Лентические экосистемы
Лентические экосистемы в литоральной зоне содержат два типа продуцентов: укрепившиеся в дне цветковые растения и плавающие зеленые растения — водоросли, некоторые высшие (рдесты). Растения, укрепленные в дне, образуют три концентрические зоны: 1) зона надводной вегетации — фотосинтезирующая часть растений (камыши, рогозы и др.) находится над водой, а биогенные элементы извлекаются из донных осадков; 2) зона укрепленных в дне растений с плавающими по воде листьями (кувшинки) — у них та же роль, что и у растений первой зоны, но они могут затенять нижние толщи воды; 3) зона подводной вегетации — укорененные и прикрепленные растения, полностью находящиеся под водой и осуществляющие фотосинтез и минеральный обмен в водной среде (рдесты и прикрепленные водоросли — харовые).
Животные, консументы более разнообразны в литорали, чем в других зонах водоема. Перифитон представлен моллюсками, коловратками, мшанками, личинками насекомых и др. Многие животные нектона дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Зоопланктон представлен ракообразными, имеющими большое значение для питания рыб (дафнии и др.).
В сообществах лимнической зоны продуцентом является фитопланктон. В водоемах умеренного пояса плотность его популяции заметно изменяется по сезонам. Весной «цветение» связано с массовым развитием приспособленных к прохладной воде диатомитовых водорослей, летом — зеленых, осенью — азотфиксирующих сине-зеленых водорослей. Зоопланктон представлен растительноядными ракообразными и коловратками, все другие — хищники. Нектон лимнической зоны — это только рыбы.
Сообщества профундальной зоны существуют без света. Фауна и флора здесь — в зоне поверхностного раздела вода — ил, где накапливается органический материал, — представлена бактериями и грибами (редуценты), а также бен-тосными формами — личинками насекомых, моллюсками, кольчатыми червями (консументами).
Количество красных кольчатых червей возрастает с ростом загрязнения водоема сточными водами, т.е. по этому показателю можно судить о степени загрязнения водоема.
Действие на сообщества стоячих водоемов таких лимитирующих факторов, как содержание кислорода, температуры и освещенности, зависит от специфических особенностей этих водоемов — озер, прудов и искусственных водохранилищ.
Источник: ibrain.kz
Лентические экосистемы в литоральной зоне содержат два типа продуцентов: укрепившиеся в дне цветковые растения и плавающие зеленые растения ¾ водоросли, некоторые высшие (рдесты) (рис. 7.7). Растения, укрепленные в дне, образуют три концентрические зоны:
1) зона надводной вегетации ¾ фотосинтезирующая часть растений (камыши, рогозы и др.) находится над водой, а биогенные элементы извлекаются из донных осадков;
2) зона укрепленных в дне растений с плавающими по воде листьями (кувшинки) ¾ у них та же роль, что и у растений первой зоны, но они могут затенять нижние толщи воды;
3) зона подводной вегетации ¾ укорененные и прикрепленные растения полностью находящиеся под водой и осуществляющие фотосинтез и минеральный обмен в водной среде (рдесты и прикрепленные водоросли ¾ харовые).

Рис. 7.7. Некоторые продуценты лентических сообществ; прибрежные растения,
укореняющиеся в дне водоема (1-7), нитчатые водоросли (8-9)
и фитопланктон (10-20)
Животные, консументы, более разнообразны в литорали, чем в других зонах водоема. Перифитон представлен моллюсками, коловратками, мшанками, личинками насекомых и др. Многие животные нектона дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Зоопланктон представлен ракобразными, имеющими большое значение для питания рыб (дафнии и др.).
В сообществах лимнической зоны продуцентом является фитопланктон. В водоемах умеренного пояса плотность его популяции заметно изменяется по сезонам. Весной «цветение» связано с массовым развитием приспособленных к прохладной воде диатомитовых водорослей, летом ¾ зеленых, осенью ¾ азотфиксирующих синезеленых водорослей. Зоопланктон представлен растительноядными ракоообразными и коловратками, все другие ¾ хищники. Нектон лимнической зоны ¾ это только рыбы.
Сообщества профундальной зоны существуют без света. Фауна и флора здесь ¾ в зоне поверхностного раздела вода ¾ ил, где накапливается органический материал, ¾ представлена бактериями и грибами (редуценты), а также бентосными формами ¾ личинками насекомых, моллюсками, кольчатыми червями (консументами).
Количество красных кольчатых червей возрастает с ростом загрязнения водоема сточными водами, т. е. по этому показателю можно судить о степени загрязнения водоема.
Действие на сообщества стоячих водоемов таких лимитирующих факторов, как содержание кислорода, температуры и освещенности, зависит от специфических особенностей этих водоемов ¾ озер, прудов, искусственных водохранилищ.
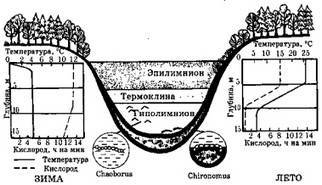
Озера¾ естественные пресноводные водоемы (см. рис. 7.9), образовались геологически сравнительно недавно ¾ за последние несколько десятков тысяч лет, и лишь возраст некоторых из них исчисляется миллионами лет, например, Байкала. Наличие у большинства озер профундальной зоны сказывается на температурном режиме водной толщи, на ее «перемешивании» и распределении кислорода в ней. Эти процессы сезонны, как и стратификация озера по температурному режиму (рис. 7.8).
