Роль аппарата гольджи
Строение комплекса Гольджи
Комплекс Гольджи (КГ), или внутренний сетчатый аппарат, — это особенная часть метаболической системы цитоплазмы, участвующая в процессе выделения и формирования мембранных структур клетки.
КГ видно в оптический микроскоп как сетку или изогнутые палочкообразные тельца, лежащие вокруг ядра.
Под электронным микроскопом выявлено, что эта органелла представлена тремя видами образований:
- многоярусной системой сплющенных дискообразных цистерн (диктиосомы), плотно расположенных пучками на расстоянии 14-25 нм с внутренним пространством 5-20 нм (чаще всего по 5-6 мешочков в комплексе);
- системой трубочек диаметром 20-50 нм;
-
системой пузырьков (везикул) — размеры мелких пузырьков – 20-30 нм, больших – до 2000 нм.

Все компоненты аппарата Гольджи образованы гладкими мембранами.
АГ встречается во всех клетках растений и животных.
Внутреннее
.
ферменты.
Аппарат Гольджи имеет две зоны:
- зону формирования, куда с помощью везикул поступает материал, который синтезируется в эндоплазматической сети;
- зону созревания, где формируется секрет и секреторные мешочки.
1069;тот секрет накопляется на терминальных участках АГ, откуда отпочковываются секреторные везикулы. Как правило, такие везикулы переносят секрет за пределы клетки. -
Локализация КГ
В ап&#.
080;мает место между ядром и апикальным полюсом.
Комплекс мешочков Гольджи имеет две поверхности:
формировательную (незрелую или регенераторную) цис-поверхность (от лат.
1057;is – с этой стороны); функциональную (зрелую) – транс-поверхность (от лат. Trans – через, за).
Столбик Гольджи своей выпуклой формировательной поверхностью обращён в сторону ядра, прилегает к гранулярной эндоплазматической сети и содержит мелкие круглые пузырьки, названные промежуточными.
1047;релая вогнутая поверхность столбика мешочков обращена к вершине (апикальному полюсу) клетки и оканчивается большими пузырьками.

Образование комплекса Гольджи
Мембраны КГ синтезируются гранулярной эндоплазматической сетью, которая прилегает к комплексу. Соседние с ним участки ЭПС теряют рибосомы, от них отпочковываются мелкие, так называемые, транспортные, или промежуточные везикулы. Они перемещаются к формировательной поверхности столбика Гольджи и сливаются с первым её мешочком. На противоположной (зрелой) поверхности комплекса Гольджи находится мешочек неправильной формы. Его расширение – просекреторные гранулы (конденсирующие вакуоли) – непрерывно отпочковываюся и превращаются в пузырьки, заполненные секретом – секреторные гранулы. Таким образом, в меру использования мембран зрелой поверхности комплекса на секреторные везикулы, мешочки формировательной поверхности пополняются за счёт эндоплазматической сетки.
Функции комплекса Гольджи
Основная функция аппарата Гольджи – выведение синтезированных клеткой веществ. Эти вещества транспортируются по клетках эндоплазматической сети и накопляются в пузырьках сетчатого аппарата. Потом они или выводятся во внешнюю среду или же клетка использует их в процессе жизнедеятельности.
В комплексе так же концентрируются некоторые вещества (например, красители), которые поступают в клетку извне и должны быть выведены из неё.
В растительных клетках комплекс содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения целлюлозной оболочки клетки.
Кроме того, КГ синтезирует те химические вещества, которые образуют клеточную мембрану.
В общем, аппарат Гольджи выполняет такие функции:
- накопление и модификация макромолекул, которые синтезировались в эндоплазматической сети;
- образование сложных секретов и секреторных везикул путём конденсации секреторного продукта;
- синтез и модификация углеводов и гликопротеидов (образование гликокаликса, слизи);
- модификация белков – добавление к полипептиду различных химических образований (фосфатных – фосфориллирование, карбоксильных – карбоксилирование), формирование сложных белков (липопротеидов, гликопротеидов, мукопротеидов) и расщепление полипептидов;
- имеет важное значение для формирования, обновления цитоплазматической мембраны и других мембранных образований благодаря образованию мембранных везикул, которые в дальнейшем сливаются с клеточной мембраной;
- образование лизосом и специфической зернистости в лейкоцитах;
- образование пероксисом.
