Урикотелические животные
- АНАЭРОБНЫЕ ОРГАНИЗМЫ — анаэробы , организмы, способные жить и развиваться при отсутствии в среде свободного кислорода. Термин «анаэробы» ввёл Л. Пастер, открывший в 1861 бактерии маслянокислого брожения…
- АВТОТРОФНЫЕ ОРГАНИЗМЫ — автотрофы , организмы, использующие для построения своего тела CO2 в качестве единственного или главного источника углерода, т. е. синтезирующие необходимые для жизнедеятельности органич. в-ва из неорганических…
- АНАЭРОБНЫЕ ОРГАНИЗМЫ — анаэробы , организмы, не нуждающиеся для нормальной жизнедеятельности в присутствии кислорода…
- АВТОТРОФНЫЕ ОРГАНИЗМЫ — автотро́фные органи́змы , организмы, синтезирующие из неорганических веществ необходимые для жизни органические вещества…
- АЦИДОФИЛЬНЫЕ ОРГАНИЗМЫ — организмы, нормально развивающиеся только на кислом субстрате …
- Анаэробные организмы — организм, способный жить в бескислородной среде; эта способность называется анаэробиозом и относится к бактериям, некоторым червям и моллюскам …
- АВТОТРОФНЫЕ ОРГАНИЗМЫ — автотрофы , организмы, использующие для построения своего тела С02 в качестве единственного или гл. источника углерода и обладающие как системой ферментов для ассимиляции СО2, так и способностью синтезировать все…
- АПОХЛОРОТИЧЕСКИЕ ОРГАНИЗМЫ — бесхлорофилльные организмы, утратившие хлорофилл благодаря переходу на питание органическими веществами…
- АЦИДОФИЛЬНЫЕ ОРГАНИЗМЫ — способны существовать в условиях значит. кислотности среды…
- АДВЕНТИВНЫЕ ОРГАНИЗМЫ — растения, животные, преднамеренно или случайно завезенные человеком из других климатических зон в новую для них область. См. также Акклиматизация…
- Аллотропные организмы — В ботанике так называются организмы, не способные пользоваться углекислотой, как источником углерода для построения органических веществ. Им противополагаются аутотрофные организмы , обладающие этой способностью…
- Аутотрофные организмы — организмы, способные вырабатывать потребное для них количество органических веществ самостоятельно из веществ минеральных. Такие организмы, очевидно, не зависят в своем существовании от других организмов…
- Организмы — …
- Автотрофные организмы — аутотрофные организмы, синтезирующие из неорганических веществ необходимые для жизни органические вещества…
- Аутотрофные организмы — то же, что Автотрофные организмы…
- АУТОТРОФНЫЕ ОРГАНИЗМЫ — Зеленые растения, нитробактерии, способные вырабатывать потребные для них органические вещества самостоятельно из веществ минеральных. Противопол. аллотрофным…
Источник: slovar.wikireading.ru
Виды конечных продуктов азотистого обмена у животных разных групп. В чем причины, определяющие их различия При расщеплении белков, нуклеиновых кислот и других азотсодержащих соединений образуются токсичные вещества — аммиак, мочевина и мочевая кислота, токсический эффект которых соответственно снижается в приведенном ряду. В зависимости от того, в какой из этих трех форм преимущественно выделяется азот, животных подразделяют на три группы: аммониотелические "font-size:10pt;"> урикотелические (выделяющие мочевую кислоту).
Форма выделения продуктов азотистого обмена тесно связана с условиями жизни животного и обеспеченностью водой. Аммиак весьма токсичен даже в малых концентрациях. Благодаря хорошей растворимости и небольшому молекулярному весу, он легко диффундирует через любую поверхность, соприкасающуюся с водой. Аммиак является конечным продуктом азотистого обмена у водных беспозвоночных, костных рыб, личинок и постоянно живущих в воде земноводных.
Наземные животные ограничены в воде: чтобы избежать накопления аммиака в тканях и жидкостях тела, они должны преобразовать его в конечные продукты, нетоксичные для организма. Наземные ресничные черви, земноводные, млекопитающие выделяют мочевину.