Рис. 7.8. Температурная стратификация в озере умеренной зоны Северного полушария
(оз. Линсли, Коннектикут, США, по Ю. Одуму, 1975)
В озерах умеренного пояса в летнее время можно выделить в вертикальном разрезе три зоны: эпилимнион ¾ до глубины, где происходит конвекция (циркуляция) воды; термоклина ¾ это промежуточная зона, где вода не смешивается с водой верхней зоны; гиполимнион ¾ область холодной воды, где нет циркуляции.
Термоклина обычно расположена ниже границы проникновения света, и запасы кислорода, в отрезанном от его источников гиполимнионе, истощаются. Наступает летний период стагнации. Осенью, вследствие выравнивания температур, происходит общее перемешивание воды и обогащение гиполимниона кислородом. Зимой, когда температура воды под льдом становится ниже плюс 4 °С, что снижает ее плотность и снова приводит к стратификации озера и к зимней стагнации.
сной, после таяния льда, температура воды достигает плюс 4 °С, она тяжелеет и снова происходит весеннее перемешивание. Это классическая схема для водоемов Евразии и Северной Америки. В полярных областях и субтропиках перемешивание бывает только один раз в году: в первом случае ¾ летом, во втором ¾ зимой. В водоемах тропиков перемешивание воды идет постоянно, но медленно, а общее перемешивание происходит редко и нерегулярно.
Цветение фитопланктона обычно приурочено к перемешиванию, когда в эвфотической зоне появляются воды, обогащенные природными биогенными компонентами. С точки зрения продуктивности озера подразделяются на две группы: 1) олиготрофные (малокормные) и 2) эвтрофные (кормные). Продуктивность лентических экосистем зависит также от поступающих веществ с окружающей суши и от глубины озера (наиболее продуктивны мелкие озера).
Пруды обладают хорошо развитой литоралью и практическим отсутствием стратификации, образуются они в различных понижениях, часто временно пересыхают летом или в засушливые годы. Фауна прудов способна переживать сухие периоды в покоящемся состоянии или перебираться в другие водоемы (земноводные). Естественные пруды ¾ высокопродуктивны. В искусственных прудах, в основном, человек сам подкармливает рыб.
Водохранилища (см. рис. 7.9) создаются человеком при возведении гидроэнергетических и гидромелиоративных комплексов.
о уже не природная экосистема, а природно-техническая система. Распределение тепла и биогенов в ней зависит от типа плотины. Если вода сбрасывается придонная, то в этом случае водохранилище аккумулирует тепло и экспортирует биогенные вещества, если сброс идет поверх плотины, то экспортируется тепло и аккумулируются биогены. В первом случае спускается вода гиполимниона, во втором ¾ эпилимниона. Через глубоководные шлюзы в реку поступает и более соленая вода, а биогены вызывают эвтрофикацию участка реки.
Рис. 7.9. Пресноводные экосистемы
Лотические экосистемы¾реки (см. рис. 7.9) отличаются от стоячих водоемов тремя условиями:
1) течение ¾ важный лимитирующий и контролирующий фактор;
2) обмен между водой и сушей значительно более активен;
3) распределение кислорода более равномерно, так как практически отсутствует стратификация.
Скорость течения влияет на распределение рыб в реках ¾ они могут жить и под камнями, и в заводях, под перекатами, но это будут разные виды, адаптированные к конкретным условиям. Река ¾ открытая экосистема, в которую поступает с прилегающих пространств большое количество органического вещества.
Детритное питание ¾ основа трофических цепей лотических экосистем: более 60% энергии консументы получают от привнесенного материала. Зато кислорода в реках достаточно и содержание его в воде постоянно, что обусловило узкую толерантность организмов по отношению к кислороду.
Выделяют лотические сообщества перекатов и плесов. На перекатах поселяются организмы, способные прикрепиться к субстрату (нитчатые водоросли), или хорошие пловцы (форель). На участках плеса сообщества напоминают прудовые.
В больших реках прослеживается продольная зональность: в верховьях ¾ сообщества перекатов, в низовьях и дельте ¾ плесов, между ними местами могут возникать и те и другие. Продольная зональность подчеркивается изменениями видового состава рыб. К низовьям видовой состав обедняется, но увеличиваются размеры рыб.
Заболоченные пресноводные участки, обычно¾собственно болота (см. рис. 7.9) ¾ низинные и верховые. Низинные имеют, как правило, питание подземными водами, а верховые ¾ атмосферными осадками. Верховые могут встречаться в любом понижении или даже на склонах гор, низинные возникают вследствие зарастания озер и речных стариц. Они покрыты водными макрофитами, болотными растениями и кустарниками.
Болотные почвы и торфяники содержат много углерода (14-20%), сельскохозяйственная отработка которых приводит к выделению в атмосферу большого количества углекислого газа, что усугубляет «CO2-проблему».
Источник: studopedia.org
Вопрос 4. Классификация природных экосистем биосферы
Классификации природных систем биосферы базируются на ландшафтном подходе, так как экосистемы — неотъемлемая часть природных географических ландшафтов, образующих географическую (ландшафтную) оболочку Земли.
