити. Зиготена характеризуется началом конъюгации гомологичных хромосом, которые объединяются синаптонемальным комплексом в бивалент (рис. 5.6). Пахитена — стадия, в которой на фоне продолжающейся спирализации хромосом и их укорочения, между гомологичными хромосомами осуществляется кроссинговер — перекрест с обменом соответствующими участками. Диплотена характеризуется возникновением сил отталкивания между гомологичными хромосомами, которые начинают отдаляться друг от друга в первую очередь в области центромер, но остаются связанными в областях прошедшего кроссинговера — хиазмах (рис. 5.7).
Диакинез — завершающая стадия профазы I мейоза, в которой гомологичные хромосомы удерживаются вместе лишь в отдельных точках хиазм. Биваленты приобретают причудливую форму колец, крестов, восьмерок и т.д. (рис. 5.8).
Рис. 5.6. Образование бивалентов конъюгирующими хромосомами в зиготене:
1 — центрмеры
Таким образом, несмотря на возникающие между гомологичными хромосомами силы отталкивания, в профазе I не происходит окончательного разрушения бивалентов. Особенностью мейоза в овогенезе является наличие специальной стадии — диктиотены, отсутствующей в сперматогенезе. На этой стадии, достигаемой у человека еще в эмбриогенезе, хромосомы, приняв особую морфологическую форму «ламповых щеток», прекращают какие-либо дальнейшие структурные изменения на многие годы. По достижении женским организмом репродуктивного возраста под влиянием лютеинизирующего гормона гипофиза, как правило, один овоцит ежемесячно возобновляет мейоз.
В метафазе I мейоза завершается формирование веретена деления. Его нити прикрепляются к центромерам хромосом, объединенных в биваленты, таким образом, что от каждой центромеры идет лишь одна нить к одному из полюсов веретена. В результате нити, связанные с центромерами гомологичных хромосом, направляясь к разным полюсам, устанавливают бивалентны в плоскости экватора веретена деления.
Рис. 5.7. Стадии диплотены в мейозе кузнечика
В анафазе I мейоза ослабляются связи между гомологичными хромосомами в бивалентах и они отходят друг от друга, направляясь к разным полюсам веретена деления. При этом к каждому полюсу отходит гаплоидный набор хромосом, состоящих из двух хроматид (см. рис. 5.5).
Рис. 5.8. Стадия диакинеза в мейозе у человека.
Стрелками указаны хиазмы
В телофазе I мейоза у полюсов веретена собирается одинарный, гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК.
Формула генетического материала образующихся дочерних клеток соответствует п2с.
Второе мейотическое (эквационное) деление приводит к образованию клеток, в которых содержание генетического материала в хромосомах будет соответствовать их однонитчатой структуре пс (см. рис. 5.5). Это деление протекает, как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом. В процессе такого деления материнские двунитчатые хромосомы, расщепляясь, образуют дочерние однонитчатые.
Одна из главных задач мейоза — создание клеток с гаплоидным набором однонитчатых хромосом —достигается благодаря однократной редупликации ДНК для двух последовательных делений мейоза, а также благодаря образованию в начале первого мейотического деления пар гомологичных хромосом и дальнейшего их расхождения в дочерние клетки.
Процессы, протекающие в редукционном делении, обеспечивают также не менее важное следствие — генетическое разнообразие гамет, образуемых организмом. К таким процессам относят кроссинговер, расхождение гомологичных хромосом в разные гаметы и независимое поведение бивалентов в первом мейотическом делении (см. разд. 3.6.2.3).
Кроссинговер обеспечивает перекомбинацию отцовских и материнских аллелей в группах сцепления (см. рис. 3.72). Ввиду того что перекрест хромосом может происходить в разных участках, кроссинговер в каждом отдельном случае приводит к обмену разным по количеству генетическим материалом. Необходимо отметить также возможность возникновения нескольких перекрестов между двумя хроматидами (рис. 5.9) и участия в обмене более чем двух хроматид бивалента (рис. 5.10). Отмеченные особенности кроссинговера делают этот процесс эффективным механизмом перекомбинации аллелей.