Низкая растворимость мочевой кислоты, выпадение ее в осадок делает ее осмотически неактивной. Для ее выведения из организма вода практически не нужна. Урикотелия в основном характерна для животных, освоивших наземную, в том числе и засушливую, среду (наземные насекомые, чешуйчатые пресмыкающиеся, птицы).
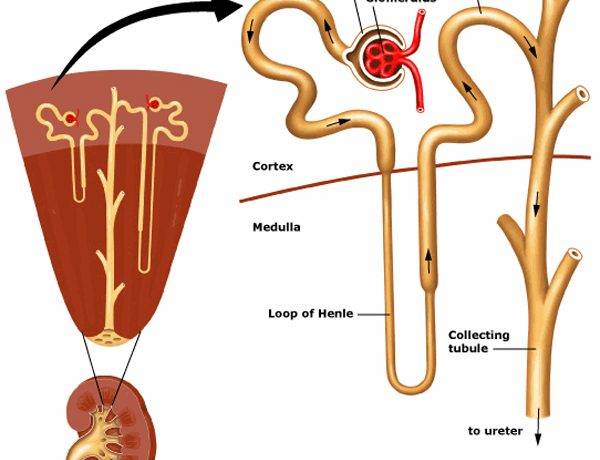
Водно-солевой обмен рыб
Почки рыб выводят аммиак, соли, воду; почки наземных позвоночных — мочевину, мочевую кислоту, соли, воду.Выделительная система рыб служит для выведения из организма продуктов обмена и обеспечения его водно-солевого состава. Она включает:
Основную массу туловищной почки заполняют нефроны. Нефрон состоит из:
1) мальпигиева тельца (клубочек капиллярных сосудов, заключенный в боуменову капсулу);
2) выделительного канальца.
Артериальная кровь по почечным артериям поступает в сосудистые клубочки, где фильтруется и образуется первичная моча. В средней части выделительных канальцев происходит обратное всасывание полезных для организма веществ (сахара, витамины, аминокислоты, вода) и образуется вторичная, или окончательная, моча. У хрящевых рыб основным компонентом мочи является мочевина, у костистых — аммиак (аммиак намного токсичнее мочевины).
Выделение продуктов распада тесно связано с водно-солевым обменом рыб. У морских и пресноводных рыб эти процессы протекают различно.
When a fish eats protein, like the peel of an orange part of it goes unused and becomes waste. As Dave McShaffrey, professor of biology at Marietta College in Ohio, explains on the college website, “When proteins are converted to carbohydrates to provide energy, the amino group is removed and must be dealt with.” In saltwater fish, this nitrogen-rich waste is usually converted to either ammonia or urea, which is one of the main excretory products of saltwater fish.
monia is easier to produce, but urea is less toxic, requires less water and gets rid of twice as much nitrogen. Theword “urine” is related to “urea.”
Морские хрящевые рыбы живут в изотоничной среде (т.е. осмотическое давление крови и тканевых жидкостей равно давлению окружающей среды). У них изотоничность внутренней и внешней среды обеспечивается за счет удержания в крови и тканевых жидкостях мочевины и солей (концентрация мочевины в крови у них достигает 2,6%). Через почки наружу у них выводятся лишь излишки мочевины, солей и воды, количество выделяемой мочи невелико (2-50 мл на 1 кг массы тела в сутки). У морских хрящевых рыб для выведения избытка солей сформировалась особая ректальная железа, открывающаяся в прямую кишку.
Все пресноводные рыбы живут в гипотонической среде (т.е. осмотическое давление крови и тканевых жидкостей выше, чем в окружающей среде), поэтому вода постоянно проникает в организм через кожу, жабры, с пищей. Чтобы избежать обводнения пресноводные рыбы имеют хорошо развитый фильтрационный аппарат почек, что позволяет им выделять большое количество мочи (50-300 л на 1 кг массы тела в сутки). Потеря солей с мочой компенсируется активной их реабсорбцией в почечных канальцах и поглощением солей жабрами из воды, часть солей поступает с пищей.