Белковое и, частично, углеводное содержимое КГ поступает с гранулярной эндоплазматической сетки, где оно синтезируется. Основная часть углеводного компонента образуется в мешочках комплекса с участием ферментов гликозилтрансфераз, которые находятся в мембранах мешочков.
В комплексе Гольджи окончательно формируются клеточные секреты, содержащие гликопротеиды и гликозаминогликаны. В КГ созревают секреторные гранулы, которые переходят в пузырьки, и перемещение этих пузырьков в направлении плазмалеммы Окончательный этап секреции – это выталкивание сформированных (зрелых) везикул за пределы клетки. Выведение секреторных включений из клетки осуществляется путём вмонтирования мембран пузырька в плазмалемму и выделение секреторных продуктов за пределы клетки. В процессе перемещения секреторных пузырьков к апикальному полюсу клетки мембраны их утолщаются из начальных 5-7 нм, достигая толщины плазмалеммы 7-10 нм.
Источник: spravochnick.ru
Эндоплазматический ретикулум, плазматическая мембрана и аппарат Гольджи составляют единую мембранную систему клетки, в пределах которой происходят процессы обмена белками и липидами с помощью направленного и регулируемого внутриклеточного мембранного транспорта.
Каждая из мембранных органелл характеризуется уникальным составом белков и липидов.
Строение АГ
АГ состоит из группы плоских мембранный мешков — цистерны, собранные в стопки — диктиосомы (~5-10 цистерн, у низших эукариот >30). Число диктиосом в разных клетках от 1 до ~500.
Отдельные цистерны диктиосомы переменной толщины — в центре ее мембраны сближены — просвет 25 нм, на переферии образуются расширения — ампулы ширина которых не постоянна. От ампул отшнуровываются ~50нм-1мкм пузырьки связанные с цистернами сетью трубочек.
У многоклеточных организмов АГ состоит из стопок цистерн связанных между собой в единую мембранную систему. АГ представляет собой полусферу, основание которой обращено к ядру. АГ дрожжей представлен изолированными единичными цистернами, окруженными мелкими пузырьками, тубулярной сетью, секреторными везикулами и гранулами. У мутантов дрожжей Sec7 и Sec14 наблюдается структура, напоминающая стопку цистерн клеток млекопитающих.
Для АГ характерна полярность его структур. Каждая стопка имеет два полюса: проксимальный полюс (формирующийся, цис-поверхность) и дистальный (зрелый,
транс-поверхность). Цис-полюс – сторона мембраны с которой сливаются пузырьки. Транс-полюс – сторона мембраны от которой пузырьки отпочковываются.
Пять функциональных компартментов АГ:
1. Промежуточные везикуло-тубулярные структуры (VTC или ERGIC — ER-Golgi intermediate compartment)
2. Цис-цистерна (cis) — цистерны расп ближе к ЭР:
3. Срединные (medial) цистерны – центральные цистерны
4. Транс-цистерна (trans) — наиболее удаленные от ЭР цистерны.
5. Тубулярная сеть, примыкающая к трансцистерне — транссеть Гольджи (TGN)
Стопки цистерн изогнуты, так что вогнутая трансповерхность обращена к ядру.
В среднем в АГ 3-8 цистерн, в активно секретирующих клеток может быть больше (в экзокринных клетках поджелудочной железы до 13).
Каждая цистерна имеет цис и транс поверхности. Синтезированные белки, мембранные липиды, гликозилированные в ЭР, попадают в АГ через цис-полюс. Вещества через стопки передаются транспортными
пузырьками отделяющиеся от ампул. При прохождении белков или липидов через стопки Гольджи, они претерпевают серию посттрансляционных модификаций, включающих изменение N-связанных олигосахаридов:
цис: маннозидазаI подравнивает длинные маннозные цепи до М-5
промежуточный: N-ацетилглюкоэаминтрансферазаI переносит N-ацетилглюкозамин
транс: добавляются концевые сахара –остатки галактозы и сиаловая к-та.