Ландшафт — природный географический комплекс, в котором все основные компоненты (рельеф, верхние горизонты литосферы, климат, воды, почвы, растительность, животный мир) находятся в сложном взаимодействии, образуя однородную по условиям развития единую систему. По происхождению выделяют два типа ландшафтов — природный и антропогенный.
Природный ландшафт формируется исключительно под влиянием природных факторов и не преобразован хозяйственной деятельностью человека. Выделяют следующие природные ландшафты:
— геохимический — обозначает участок, выделенный на основе единства состава и количества химических элементов и соединений;
— элементарный ландшафт — обозначает участок, сложенный определенными породами, находящимися на одном элементе рельефа, в равных условиях залегания грунто вых вод, с одинаковым характером растительных ассоциаций и одним типом почв;
— охраняемый ландшафт, на котором в установленном порядке регламентированы или запрещены все или отдельные виды хозяйственной деятельности.
Антропогенный ландшафт — это преобразованный хозяйственной деятельностью бывший природный ландшафт настолько, что изменена связь природных компонентов (занимают 50% суши).
Сюда относятся ландшафты:
— агрокультурный (сельскохозяйственный) — растительность в значительной степени заменена сельскохозяйственными и садовыми культурами;
— техногенный — структура обусловлена техногенной деятельностью человека, с использованием мощных технических средств (нарушение земель, загрязнение промышленными отходами). Сюда входит ландшафт индустриальный, образующийся под воздействием на него промышленных комплексов;
— городской (урбанистический) — спостройками, улицами и парками.
И.А. Шилов (2000) выделяет три главных направления в экологических механизмах антропогенного преобразования ландшафтов:
1) обеднение видового состава и упрощение связей в экосистемах, что снижает их устойчивость;
2) введение в исходный ландшафт мозаичности в результате деятельности человека повышает биоразнообразие и, следовательно, устойчивость системы;
3) антропогенные ландшафты сохраняют в чем-то черты природного и привлекательность для ряда организмов.
Границы ландшафтной оболочки Земли совпадают с границами биосферы. Главный источник энергии для ландшафтной оболочки, как и для биосферы — солнечная радиация. Но для биосферы она «движитель» биогеохимических циклов и главный компонент фотосинтеза. Однако на эти цели расходуется всего 2-3% энергии, остальная — на абиотическую среду, факторы которой определяют вместе с биотическими гомеостаз экосистем.
Неразрывное единство ландшафтной оболочки и биосферы свидетельствует о том, что живая природа существует в едином энергетическом поле всего ландшафта, поэтому природные экосистемы следует рассматривать как хорологические (пространственные) единицы биосферы, подобно тому, как ландшафты различного уровня следует рассматривать как пространственные единицы географической оболочки Земли.
Границы распространения биомов определяются ландшафтными компонентами материков, в названии — доминирующая растительность (кустарниковая, лесная). В водных экосистемах растительные организмы не доминируют, поэтому за основу взяты физические признаки среды обитания («стоячая», «текучая» вода, открытый океан и т.п.). Ю. Одум предложил следующую классификацию природных экосистем биосферы на принципах биомного подхода:
1. Наземные биомы. Тундра: арктическая и альпийская. Бореальные хвойные леса. Листопадный лес умеренной зоны. Степь умеренной зоны. Тропические степи и саванны. Чапарраль — районы с дождливой зимой и засушливым летом. Пустыня: травянистая и кустарниковая. Полувечнозеленый тропический лес: выраженный влажный и сухой сезоны. Вечнозеленый тропический дождевой лес.
1. Типы пресноводных экосистем. Лентические (стоячие воды): озера, пруды ит.д. Лотические (текучие воды): реки, ручьи и т. д. Заболоченные угодья: болота и болотистые леса.
2. Типы морских экосистем. Открытый океан (пелагическая): Воды континентального шельфа (прибрежные воды). Районы апвелинга (плодородные районы с продуктивным рыболовством).
Вопрос 5. Наземные экосистемы (биомы)
К наземным экосистемам относятся — экосистемы или биомы, сформированные в определенных климатических условиях. Рассмотрим основные наземные экоситемы.
Тундры характеризуются суровыми условиями (вечная мерзлота). Но длительный фотопериод и низкая испаряемость сильно снижают лимитирующее действие влажности и света. В тундре нет деревьев, есть мхи, лишайники, зимнезеленые кустарники, карликовые растения. Видовое разнообразие и продуктивность наземных экосистем тундры значительно ниже, чем подобных систем других районов.
Бореальные хвойные леса — тайга — распространены в северной части умеренной климатической зоны Северного полушария. Таежные растительные сообщества представлены хвойными деревьями — ель, пихта, лиственница, сосна. Из крупных животных в тайге — медведь, волк, лось, из мелких — птицы, белки, обилие насекомых. Леса весьма продуктивны, так как зеленый покров сохраняется круглый год. Они самые крупные в мире поставщики лесоматериалов. Восстановление леса после вырубки требует длительного сукцессионного цикла. Для ускорения процесса предлагается, например, вырубать его узкими просеками, и тогда обсеменение с незатронутых участков поможет быстро восстановить лес до первоначального вида.