Морские костистые рыбы живут в гипертонической среде (т.е. осмотическое давление крови и тканевых жидкостей ниже, чем в окружающей среде), поэтому вода выходит из организма через кожу, жабры, с мочой и фекалиями. Во избежание иссушения они пьют соленую воду (от 40 до 200 мл на 1 кг массы в сутки), которая из кишечника всасывается в кровь. У морских костистых рыб уменьшается число клубочков в почках, а у некоторых рыб исчезают совсем (морская игла, морской черт). Таким образом, почки выводят небольшое количество мочи (0,5—20 мл на 1 кг массы тела в сутки).
Проходные рыбы при переходе из одной среды в другую могут изменять способ осморегуляции: в морской среде она осуществляется как у морских рыб, а в пресной — как у пресноводных. Такие адаптации водно-солевого обмена позволили костистым рыбам широко освоить пресные и соленые водоемы.
Адаптации наземных животных к выделению веществ
According to New World Encyclopedia, reptiles use two small kidneys as tools for excretion. The kidneys serve to filter the nitrogen from the animal’s bloodstream, then turn it into waste. The nitrogen then exits the body in dry form as uric acid crystals along with the feces. According to Stanford University, the kidneys in a bird also function as a means to remove nitrogen from the blood. The white substance found in bird droppings is actually uric acid, which is not water soluble. In both reptiles and birds, eliminating the nitrogen requires that the body exerts a great deal of energy. Both species are able to efficiently remove the nitrogen while losing very little water in the waste product.
Адаптации пустынных животных к выделению веществ
Обитатели полупустынных биотопов большую часть воды получают, поедая сочные части суккулентных растений.
кожно-легочные потери воды минимальны. Так, при температуре 20°С они достигают у относительно влаголюбивого вида — гребенщиковой песчанки 170 см3, а у сухолюбивой большой песчанки — только 50 см3 на 1 кг массы за 1 ч. Настоящие пустынные млекопитающие способны поедать почти сухие корма и практически не пить в течение всей жизни, удовлетворяя свои потребности лишь за счет образующейся в организме метаболической воды. Верблюды в кормные и влажные сезоны запасают жир, расходуемый в малокормное и сухое время — при этом образуется некоторое количество воды; наконец, во время отдыха и сна они снижают температуру тела, что также сокращает расход воды.
Desert Animals
Coping with water loss is a particular problem for animals that live in dry conditions. Some, like the camel, have developed great tolerance for dehydration. For example, under some conditions, camels can withstand the loss of one third of their body mass as water. They can also survive wide daily changes in temperature. This means they do not have to use large quantities of water in sweat to cool the body by evaporation.Smaller animals are more able than large ones to avoid extremes of temperature or dry conditions by resting in sheltered more humid situations during the day and being active only at night.The kangaroo rat is able to survive without access to any drinking water at all because it does not sweat and produces extremely concentrated urine.
ter from its food and from chemical processes is sufficient to supply all its requirements.

Какой из нефронов принадлежит верблюду, а какой – рептилии? Почему вы сделали такой выбор?
Fresh Water Fish
Although the skin of fish is more or less waterproof, the gills are very porous. The body fluids of fish that live in fresh water have a higher concentration of dissolved substances than the water in which they swim. In other words the body fluids of fresh water fish are hypertonic to the water (see chapter 3). Water therefore flows into the body by osmosis. To stop the body fluids being constantly diluted fresh water fish produce large quantities of dilute urine.
Marine Fish
Marine fish like the sharks and dogfish have body fluids that have the same concentration of dissolved substances as the water (isotonic) have little problem with water balance. However, marine bony fish like red cod, snapper and sole, have body fluids with a lower concentration of dissolved substances than seawater (they are hypotonic to seawater). This means that water tends to flow out of their bodies by osmosis. To make up this fluid loss they drink seawater and get rid of the excess salt by excreting it from the gills.