Строение Аппарата Гольджи и схема транспорта.
Пять компанентов АГ и схема транспорта: промежуточный (ERGIC), цис, промежуточный, транс и транссеть Гольджи (TGN). 1. Вход синтезированных белков, мембранных гликопротеинов и лизосомных ферментов в цистерну переходного ЭР, прилегающую к АГ и 2 — их выход из ЭР в пузырьках окаймленных COPI (антероградный транспорт). 3 — возможный транспорт карго от тубуло-везикулярных
кластеров к цис-цистерне АГ в пузырьках COPI; 3* — транспорт карго от более ранних к более поздним цистернам; 4 — возможный ретроградный везикулярный транспорт карго между цистернами АГ; 5 — возврат резидентных протеинов из АГ в tER с помощью пузырьков, окаймленных COPI (ретроградный транспорт); 6 и 6* — перенос лизосомных ферментов с помощью окаймленных клатрином пузырьков соответственно в ранние EE и поздние LE эндосомы; 7 — регулируемая секреция секреторных гранул; 8 — конститутивное встраивание мембранных белков в апикальную плазматическую мембрану ПМ; 9 — опосредованный рецептором эндоцитоз с помощью окаймленных клатрином пузырьков; 10 возвращение ряда рецепторов из ранних эндосом в плазматическую мембрану; 11 — транспорт лигандов из EE в LE и и лизосомы Ly; 12 — транспорт лигандов в неклатриновых пузырьках.
Функции АГ
1. Транспорт — через АГ проходят три группы белков: белки периплазматической мембраны, белки, предназначенные
на экспорт из клетки, и лизосомные ферменты.
2. Cортировка для транспорта: сортировка для дольнейшего транспорта к органеллам, ПМ, эндосомам, секреторным пузырькам происходит в транс-комплексе Гольджи.
3. Секреция — секреция продуктов, синтезируемых в клетке.
3. Гликозилирование белков и липидов: гликозидазы удаляют остатки сахаров — дегликозилирование, гликозилтрансферазы прикрепляют сахара обратно на главную углеводную цепь — гликозилирование.В нем происходят гликозилирование олигосахаридных цепей белков и липидов, сульфатирование ряда ахаров и тирозиновых остатков белков, а также активация предшественников полипептидных гормонов и нейропептидов.
4. Синтез полисахаридов — многие полисахариды образуются в АГ в том числе пектин и гемицеллюлоза, образующие клеточные стенки растений и большинство гликозаминогликанов образующих межклеточный матрикс у животных
5. Сульфатирование — большинство сахаров, добавляемых к белковай сердцевине протеогликана, сульфатируются
6. Добавление маннозо-6-фосфата: М-6-P добавляется как направляюций сигнал к ферментам, предназначенным для лизосом.
ГЛИКОЗИЛИРОВАНИЕ
Большинство белков начинает гликозилироваться в шероховатом ЭР посредством добавления к растущей полипептидной цепи N-связанных олигосахаридов. Если гликопротеин свернут в нужной конформации, он выходит из ЭР и направляется в АГ, где происходит его посттрансляционная модификация.
В гликозилировании секретируемых продуктов принимают участие ферменты — гликозилтрансферазы. Они участвуют в ремоделированиии Т-связанных боковых олигосахаридных цепей и добвлении О-связанных гликанов и олигосахаридных частей протеогликанов гликолипидов.В модификации олигосахаридов участвуют фрменты а-маннозидаза I и II, которые также являются резидентными белками АГ.
Кроме того в АГ происходит гликозилирование липидно-протеиновых мембранных доменнов, называемых рафтами.