Листопадные леса умеренной зоны (широколиственные леса) расположены южнее тайги, не имеют сплошного распространения. Климат здесь более мягкий. Доминируют бук и дуб. Крупные животные здесь те же, что и в тайге. Листопадные леса — это те районы суши, где человеческая цивилизация получила наибольшее развитие. Поэтому трудно сейчас найти широколиственные нетронутые леса — они заменены культурными сообществами.
Степи умеренной зоны — открытые пространства между лесами и пустынями в Евразии, в Северной Америке и Южной Америке, в Австралии и Новой Зеландии. Растительность преимущественно ксерофильного облика — дерновинные злаки. Животные ведут парный образ жизни (суслики) и стадный (копытные ). При умеренном выпасе копытные разбивают скопления мертвой листвы на поверхности почвы, что способствует дальнейшему росту трав. При перевыпасе — степная растительность деградирует, исчезают многолетние травы. Последствия — опустынивание. Почвы степей отличаются от лесных высоким содержанием гумуса — в 5-10 раз выше.
Пустыни травянистые и кустарниковые располагаются в фонических и субтропических регионах в Азии, Африке, Австралии, Северной и Южной Америках. Почвы пустынь — сероземы и светло-бурые. Критерий пустыни — осадков менее 250 мм/год, а испарение с водной поверхности — более 1000 мм/год. Растительность — весьма ксерофильные травы и полукустарники, суккуленты. Она разрежена, из-за чего травоядные животные существуют небольшими группами, парами и в одиночку. Стада образуют только копытные, способные быстро находить новые участки с кормами (антилопы и др.). Животные пустыни по-разному адаптированы к нехватке воды: обладают особыми покровами, могут образовывать метаболическую воду и т.д. Земледелие в пустынях невозможно без орошения. Однако антропогенное воздействие приводит к сокращению площади всех остальных регионов, кроме пустынь — пустыни увеличиваются, особенно в Африке.
Тропические степи и саванны — теплые области в Центральной и Восточной Африке, в Южной Америке и Австралии. Сезонность связана с распределением осадков — сезоны влажные и сухие. Цветут деревья в конце сухого сезона, с началом дождей распускается листва, в засуху листья опадают. В саванне сочетаются мощный травяной покров с редкими деревьями (баобабы, пальмы), она не имеет себе равных по разнообразию и численности копытных (антилопы, зебры и др.) и хищников (львы, гепарды и др.). Разнообразны птицы (грифы, страусы и др.), множество змей и ящериц, насекомых. Среди последних много переносчиков возбудителей болезней.
Вечнозеленые тропические дождевые леса расположены вдоль экватора, где 2000-2500 мм/год осадков при равномерном распределении по месяцам: в бассейнах рек Амазонки и Ориноко; Конго; Нигера и Замбези; на острове Мадагаскар; в зонах Индо-Малайской и островов Борнео — Новая Гвинея. Годовой ход температур здесь ровный. Травянистая растительность и кустарники практически отсутствуют, но большое количество лиан и эпифитов. Видовое разнообразие растений очень велико — на нескольких гектарах столько видов, сколько нет во флоре всей Европы.
Влажные тропические леса — это достаточно древние климаксные экосистемы, в которых питательные вещества теряются и немедленно поступают в биологический круговорот, осуществляемый мутуалистическими организмами и неглубо кими, воздушными, корнями деревьев. Поэтому на скудных почвах так пышно растут леса. Разнообразие видов животных в этих лесах можно проиллюстрировать такими цифрами: на 15 км2 леса в Панаме — 20 тыс. видов насекомых, а на такой же территории Европы — несколько сотен. Из крупных животных тропических лесов наиболее известны обезьяны, ягуары, муравьеды, пумы, буйволы, индийские слоны, попугаи и др. Влажные тропические леса обладают самой большой биомассой и самой высокой продуктивностью из биоценозов суши. У них самая высокая скорость эволюции и видообразования. Многие виды вошли в состав северных сообществ и очень важно сохранить эти леса, как «ресурс генов». Тем не менее они продолжают уничтожаться.
Составляя весьма малую часть от всех экосистем биосферы (например, в любой отрезок времени в атмосфере воды содержится в 10 раз больше, чем во всех реках мира), пресноводные экосистемы для человека имеют непреходящее значение, вследствие того, что они практически единственный источник для бытовых и промышленных нужд. Пресные воды на поверхности континентов образуют реки, озера, болота, заполняют искусственные пруды и крупные водохранилища. Значит, пресные воды могут находиться в текучем, в относительно неподвижном стоячем и промежуточном состояниях.
Лимитирующие факторы водной среды — температура, прозрачность, течение, соленость и др. Многие водные животные — стенотермны — для них опасно даже небольшое тепловое загрязнение. Прозрачность — глубина зоны, в которой возможен фотосинтез при проникновении солнечного света. Течение влияет на распространение организмов и содержание газов и солей. Важнейшим лимитирующим фактором является концентрация кислорода. Лимитирующие из биогенных солей — нитраты и фосфаты. Разница в концентрации солей у гидробионта с окружающей водной средой может повышать или понижать давление жидкости в теле рыбы, и то и другое ведет к ее гибели. Поэтому пресноводные рыбы не могут жить в море, а морские — в пресноводном водоеме. Но есть рыбы со специальным механизмом осмотической регуляции, способные жить в обеих средах (лосось идр.).