Рис. 5.9. многократный кроссинговер между гомологичными хромосомами:
А—Е, а—е — локусы хромосом
Расхождение гомологичных хромосом в разные гаметы в случае гетерозиготности приводит к образованию гамет, различающихся по аллелям отдельных генов (см. рис. 3.74).
Случайное расположение бивалентов в плоскости экватора веретена деления и последующее их расхождение в анафазе I мейоза обеспечивают перекомбинацию родительских групп сцепления в гаплоидном наборе гамет (см. рис. 3.75).
Рис. 5.10. Множественный обмен участками на стадии четырех хроматид
в пахитене мейоза
В кроссинговер могут вступать все четыре хроматиды бивалента, латинскими буквами обозначены мутантные аллели; знаком «+» —нормальные аллели
Последние стадии овогенеза воспроизводятся и вне организма женщины, в искусственной питательной среде. Это позволило осуществить зачатие человека «в пробирке». Перед овуляцией хирургическим путем яйцо извлекается из яичника и переносится в среду со сперматозоидами. Возникающая в результате оплодотворения зигота, будучи помещена в подходящую среду, осуществляет дробление. На стадии 8—16 бластомеров зародыш переносится в матку женщины-реципиента, которая осуществляет донашивание и роды. Число успешных результатов такого переноса в последнее время возрастает.
Гаметогенез отличается высокой производительностью. За время половой жизни мужчина продуцирует не менее 500 млрд. сперматозоидов. На пятом месяце эмбриогенеза в зачатке женской половой железы насчитывается 6—7 млн. клеток-предшественниц яйцеклеток. К началу репродуктивного периода в яичниках обнаруживается примерно 100 000 овоцитов. От момента полового созревания до прекращения гаметогенеза в яичниках созревает 400—500 овоцитов.
Источник: megaobuchalka.ru
Бесполое размножение
При бесполом делении, где задействована только одна особь, процесс размножения происходит без формирования гамет. Потомство формируется за счет отпочковывания от материнского организма или откладывания в спецорганах.
Существуют следующие виды бесполого размножения:
Деление — преимущественно встречается у простейших организмов, при этом исходная материнская клетка разделяется на две части, образуя идентичное дочернее поколение.
Выделяют следующие подвиды:
- Деление надвое — свойственно доядерным видам;
- митотическое деление — встречается у простейших;
- множественное деление — типичное явление для малярийного плазмодия.
Почкование — характеризуется формированием дочерних организмов в виде выпячиваний на материнском теле. После созревания они отделяются от тела родителя и развиваются дальше самостоятельно. Если дочерние формы не отпочковываются и сохраняют связь с материнским организмом, формируются колонии (представители типа стрекающие).
Фрагментация — процесс, при котором зрелые особи развиваются из отдельных частей тела взрослой особи (первичноротые, хирофитовые водоросли, водяная чума). Фрагментация возможна благодаря регенеративным способностям организма.
Полиэмбриония — новые особи формируются при разделении зародыша на несколько частей (однояйцевые близнецы).
Вегетативное размножение — зарождение новых особей идет из отдельных органов материнского организма. Формирование молодого растения возможно из корневой системы, ветвей или листьев (редко).
Корень служит основой для образования придаточных почек, из которых развиваются надземные побеги. Новообразованные побеги крепятся к почве с помощью добавочных корней. После гибели материнского корня, проросшие растения начинают самостоятельную жизнь.
Вегетативное размножение способствует быстрому распространению калины, осота, кипрея узколистого. Растения из семейства бобовые или рода Вербейник размножаются при помощи поверхностных побегов, которые стелются по земле и в местах соприкосновения почвы и узлов побегов прорастают добавочные корни. Так растение начинает самостоятельное развитие.
Спорообразование — характерно для некоторых простейших и растений, которые могут образовывать споры. Споровые клетки, попадая во влажную среду, развиваются, достигая зрелости. Формирование споровых клеток идет в спорангиях – специальных органах покрытосеменных растений. У грибов и водорослей споры образуются из всех клеток тела.