Marine Birds
Marine birds that eat marine fish take in large quantities of salt and some only have access to seawater for drinking. Bird’s kidneys are unable to produce very concentrated urine, so they have developed a salt gland. This excretes a concentrated salt solution into the nose to get rid of the excess salt.
2. Using the words/phrases in the list below fill in the blanks in the following statements.
| cortex | amino acids | renal | glucose | water reabsorption | large proteins |
| bowman’s capsule | diabetes mellitus | secreted | antidiuretic hormone (ADH) | blood cells |
| glomerulus | concentration of the urine | medulla | nephron |
a) Blood enters the kidney via the ……………………. artery.
b) When cut across the kidney is seen to consist of two regions, the outer………….. and the inner…………..
c) Another word for the kidney tubule is the………………………….
d) Filtration of the blood occurs in the…………………………
e) The filtered fluid (filtrate) enters the………………………..
f) The filtrate entering the e) above is similar to blood but does not contain……………… or………………..
g) As the fluid passes along the first coiled part of the kidney tubule……………… and……………….. are removed.
h) The main function of the loop of Henle is…………………………………………………….
i) Hydrogen and potassium ions are………………………… into the second coiled part of the tubule.
j) The main function of the collecting tube is…………………………………………..
k) The hormone……………………………….. is responsible for controlling water reabsorption in the collecting tube.
l) When the pancreas secretes inadequate amounts of the hormone insulin the condition known as…………………………. results. This is most easily diagnosed by testing for………………………….. in the urine.
Источник: infourok.ru
Источник: www.chem21.info
- ЧАСТЬ II. БИОЭНЕРГЕТИКА И МЕТАБОЛИЗМ
ГЛАВА 13. МЕТАБОЛИЗМ. ОБЩИЙ ОБЗОР
- 13.1. Живые организмы принимают участие в круговороте углерода и кислорода
- 13.2. В биосфере существует круговорот азота
- 13.3. Метаболические пути представляют собой последовательности реакций, катализируемых мультиферментными системами
- 13.4. Метаболизм включает катаболические и анаболические пути (процессы распада и процессы синтеза)
- 13.5. Катаболические пути сходятся — образуется лишь небольшое число конечных продуктов
- 13.6. Биосинтетические (анаболические) пути расходятся — образуется много разных продуктов
- 13.7. Соответствующие катаболические и анаболические пути различаются, и эти различия имеют важное значение.
- 13.8. Энергия перелается от катаболических реакций к анаболическим при помощи АТР
- 13.9. NADPH переносит энергию в форме восстановительной способности
- 13.10. Клеточный метаболизм — это экономичный, строго регулируемый лроцесс
- 13.11. Регуляция метаболических путей осуществляется на трех уровнях
- 13.12. Вторичный метаболизм
- 13.13. Метаболические пути могут быть идентифицированы в прямых опытах
- 13.14. Промежуточные стадии метаболизма можно выявлять с помощью мутантных организмов
- 13.15. Включение изотопной метки — весьма эффективный метод изучения метаболизма
- 13.16. Различные метаболические пути могут быть локализованы в разных участках клетки
- ЛИТЕРАТУРА
ГЛАВА 14. ATP — ЦИКЛ И БИОЭНЕРГЕТИКА КЛЕТКИ
- 14.1. Первый и второй законы термодинамики
- 14.2. Клеткам необходима свободная энергия
- 14.3. Изменение стандартной свободной энергии химической реакции можно вычислить
- 14.4. Химические реакции характеризуются определенной величиной
- 14.5. Величины различаются, и это различие имеет важное значение
- 14.6. Изменения стандартной свободной энергии химических реакций аддитивны
- 14.8. Химические свойства АТР хорошо известны
- 14.9. Характерное значение, стандартной свободной энергии АТР
- 14.10. Почему стандартная свободная энергия гидролиза АТР относительно велика?