Пищевые цепив водоемах хорошо развиты и представлены организмами всех трофических уровней. Водные организмы, с экологических позиций, классифицируются по местообитанию в водоеме: бентос — организмы, живущие на дне; перифитон- прикрепленные к стеблям водных растений или к другим выступам над дном водоема. Планктон — организмы плавают в поверхностном слое воды; нектон— свободно перемещающиеся в воде организмы.
Особое значение имеет распределение организмов по трем зонам водоема. Литоральная зона— толща воды, где солнечный свет доходит до дна. Лимническая зона— толща воды до глубины, куда проникает всего один процент от солнечного света и где затухает фотосинтез. Эвфотической зонойназывают всю освещенную толщу воды в литоральной и лимнической зонах. Профундальная зона— дно итолща воды, куда не проникает солнечный свет. В проточных водоемах последние три зоны не выражены, хотя их элементы встречаются — перекаты и плесы.
Лентические экосистемы.Озера — естественные пресноводные водоемы. Наличие убольшинства озер профундальной зоны сказывается на температурном режиме водной толщи, на ее «перемешивании» и распределении кислорода в ней. Эти процессы сезонны, как истратификация озера по температурному режиму. Известно, что наибольшую плотность, равную 1 г/см3, вода имеет при 4°С, а выше и ниже этой отметки плотность ее понижается. В весенний и осенний периоды, когда вода на поверхности водоема имеет температуру +4°С, происходит ее перемешивание, а в летний и зимний периоды — наступают стратификация водоема и периоды стагнации. Цветение фитопланктона приурочено к перемешиванию, когда вфотической (освещенной) зоне появляются воды, обогащенные биогенными компонентами. С точки зрения продуктивности озера подразделяются на две группы: 1) олиготрофные (малокормные) и 2) эвтрофные (кормные).
Водохранилища создаются человеком при возведении гидроэнергетических и гидромелиоративных комплексов. Это уже не природная экосистема, а природно-техническая система. Распределение тепла и биогенов в ней зависит от типа плотины. Если вода сбрасывается придонная, то в этом случае водохранилище аккумулирует тепло и экспортирует биогенные вещества. Если сброс идет поверх плотины, то экспортируется тепло и аккумулируются биогены. Через глубоководные шлюзы в реку поступает и более соленая вода, а биогены вызывают эвтрофикацию участка реки.
Лотические экосистемы — реки — отличаются от стоячих водоемов тремя основными условиями:
1) течение — важный лимитирующий и контролирующий фактор;
2) обмен между водой и сушей значительно более активен;
3)распределение кислорода более равномерно, так как практически отсутствует стратификация. В больших реках наблюдается продольная зональность: в верховьях — сообщества перекатов, в низовьях — плесов, между ними местами возникают и те и другие. К низовьям видовой состав рыб обедняется, но увеличиваются их размеры.
Заболоченные угодья и болота. Болота по своему происхождению бывают верховые и низинные. Низинные питаются подземными водами и образуются при зарастании озер, речных стариц и других водоемов, а верховые — атмосферными осадками и могут возникнуть в любом понижении, даже на склонах гор. Болота покрыты водными макрофитами, болотными растениями и кустарниками. Болотные почвы и торфяники содержат много углерода (14-20%). Их сельскохозяйственная обработка приводит к выделению в атмосферу большого количества углекислого газа.
Источник: studopedia.ru
— лентические экосистемы – озёра и пруды – стоячие воды;
— лотические экосистемы – родники, ручьи, реки – текучие воды;
— заболоченные участки, с колеблющимся уровнем по сезонам и годам – марши и болота.
Лимитирующие факторы водной среды — температура, прозрачность, течение, соленость и др. Многие водные животные — стенотермны — для них опасно даже небольшое тепловое загрязнение. Для жизни в водоёмах очень важна прозрачность воды. Прозрачность — глубина зоны, в которой возможен фотосинтез при проникновении солнечного света. Течение влияет на распространение организмов и содержание газов и солей. Важнейшим лимитирующим фактором является концентрация кислорода. Лимитирующие из биогенных солей — нитраты и фосфаты. Разница в концентрации солей у гидробионта с окружающей водной средой может повышать или понижать давление жидкости в теле рыбы, и то и другое ведет к ее гибели. Поэтому пресноводные рыбы не могут жить в море, а морские — в пресноводном водоеме. Но есть рыбы со специальным механизмом осмотической регуляции, способные жить в обеих средах (лосось и др.), потому что у них есть специальные механизмы осмотической регуляции.
Пищевые цепи в водоемах хорошо развиты и представлены организмами всех трофических уровней. Продуценты представлены автотрофами, консументы – полным набором от растительноядных и хищников до паразитов, редуценты – отличаются разнообразием, так как связаны с природой субстрата. Водные организмы, с экологических позиций, классифицируются по местообитанию в водоеме: бентос — организмы, живущие на дне; перифитон – животные и растения, прикрепленные к стеблям водных растений или к другим выступам над дном водоема; планктон — организмы плавают в поверхностном слое воды; нектон — свободно перемещающиеся в воде организмы.
Особое значение имеет распределение организмов по трем зонам водоема. Литоральная зона — толща воды, где солнечный свет доходит до дна. Лимническая зона — толща воды до глубины, куда проникает всего один процент от солнечного света и где затухает фотосинтез. Эвфотической зоной называют всю освещенную толщу воды в литоральной и лимнической зонах. Профундальная зона — дно и толща воды, куда не проникает солнечный свет. В проточных водоемах последние три зоны не выражены, хотя их элементы встречаются — перекаты и плесы.