Долихолфосфат добавляет углеводный комплекс – 2GlcNAc-9-манноз-3-глюкозы к аспарагину растущего полипептида. Терминальная глюкоза отщепляется в два этапа: глюкозидаза I отщепляет терминальный остаток глюкозы, глюкозидаза II удаляет еще два остатка глюкозы. Затем отщепляется манноза. На этом начальный этап процессинга углеводов в ЭР завершается и белки несущие олигосахаридный комплекс, поступают в АГ
В первых цистернах АГ удаляются еще три остатка маннозы. На этой стадии стержневой комплекс имеет еще 5 маннозных остатков. N-ацетилглюкозаминтрансфераза I добавляет один остаток N-ацетилглюкозамина GlcNAc. От образовавшегося комплекса отщепляется еще 3 остатка маннозы. Состоит теперь из двух молоекул GlcNAc-3-маннозо-1-GlcNAc является стержневой структурой, к которой гликозилтрансферезы добавляют другие
углеводы. Каждая гликозилтрансфераза распознает развивающуюся углеводную структуру и добавляет к цепи свой собственный сахарид.
СЕКРЕЦИЯ
Схема секреции:
Синтезированные в ЭР белки концентрируются в сайтах выхода переходного ЭР благодаря активности коатомерного комплекса COPII и сопутствующих компонентов и транспортируются в промежуточный между ЭР и АГ компартмент ERGIC, из которого они переходят в АГ в отпочковывающихся пузырьках, или по тубулярным структурам. Белки ковалентно модифицируются, проходя через цистерны АГ, на транс-поверхности АГ сортируются и отправляются к местам своего назначения. Секреция белков требует пассивного встраивания новых мембранных компонентов в плазматическую мембрану. Для восстановления баланса мембран служит контитутивный рецепторопосредованный эндоцитоз.
Эндо и экзоцитозный пути переноса мембран имеют общие закономерности в направленности движения мембранных переносчиков к сооответствующей
мишени и в специфичности слияния и почкования. Основным местом встречи этих путей является АГ.
Источник: cellbiol.ru
Комплекс (аппарат) Гольджи
Как и в любой другой клетке, в мышечном волокне имеется органелла общего назначения, известная под названием комплекса или аппарата Гольджи. Эту органеллу обнаружил в 1889 году итальянский ученый Камилло Гольджи. В саркоплазме мышечного волокна таких органелл насчитывается тысячи.
Строение комплекса Гольджи
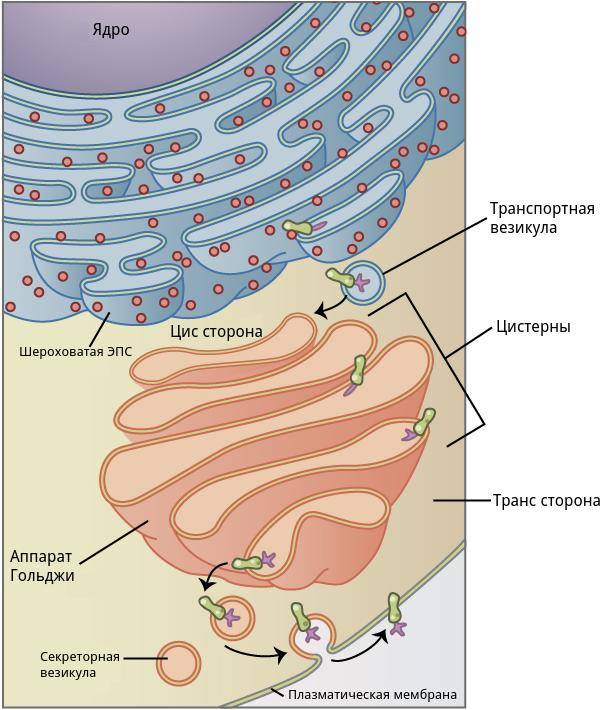
Комплекс Гольджи представляет собой стопку дискообразных мембранных цистерн (рис.1) и связанную с ними систему пузырьков (везикул). Между цистернами имеются связывающие их трубчатые структуры.
Рис.1. Комплекс Гольджи
Комплекс Гольджи расположен недалеко от ядер мышечного волокна и шероховатой эндоплазматической сети (рис.2).