Клонирование — один из видов бесполого размножения, применяемый учеными для копирования исходного генетического материала. Так получают идентичные копии с материнских особей.
Роль бесполого размножения
Организмы, которые размножаются бесполым путем, хорошо адаптируются к плавно изменяющимся условиям окружающей среды. Их потомство всегда множественное, быстро созревает и начинает тоже делиться, что способствует росту численности популяции. Хорошо известные виды с бесполым размножением: гидра, амеба, дрожжевые грибы.
Все клетки нашего организма постоянно обновляются, это возможно благодаря бесполому размножению. Соматические клетки делятся в процессе митоза.
Учитывая быстрые темпы созревания и деления, растения и животные, которые делятся бесполым путем, часто используются селекционерами.
Половое размножение
Половое размножение осуществляется при взаимодействии пары разнополых особей. Они имеют репродуктивную систему, где формируются половые клетки — гаметы. Для женских особей характерно образование яйцеклеток, а для мужских — сперматозоидов.
Формирование половых клеток называют гаметогенезом, главным моментом их образования является мейоз. Во время слияния гамет происходит оплодотворение и зарождения новой жизни. Сформированная зигота не будет точной версией родителей, потому что в процессе мейоза идет перестройка генетической информации.
Гаметы у разных представителей отличаются между собой, поэтому выделяют следующие формы полового размножения: гомогамию, анизогамию и оогамию.
Гомогамия — разделение гамет на мужские и женские является условностью, потому что половые клетки организмов разных полов имеют идентичное строение и форму.
Анизогамия — половые клетки обеих полов могут передвигаться. Яйцеклетки превосходят по размеру сперматозоиды, но они почти неподвижны.
Оогамия — женские гаметы не могут самостоятельно перемещаться и намного больше мужских.
Оогамия встречается наиболее часто, свойственна многим представителям животного и растительного мира. Гомогамия и анизогамия присуща для простейших видов (фотоавтотрофные одноклеточные).
Отдельные виды водорослей и грибов могут размножаться, не образуя гамет, такие формы деления называются: гологамия и конъюгация.
В процессе гологамии одноклеточные особи с одинарным набором хроматид сливаются между собой, тем самым берут на себя роль половых клеток. Новообразованная зигота следом делится мейотическим путем, образуя 4 гаплоидные особи.
Деление путем конъюгации характерно для грибов, при этом идет слияние между гаплоидными клетками нитей таллома. После обмена информацией формируются диплоидные зародышевые клетки.
Роль полового размножения
Половое размножение важное природное явление, обеспечивающее высокий уровень изменчивости и дает возможность выживать в резко меняющихся условиях. Именно поэтому половое размножение превосходит бесполое, при котором все оставленное потомство унаследует точную копию генома родителей.
При половом делении идет перестройка генетического кода, что проявляется разнообразием признаков у потомков. Возникновение новых характеристик, адаптационных механизмов является основой для эволюционных процессов. Поэтому половое размножение занимает главное положение в природе.
Источник: animals-world.ru
по теме: «Строение прокариотической клетки»
Вариант №1
Выпишите номера верных утверждений
по теме: «Строение прокариотической клетки»
Вариант №2
1)Антони ван Левенгук ввел термин «бактерия»
2)Эренберг Готфрид ввел термин «бактерия»
3)Бактерии произошли 1,5млрд.лет назад
4)Бактерии погибают при незначительных изменениях в температуре
5)Кокки — это эукариоты
6)Муреин — это углевод в клеточной стенке прокариот
7)В прокариотической клетке содержится митохондрия, как источник энергии
8)Генетический материал — это кольцевая молекула ДНК
9)Автотрофы это организмы, которые используют готовые органические вещества
10)Аэробам кислород губителен
11)Представитель патогенных бактерий — молочнокислая бактерия
12)Выросты на бактериальной клетке — это пили
13)Прокариоты размножаются только бесполым путем
14)Прокариоты при бесполом размножении делятся надвое каждые 20-30 минут
15)Наука о бактериях — цитология
16)Бактерии принимают участие в формировании и плодородии почв
17)Холера, чума, дифтерия — это заболевания, которые вызваны одноклеточными животными
18)Приспособление к выживанию в неблагоприятных условиях — это спорообразование
19)Аэробам — кислород губителен
20)Сапрофиты — питаются органическими веществами отмерших организмов.