Лентические экосистемы. Озера — естественные пресноводные водоемы. Наличие у большинства озер профундальной зоны сказывается на температурном режиме водной толщи, на ее «перемешивании» и распределении кислорода в ней. Эти процессы сезонны, как и стратификация озера по температурному режиму. Известно, что наибольшую плотность, равную 1 г/см3, вода имеет при 4° С, а выше и ниже этой отметки плотность ее понижается. В весенний и осенний периоды, когда вода на поверхности водоема имеет температуру + 4° С, происходит ее перемешивание, а в летний и зимний периоды — наступают стратификация водоема и периоды стагнации. Цветение фитопланктона приурочено к перемешиванию, когда в фотической (освещенной) зоне появляются воды, обогащенные биогенными компонентами. С точки зрения продуктивности озера подразделяются на две группы:
1) олиготрофные (малокормные);
2) эвтрофные (кормные).
Водохранилища создаются человеком при возведении гидроэнергетических и гидромелиоративных комплексов. Это уже не природная экосистема, а природно — техническая система. Распределение тепла и биогенов в ней зависит от типа плотины. Если вода сбрасывается придонная, то в этом случае водохранилище аккумулирует тепло и экспортирует биогенные вещества. Если сброс идет поверх плотины, то экспортируется тепло и аккумулируются биогены. Через глубоководные шлюзы в реку поступает и более соленая вода, а биогены вызывают эвтрофикацию участка реки.
Лотические экосистемы — реки — отличаются от стоячих водоемов тремя основными условиями:
1) течение — важный лимитирующий и контролирующий фактор;
2) обмен между водой и сушей значительно более активен;
3) распределение кислорода более равномерно, так как практически отсутствует стратификация. В больших реках наблюдается продольная зональность: в верховьях — сообщества перекатов, в низовьях — плесов, между ними местами возникают и те и другие. К низовьям видовой состав рыб обедняется, но увеличиваются их размеры.
Заболоченные угодья и болота. Болота по своему происхождению бывают верховые и низинные. Низинные питаются подземными водами и образуются при зарастании озер, речных стариц и других водоемов, а верховые — атмосферными осадками и могут возникнуть в любом понижении, даже на склонах гор. Болота покрыты водными макрофитами, болотными растениями и кустарниками. Болотные почвы и торфяники содержат много углерода (14 — 20%). Их сельскохозяйственная обработка приводит к выделению в атмосферу большого количества углекислого газа.
4. Морские экосистемы
Морская среда непрерывна и занимает более 70% поверхности планеты. Глубина океана огромна (до 11 км), но жизнь есть во всех его уголках и наиболее богата вблизи суши. Барьерами для передвижения животных являются температура, соленость, глубина, тем не менее, в океане отсутствуют абиотические зоны. Из — за постоянно действующих ветров — пассатов — в океанах и морях происходит постоянная циркуляция воды за счет мощных течений, что исключает дефицит кислорода в глубинах океана. Наиболее продуктивны в Мировом океане области апвеллинга. Апвеллинг — процесс подъема холодных вод с глубины океана там, где ветры постоянно перемещают воду прочь от крутого материкового склона, взамен которой поднимается из глубины вода, обогащенная биогенами. Соленость в них колеблется по сезонам года, поэтому здесь живут эвригалинные организмы, в отличие от таковых в открытом океане (средняя соленость 35 г/л), которые являются стеногалинными. Для морских водоёмов характерна устойчивая щелочная среда: рН = 8,2.
Биогенные элементы — важный лимитирующий фактор в морской среде, где их содержится несколько частей на миллиард частей воды. Но эти элементы быстро перехватываются организмами, попадая в их трофические цепи, практически не достигнув гетеротрофной зоны (биологический круговорот). Значит, низкая концентрация биогенов еще не говорит об их всеобщем дефиците.
Главным фактором, дифференцирующим морскую биоту, является глубина моря: материковый шельф резко сменяется материковым склоном, плавно переходящим в материковое подножие, которое опускается ниже к ровному ложу океана — абиссальной равнине. Этим морфологическим частям океана примерно соответствуют следующие зоны: неритическая — шельфу (с литоралью — приливно — отливной зоной), батиальная — материковому склону и его подножию; абиссальная — область океанических глубин от 2000 до 5000 м. Область открытого океана за пределами шельфа называют океанической. Все население океана, так же как и в пресноводных экосистемах, делится на планктон, нектон, бентос. Планктон и нектон, все, что живет в открытых водах, образует так называемую пелагическую зону.
Самая верхняя часть океана, куда проникает свет и где создается первичная продукция, называется эвфотической. Ее мощность в открытом океане доходит до 200 м, а в прибрежной части — не более 30 м. По сравнению с километровыми глубинами, эта зона достаточно тонкая и отделяется компенсационной зоной от значительно большой водной толщи, вплоть до самого дна — афотической зоны.