- 14.11. АТР служит общим промежуточным продуктом в реакциях переноса фосфатных групп
- 14.12. При расщеплении глюкозы до лактата образуются два сверхвысокоэнергетических фосфорилированных соединения
- 14.13. В результате переноса фосфатной группы от АТР на какую-нибудь акцепторную молекулу этой молекуле сообщается энергия
- 14.14. АТР используется для обеспечения энергией мышечного сокращения
- 14.15. Креатинфосфат в мышцах выполняет роль резервуара высокоэнергетических фосфатных групп
- 14.17. АТР может расщепляться также до АМР и пирофосфата
- 14.18. Помимо АТР есть и другие высокоэнергетические нуклеотид-5-трифосфаты
- 14.19. Система АТР функционирует в стационарно-динамическом режиме
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 15. ГЛИКОЛИЗ-ЦЕНТРАЛЬНЫЙ ПУТЬ КАТАБОЛИЗМА ГЛЮКОЗЫ
- 15.1. Гликолиз является одним из центральных метаболических путей у большинства организмов
- 15.2. С гликолизом сопряжен синтез АТР
- 15.3. В продуктах гликолиза сохраняется еще много свободной энергии
- 15.4. Гликолиз включает две стадии
- 15.5. В ходе гликолиза образуются фосфорилированные промежуточные продукты
- 15.6. Первая сталия гликолиза завершается расщеплением углеродного скелета глюкозы
- 15.7. На второй стадии гликолиза запасается энергия
- 15.8. Пути, ведущие от гликогена и других углеводов, к центральному гликолитическому пути
- 15.9. В гликолиз могут вовлекаться и другие простые сахара
- 15.10. Дисахариды должны предварительно подвергнуться гидролизу то моносахаридов
- 15.11. Вовлечение остатков глюкозы в процесс гликолиза регулируется
- 15.12. Взаимопревращения фосфорилазы а и фосфорилазы b регулируются в конечном счете гормонами
- 15.13. Сама последовательность гликолитических реакций регулируется на двух главных этапах
- 15.14. Каким образом можно выявить регулируемые этапы гликолиза в интактных клетках?
- 15.15. Спиртовое брожение отличается от гликолиза только на последних этапах
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
- ГЛАВА 16. ЦИКЛ ЛИМОННОЙ КИСЛОТЫ
- 16.1. При окислении глюкозы до СО2 и Н2О высвобождается значительно больше энергии, чем при гликолизе
- 16.2. Пируват должен сначала окислиться до ацетил-СоА и СО2
- 16.3. Цикл лимонной кислоты — это не линейный, а замкнутый путь
- 16.4. Как родилась сама мысль о существовании цикла лимонной кислоты?
- 16.5. Цикл лимонной кислоты включает восемь стадий
- 16.6. Общая характеристика цикла
- 16.7. В чем смысл цикла лимонной кислоты?
- 16.8. Применение изотопных методов в изучении цикла лимонной кислоты
- 16.9. Превращение пирувата в ацетил-СоА регулируется
- 16.10. Цикл лимонной кислоты регулируется
- 16.11. Промежуточные продукты цикла лимонной кислоты используются также и в других метаболических реакциях, а убыль их постоянно восполняется
- 16.12. Глиоксилатный цикл — одна из модификаций цикла лимонной кислоты
- 16.13. Вторичные пути катаболизма глюкозы: пентозофосфатный путь
- 16.14. Вторичный путь, по которому происходит превращение глюкозы в глюкуроновую и аскорбиновую кислоты
- Краткое содержание главы
- ЛИТЕРАТУРА
- Вопросы и задачи
ГЛАВА 17. ПЕРЕНОС ЭЛЕКТРОНОВ, ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ И РЕГУЛЯЦИЯ СИНТЕЗА АТР
- 17.1. Перенос электронов от субстратов на кислород служит источником энергии АТР
- 17.2. Перенос электронов и окислительное фосфорилирование происходят во внутренней митохондриальной мембране
- 17.3. Реакции переноса электронов — это окислительновосстановительные реакции
- 17.4. Каждая сопряженная окислительно-восстановительная пара характеризуется определенным стандартным потенциалом
- 17.5. Перенос электронов сопровождается изменениями свободной энергии
- 17.6. Цепь переноса электронов включает большое число переносчиков
- 17.7. Пиридиновые нуклеотиды выполняют коллекторную функцию
- 17.8. NАDH-дегидpoгеназа принимает электроны от NADH
- 17.9. Убихинон представляет собой жирорастворимый хинон
- 17.10. Цитохромы — это гемопротеины, осуществляющие перенос электронов
- 17.11. Неполное восстановление кислорода ведет к повреждению клеток
- 17.12. Переносчики электронов действуют всегда в определенной последовательности
- 17.13. Энергия, выделяемая при переносе электронов, запасается в результате окислительного фосфорилирования
- 17.14. Фермент, катализирующий синтез АТР, был выделен и реконструирован
- 17.15. Каким образом окислительно-восстановительная энергия переноса электронов передается АТР-синтетазе?