ОТВЕТЫ:
1)Антони ван Левенгук открыл одноклеточных животных
2)Луи Пастер изучил болезнетворные свойства бактерий
3)Роберт Кох изучил бактерию — туберкулеза
4)Бактерии- одноклеточные организмы, имеющие клеточное ядро
5)Бактерии не могут заселять атмосферу
6)Бактерии имеющие форму шара — это кокки
7)Снаружи бактерии покрыты только оболочкой
8)Питательные вещества — белок, бактерия получает из рибосом
9)Прокариоты передвигаются только благодаря слизи
10)Плазматические выпячивания у мембраны — это мезосомы
11)Гетеротрофы — это организмы, создающие питательные вещества за счет энергии солнечного света
12)В мезосомах находятся ферменты, ускоряющие скорость химических реакций
13)Аэробы — кислород вреден
14)Аэробы — используют кислород в энергетических обменах
15)Пили — это палочковидные бактерии
16)Патогенные бактерии питаются органическими веществами живых организмов
17)Бесполое размножение наступает только при неблагоприятных условиях
18)Спора — это приспособление к выживанию в неблагоприятных условиях.
19)Половой процесс у прокариот отсутствует
20)Бактерии принимают участие в образовании ископаемых и разрушении погибших животных и растений
Источник: himia.neznaka.ru
Бесполое размножение
Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение.
Деление — способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить: а) простое бинарное деление (прокариоты), б) митотическое бинарное деление (простейшие, одноклеточные водоросли), в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время деления парамеции (1) микронуклеус делится митозом, макронуклеус — амитозом. Во время шизогонии (2) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов.
Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
Фрагментация (4) — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
| Вегетативный орган | Способ вегетативного размножения | Примеры |
|---|---|---|
| Корень | Корневые черенки | Шиповник, малина, осина, ива, одуванчик |
| Корневые отпрыски | Вишня, слива, осот, бодяк, сирень | |
| Надземные части побегов | Деление кустов | Флокс, маргаритка, примула, ревень |
| Стеблевые черенки | Виноград, смородина, крыжовник | |
| Отводки | Крыжовник, виноград, черемуха | |
| Подземные части побегов | Корневище | Спаржа, бамбук, ирис, ландыш |
| Клубень | Картофель, седмичник, топинамбур | |
| Луковица | Лук, чеснок, тюльпан, гиацинт | |
| Клубнелуковица | Гладиолус, крокус | |
| Лист | Листовые черенки | Бегония, глоксиния, колеус |
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
Клонирование — комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон — совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
Половое размножение
Половое размножение осуществляется при участии двух родительских особей (мужской и женской), у которых в особых органах образуются специализированные клетки — гаметы. Процесс формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз. Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния мужской и женской гамет. Процесс слияния мужской и женской гамет называется оплодотворением. Обязательным следствием полового размножения является перекомбинация генетического материала у дочернего поколения.
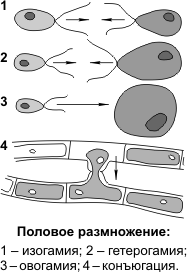
В зависимости от особенностей строения гамет, можно выделить следующие формы полового размножения: изогамию, гетерогамию и овогамию.
Изогамия (1) — форма полового размножения, при которой гаметы (условно женские и условно мужские) являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия (2) — форма полового размножения, при которой женские и мужские гаметы являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия (3) — форма полового размножения, при которой женские гаметы неподвижные и более крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями.
Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет. Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных организмов. При конъюгации (4) происходит слияние содержимого отдельных гаплоидных клеток нитевидных талломов. По специально образующимся каналам содержимое одной клетки перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также делится мейозом.
Источник: licey.net