Биотические сообщества каждой из указанных зон, кроме эвфотической, разделяются на бентосные и пелагические. В них к первичным консументам относятся зоопланктон, насекомых в море экологически заменяют ракообразные. Подавляющее число крупных животных — хищники. Для моря характерна очень важная группа животных, которую называют сессилъными (прикрепленными). Их нет в пресноводных системах (морские лилии). Все животные бентоса в своем жизненном цикле проходят пелагическую стадию в виде личинок.
Область континентального шельфа, ограниченная глубиной моря до 200 м. Самая богатая фауной в океане. Очень богат кормом планктон за счет личинок бентосной фауны. Области апвелинга расположены вдоль западных пустынных берегов континентов. Они богаты рыбой и птицами, живущими на островах. Но при изменении направления ветра приходит спад «цветения» планктона и наступает массовая гибель рыб вследствие развития бескислородных условий (эвтрофикация). Лиманы — это полузамкнутые прибрежные водоемы. Они представляют собой экотоны между пресноводными и морскими экосистемами. Лиманы высокопродуктивны, являются ловушками биогенных веществ. Они служат для откорма молоди. Попадая в сферу хозяйственной деятельности человека, они могут потерять свою продуктивность вследствие загрязнения водной среды. Океанические области — эвфотическая зона открытого океана, бедны биогенными элементами. Это воды «пустыни» по сравнению с прибрежными. Зоны Арктики и Антарктики намного продуктивнее, так как плотность планктона растет при переходе к холодным морям и фауна рыб и китов здесь значительно богаче.
Экосистемы глубоководных рифтовых зон океана находятся на глубине около 3000 м и более, в сплошной темноте, где невозможен фотосинтез, преобладает сероводородное загрязнение, есть выходы горячих подземных вод, высокие концентрации ядовитых металлов, и тем не менее здесь существует жизнь.
5. Целостность биосферы как глобальной экосистемы
Закон целостности биосферы можно сформулировать так: биогенный ток атомов между компонентами биосферы связывает их в единую материальную систему, в которой изменение даже одного звена влечет за собой сопряженное изменение всех остальных. Следовательно, целостность биосферы обусловлена непрерывным обменом вещества и энергии между ее составными частями.
Характеристика природных экосистем
Примером действия закона целостности являются процессы, происходящие в экосистемах пустыни Атакама и прилегающей к ней части океана, которая находится на западном побережье Южной Америки, и пустынность ее обусловлена холодным Перуанским течением (количество осадков 10 — 50 мм/г). Холодные же океанские воды (зона апвеллинга) богаты фито — и зоопланктоном и рыбой. Но раз в 8 — 12 лет от экватора начинает распространяться теплое течение Эль — Ниньо. Приход этих бедных кислородом малопродуктивных вод приводит к катастрофическому изменению экосистемы: рыба — анчоусы, которых здесь вылавливают до 12 млн. т/год, практически исчезает, (улов падает до 1,8 млн. т), морские птицы, питающиеся рыбой, гибнут или улетают. В этот же период над пустыней Атакама разражаются тропические ливни, вызывающие мощные наводнения, появляются растения — эфемеры и масса насекомых. Пустыня «цветет». Такое состояние может продолжаться до пяти-шести месяцев, но затем снова теплое течение Эль — Ниньо отодвигается к экватору, в район Галапагосских островов, а холодное Перуанское — занимает свое обычное место. И все природные процессы развиваются в обратном направлении.
Изучение этого явления в течение многих десятилетий показало, что оно влияет на значительно большую часть биосферы — выпадение осадков в Атакаме приводит к засухе, например, в Судане и Эфиопии, из чего следует, что при решении практических задач рационального
Источник: myunivercity.ru
Рис. 7.7. Некоторые продуценты лентических сообществ: прибрежные растения, укореняющиеся в дне водоема (1—7), нитчатые водоросли (8—9) и фитопланктон (10—20) (по Ю.Одуму, 1975)
нижние толщи воды; 3) зона подводной вегетации — укорененные и прикрепленные растения, полностью находящиеся под водой и осуществляющие фотосинтез и минеральный обмен в водной среде (рдесты и прикрепленные водоросли — харовые).
Животные, консументы более разнообразны в литорали, чем в других зонах водоема. Перифитон представлен моллюсками, коловратками, мшанками, личинками насекомых и др. Многие животные нектона дышат кислородом атмосферного воздуха (лягушки, саламандры, черепахи и др.). Рыбы большую часть жизни проводят в литорали и здесь же размножаются. Зоопланктон представлен ракообразными, имеющими большое значение для питания рыб (дафнии и др.).
В сообществах лимнической зоны продуцентом является фитопланктон. В водоемах умеренного пояса плотность его популяции заметно изменяется по сезонам. Весной «цветение» связано с массовым развитием приспособленных к прохладной воде диатомитовых водорослей, летом — зеленых, осенью — азотфиксирующих сине-зеленых водорослей. Зоопланктон представлен растительноядными ракообразными и коловратками, все другие — хищники. Нектон лимнической зоны — это только рыбы.
Сообщества профундальной зоны существуют без света. Фауна и флора здесь — в зоне поверхностного раздела вода — ил, где накапливается органический материал, — представлена бактериями и грибами (редуценты), а также бентосными формами — личинками насекомых, моллюсками, кольчатыми червями (консументами).
Количество красных кольчатых червей возрастает с ростом загрязнения водоема сточными водами, т.е. по этому показателю можно судить о степени загрязнения водоема.