- 17.16. Согласно хемиосмотической гипотезе энергия переноса электронов передается на синтез АТР через протонный градиент
- 17.17. Энергия переноса электронов используется и для других целей
- 17.18. В бактериальных клетках и в хлоропластах также имеются цепи переноса электронов, транспортирующие ионы
- 17.19. Внутренняя мембрана митохондрий содержит специфические транспортные системы
- 17.20. В окислении внемитохондриального NADH участвуют челночные системы
- 17.21. При полном окислении молекулы глюкозы образуется 38 молекул АТР
- 17.22. Образование АТР путем окислительного фосфорилирования регулируется в соответствии с энергетическими нуждами клетки
- 17.23. Энергетический заряд служит еще одним показателем энергетического состояния клеток
- 17.24. Регуляторные механизмы гликолиза, цикла лимонной кислоты и окислительного фосфорилирования взаимосвязаны
- 17.25. В клетках имеются и другие ферменты, использующие в качестве акцептора электронов кислород
- Краткое содержание главы
- Вопросы и задачи
ГЛАВА 18. ОКИСЛЕНИЕ ЖИРНЫХ КИСЛОТ В ТКАНЯХ ЖИВОТНЫХ
- 18.1. Жирные кислоты активируются и окисляются в митохондриях
- 18.2. Процесс поступления жирных кислот в митохондрии состоит из трех этапов
- 18.3. Окисление жирных кислот включает две сталии
- 18.4. Первая стадия окисления насыщенных жирных кислот состоит из четырех этапов
- 18.5. На первой стадии окисления жирных кислот образуются ацетил-СоА и АТР
- 18.7. Окисление ненасыщенных жирных кислот требует двух дополнительных ферментативных этапов
- 18.8. Окисление жирных кислот с нечетным числом атомов углерода
- 18.9. Гипоглицин (токсичное вещество, вырабатываемое некоторыми растениями) подавляет окисление жирных кислот
- 18.10. Образование кетоновых тел в печени и их окисление в других органах
- 18.11. Регуляция окисления жирных кислот и образования кетоновых тел
- Краткое содержание главы
- Вопросы и задачи
ГЛАВА 19. ОКИСЛИТЕЛЬНОЕ РАСЩЕПЛЕНИЕ АМИНОКИСЛОТ. ЦИКЛ МОЧЕВИНЫ
- 19.1. Перенос а-аминогрупп катализируется трансаминазами
- 19.2. Аммиак образуется из глутамата
- 19.3. Существует 20 различных путей для расщепления углеродных скелетов аминокислот
- 19.4. Десять аминокислот превращаются в результате расщепления в ацетил-СоА
- 19.5. Наследственные нарушения катаболизма фенилаланина
- 19.6. Пять аминокислот превращаются в а-кетоглутарат
- 19.7. Три аминокислоты превращаются в сукцинил-СоА
- 19.8. Из фенилаланина и тирозина образуется фумарат
- 19.9. Оксалоацетатный путь
- 19.10. Некоторые аминокислоты могут превращаться в глюкозу, а другие — в кетоновые тела
- 19.11. Аммиак для животных токсичен
- 19.12. Аммиак переносится в печень из многих периферических тканей в виде глутамина
- 19.13. Аммиак переносится из мышц в печень в виде аланина
- 19.14. Выведение аминиого азота из организма составляет еще одну сложную биохимическую проблему
- 19.15. В выделении аммиака участвует глутаминаза
- 19.16. Мочевина образуется в цикле мочевины
- 19.17. Цикл мочевины включает ряд сложных стадий
- 19.18. Энергетическая цена синтеза мочевины
- 19.19. Генетические дефекты, затрагивающие цикл мочевины, вызывают накопление аммиака в крови
- 19.20. У птиц, змей и ящериц из организма выводится мочевая кислота