Действие на сообщества стоячих водоемов таких лимитирующих факторов, как содержание кислорода, температуры и освещенности, зависит от специфических особенностей этих водоемов — озер, прудов и искусственных водохранилищ.
Озера — естественные пресноводные водоемы, образовались геологически сравнительно недавно — за последние
несколько десятков тысяч лет, и лишь возраст некоторых из них исчисляется миллионами лет, например Байкала. Наличие у большинства озер профундальной зоны сказывается на температурном режиме водной толщи, на ее «перемешивании» и распределении кислорода в ней. Эти процессы сезон- ны, как и стратификация озера по температурному режиму (рис. 7.8).
Рис. 7.8. Температурная стратификация в озере умеренной зоны Северного полушария (оз. Линсли, Коннектикут, США; по Ю. Одуму, 1975)
В озерах умеренного пояса в летнее время можно выделить в вертикальном разрезе три зоны: эпилимнион — до глубины, где происходит конвекция (циркуляция) воды; термоклина — это промежуточная зона, где вода не смешивается с водой верхней зоны; гиполимнион — область холодной воды, где нет циркуляции.
Термоклина обычно расположена ниже границы проникновения света, и запасы кислорода, в отрезанном от его источников гиполимнионе, истощаются. Наступает летний период стагнации. Осенью, вследствие выравнивания температур, происходит общее перемешивание воды и обогащение гиполимниона кислородом. Зимой, когда температура воды подо льдом становится ниже 4 °С, что снижает ее плотность и снова приводит к стратификации озера и к зимней стагнации. Весной, после таяния льда, температура воды достигает 4 °С, она тяжелеет и снова происходит весеннее перемешивание. Это классическая схема для водоемов Евразии и Северной Америки. В полярных областях и субтропиках общее перемешивание воды в водоемах бывает только один раз в году: в первом случае — летом, во втором — зимой. В водоемах тропиков перемешивание воды идет постоянно, но медленно, а общее ее перемешивание происходит редко и нерегулярно.
Цветение фитопланктона обычно приурочено к перемешиванию, когда афотической (освещенной) зоне появляются воды, обогащенные природными биогенными компонентами. С точки зрения продуктивности озера подразделяются на две группы: 1) олиготрофные (малокормные) и 2) эвтрофные (кормные). Продуктивность лентических экосистем зависит также от поступающих веществ с окружающей суши и от глубины озера (наиболее продуктивны мелкие озера).
Пруды обладают хорошо развитой литоралью, и стратификация практически отсутствует; образуются они в различных понижениях, часто временно пересыхают летом или в засушливые годы. Фауна прудов способна переживать сухие периоды в покоящемся состоянии или перебираться в другие водоемы (земноводные). Естественные пруды высокопродуктивны. В искусственных прудах, в основном, человек сам подкармливает рыб.
Водохранилища создаются человеком при возведении гидроэнергетических и гидромелиоративных комплексов. Это уже не природная экосистема, а природно-техническая система. Распределение тепла и биогенов в ней зависит от типа плотины. Если вода сбрасывается придонная, то в этом случае водохранилище аккумулирует теплой экспортирует биогенные вещества, если сброс идет поверх плотины, то экспортируется тепло и аккумулируются биогены. В первом случае спускается вода гиполимниона, во втором — эпи- лимниона. Через глубоководные шлюзы в реку поступает и более соленая вода, а биогены вызывают эвтрофикацию участка реки.
Логические экосистемы — реки — отличаются от стоячих водоемов тремя основными условиями: 1) течения — важный лимитирующий и контролирующий фактор; 2) обмен между водой и сушей значительно более активен; 3) распределение кислорода более равномерно, так как практически отсутствует стратификация.
Скорость течения влияет на распределение рыб в реках — они могут жить и под камнями, и в заводях, под перекатами, но это будут разные виды, адаптированные к конкретным условиям. Река — открытая экосистема, в которую поступает с прилегающих пространств большое количество органического вещества.
Детритное питание — основа трофических цепей лоти- ческих экосистем: более 60% энергии консументы получают от привнесенного материала. Зато кислорода в реках достаточно и содержание его в воде постоянно, что обусловило узкую толерантность организмов по отношению к кислороду-
Выделяют лотические сообщества перекатов и плесов. На перекатах поселяются организмы, способные прикрепиться к субстрату (нитчатые водоросли), или хорошие пловцы (форель). На участках плеса сообщества напоминают прудовые.
В больших реках прослеживается продольная зональность: в верховьях — сообщества перекатов, в низовьях и дельте — плесов, между ними местами могут возникать и те и другие. Продольная зональность подчеркивается изменениями видового состава рыб. К низовьям видовой состав обедняется, но увеличиваются размеры рыб.
Заболоченные пресноводные участки, обычно собственно болота, — низинные и верховые. Низинные имеют, как правило, питание подземными водами; а верховые — атмосферными осадками. Верховые могут встречаться в любом понижении или даже на склонах гор, низинные возникают вследствие зарастания озер и речных стариц. Они покрыты водными макрофитами, болотными растениями и кустарниками.
Болотные почвы и торфяники содержат много углерода (14—20%), сельскохозяйственная отработка которых приводит к выделению в атмосферу большого количества углекислого газа, что усугубляет С02-проблему.
Источник: bookucheba.com