- Краткое содержание главы
- Вопросы и задачи
- ГЛАВА 20. БИОСИНТЕЗ УГЛЕВОДОВ В ЖИВОТНЫХ ТКАНЯХ
- 20.1. Путь глюконеогенеза включает семь этапов, общих с процессом гликолиза
- 20.2. Обходный путь требуется для превращения пирувата в фосфоенолпируват
- 20.3. Второй обходный путь в глюконеогенезе — это превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат
- 20.4. Третий обходный путь — это путь, ведущий от глюкозо-6-фосфата к свободной глюкозе
- 20.5. Глюконеогенез требует значительных затрат энергии
- 20.6. Реципрокная регуляция глюконеогенеза и гликолиза
- 20.7. Промежуточные продукты цикла лимонной кислоты являются также предшественниками глюкозы
- 20.8. Большинство аминокислот относится к глюкогенным
- 20.9. Глюконеогенез происходит в период восстановления после мышечной работы
- 20.10. Особенно активный глюконеогенез свойствен жвачным животным
- 20.11. Алкоголь тормозит глюконеогенез
- 20.12. «Холостые» циклы в углеводном обмене
- 20.13. Путь биосинтеза гликогена отличается от пути его расщепления
- 20.14. Гликоген-синтаза и гликоген-фосфорилаза регулируются реципрокно
- 20.15. Существуют генетические болезни, при которых обмен гликогена нарушен
- 20.16. Синтез лактозы регулируется особым образом
- Краткое содержание главы
- Вопросы и задачи
ГЛАВА 21. БИОСИНТЕЗ ЛИПИДОВ
- 21.1. Путь биосинтеза жирных кислот отличается от пути их окисления
- 21.2. Малонил-СоА образуется из ацетил-СоА
- 21.3. Синтазная система, катализирующая образование жирных кислот, имеет семь активных центров
- 21.4. Сульфгидрильные группы синтазы жирных кислот вначале взаимодействуют с ацильными группами
- 21.5. Присоединение каждого двухуглеродного фрагмента происходит в четыре этапа
- 21.6. Пальмитиновая кислота служит предшественником других длннноцепочечных жирных кислот
- 21.7. Регуляция биосинтеза жирных кислот
- 21.8. Биосинтез триацилглицеролов и глицеролфосфатидов начинается с общих предшественников
- 21.9. Биосинтез триацилглицеролов регулируется гормонами
- 21.10. Триацилглицеролы — источник энергии для некоторых впадающих в спячку животных
- 21.11. Для биосинтеза фосфоглицеролов нужны группы, образующие головы молекул
- 21.12. Фосфатидилхолин образуется двумя разными путями
- 21.13. Полярные липиды встраиваются в клеточные мембраны
- 21.14. Генетические дефекты липидного обмена
- 21.15. Существуют многочисленные лизосомные болезни
- 21.16. Холестерол и другие стероиды также синтезируются из двухуглеродных предшественников
- 21.17. И зопентенилпирофосфат служит предшественником многих жирорастворимых биомолекул
- Краткое содержание главы
- Вопросы и задачи
ГЛАВА 22. БИОСИНТЕЗ АМИНОКИСЛОТ И НУКЛЕОТИДОВ
- 22.1. Некоторые аминокислоты должны поступать в организм с пищей
- 22.2. К глутамату, глутамину и пролину ведет общий биосинтетический путь
- 22.3. Аланин, аспартат и аспарагин тоже образуются из центральных метаболитов
- 22.4. Тирозин образуется из незаменимой аминокислоты фенилаланина
- 22.5. Цистеин образуется из двух других аминокислот — метионина и серина
- 22.6. Серии служиг предшественником глицина
- 22.7. Биосинтез незаменимых аминокислот
- 22.8. Биосинтез аминокислот регулируется аллостерическими механизмами
- 22.9. Биосинтез аминокислот регулируется также путем изменений концентрации ферментов
- 22.10. Глицин является предшественником порфиринов
- 22.11. При некоторых генетических заболеваниях накапливаются производные порфиринов
- 22.12. В результате распада гемогрупп образуются желчные пигменты
- 22.13. Пуриновые нуклеотиды синтезируются сложным путем
- 22.14. Биосинтез пуриновых нуклеотидов регулируется по типу обратной связи
- 22.15. Пиримидиновые нуклеотиды синтезируются из аспартата и рибозофосфата
- 22.16. Регуляция биосинтеза пиримидиновых нуклеотидов
- 22.17. Рибонуклеотиды служат предшественниками дезоксирибонуклеотидов
- 22.18. Распад пуринов приводит у человека к образованию мочевой кислоты
- 22.19. Реутилизация пуриновых оснований
- 22.20. Избыточное образование мочевой кислоты вызывает подагру
- 22.21. Круговорот азота
- 22.22. Способность фиксировать атмосферный азот присуща немногим организмам
- 22.23. Фиксация азота — сложный ферментативный процесс
- Краткое содержание главы
- Вопросы и задачи
ГЛАВА 23. ФОТОСИНТЕЗ
- 23.1. О том, как было выведено уравнение фотосинтеза
- 23.2. Фотосинтезирующие организмы чрезвычайно разнообразны
- 23.3. Доноры водорода у разных фотосинтезирующих организмов различны
- 23.4. Процесс фотосинтеза состоит из двух фаз — световой и темновой
- 23.5. Фотосинтез растений протекает в хлоропластах
- 23.6. Поглощение света переводит молекулы в возбужденное состояние
- 23.7. Хлорофиллы — это главные светопоглощающие пигменты
- 23.8. В тилакоидах содержатся также вспомогательные пигменты
- 23.9. В мембранах тилакоидов содержатся два типа фотохимических реакционных систем
- 23.10. Свет индуцирует в хлоропластах поток электронов
- 23.11. Улавливаемам световая энергия создает поток электронов, направленный «вверх»
- 23.12. Перенос электронов от H2O к NADP+ происходит в результате взаимодействия фотосистем I и II
- 23.13. Z-схема представляет фотосинтетический перенос электронов в виде энергетической диаграммы
- 23.14. В фотосинтетическом переносе электронов принимает участие ряд переносчиков
- 23.15. Фосфорилирование ADP сопряжено с фотосинтетическим переносом электронов
- 23.16. В хлоропластах возможен также циклический поток электронов и циклическое фотофосфорилирование
- 23.17. Фотосинтетическое фосфорилирование сходно с окислительным фосфорилированием
- 23.18. Общее уравнение фотосинтеза растений
- 23.19. Фотосинтетическое образование гексоз связано с реальным восстановлением двуокиси углерода
- 23.20. Двуокись углерода фиксируется в форме фосфоглицерата
- 23.21. Глюкоза образуется из СО2 в цикле Кальвина
- 23.22. Глюкоза служит предшественником типичных растительных углеводов — сахарозы, крахмала и целлюлозы
- 23.23. Регуляция темновых реакций
- 23.24. В тропических растениях используется С4-путь, или путь Хэтча-Слэка
- 23.25. С4-путь обеспечивает необходимую концентрацию СО2
- 23.26. Фотодыхание ограничивает продуктивность С3-растений
- 23.27. Фотодыхание серьезная проблема для земледелия умеренной зоны
- 23.28. Галофильные бактерии используют световую энергию для синтеза АТР
- 23.29. Фотосинтезирующие организмы служат моделями для конструирования солнечных батарей
- Краткое содержание главы
- Вопросы и задачи
- Приложение. Ответы
Источник: scask.ru