Рис.2. Структура и функции комплекса Гольджи
В комплексе Гольджи различают три отдела: цис-сеть Гольджи, некомпактные зоны и транс-сеть Гольджи.
Цис-сеть Гольджи расположена перед первой цистерной, ближе к ядру мышечного волокна. В транс-сеть Гольджи, переходит последняя цистерна комплекса. Она расположена на более удаленном расстоянии от ядра мышечного волокна. Некомпактные зоны расположены между соседними стопками аппарата Гольджи.
Функции комплекса Гольджи:
- Перенос и преобразование белков;
- Сборка мембран;
- Транспорт веществ к клеточной мембране;
- Формирование лизосом
Перенос и преобразование белков
Необходимые мышечному волокну белки синтезируются на рибосомах. Затем они перемещаются в шероховатую эндоплазматическую сеть. Эти белки называют «незрелыми», потому что они еще не приняли свою пространственную структуру (то есть не свернулись определенным образом). Из шероховатой эндоплазматической сети белки перемещаются в виде мембранных пузырьков в цис-сеть комплекса Гольджи. Известно, что в цистернах, располагающихся ближе к ядру мышечного волокна, содержатся наименее зрелые белки. В цистернах аппарата Гольджи белки «созревают», то есть приобретают свою трехмерную структуру. Биохимики называют этот этап процессингом белка. Каким образом созревающие белки перемещаются по цистернам комплекса Гольджи до сих пор непонятно.
Более подробно строение и функции мышц описаны в моих книгах «Гипертрофия скелетных мышц человека» и «Биомеханика мышц«
В конце концов от цистерн, расположенных достаточно далеко от ядер отпочковываются пузырьки, содержащие полностью «зрелые» белки. С помощью мембранных пузырьков эти белки доставляются «по адресу» в зависимости от полученных ими в аппарате Гольджи «меток». Образно говоря, в мышечном волокне комплекс Гольджи играет роль почты, в которой сортируются и оправляются по адресам различные «посылки» в виде белков, заключенных в пузырьки.
Сборка мембран
В комплексе Гольджи происходит также сборка мембран. Вещества, из которых состоят мембраны (белки, липиды) поступают в комплекс Гольджи из эндоплазматической сети. Затем в цистернах комплекса Гольджи собираются участки мембран, из которых изготавливаются мембранные пузырьки. Они перемещаются в саркоплазме мышечного волокна в те места, где нужно достроить мембрану.
Транспорт веществ к клеточной мембране
Часть веществ, синтезированных в мышечном волокне, выводится наружу. Эти вещества накапливаются в комплексе Гольджи, упаковываются в мембранные пузырьки и транспортируются к сарколемме и выводятся за её пределы.
Источник: allasamsonova.ru
Строение аппарата Гольджи
Аппарат Гольджи обычно локализуется вблизи клеточного ядра. В животных клетках он часто располагается вокруг клеточного центра, образованного парой центриолей.
Аппарат Гольджи состоит из цистерн, сгруппированных в отдельные стопки Гольджи – диктиосомы.
Одна диктиосома в типичном случае содержит 6 цистерн, но в низших эукариотах их количество может возрастать до 30. Количество стопок Гольджи на одну клетку зависит от типа клеток и колеблется в пределах от одной до нескольких сотен. С диктиосомами всегда ассоциируется значительное количество маленьких (примерно 50 нм) пузырьков, ограниченных мембраной. Они группируются на стороне диктиосомы, которая приближена к эндоплазматическому ретикулуму, а также у расширенных краев каждой цистерны. В специализированных секреторных клетках, кроме маленьких пузырьков, могут образовываться крупные секреторные пузырьки или вакуоли. Они часто расположены на той стороне диктиосомы, которая возвращена к плазматической мембране.
Аппарат Гольджи асимметричен, он имеет две различные стороны:
- цис-сторону, которая формируется;
- транс-сторону, или зрелую сторону.
Цистерны стопки Гольджи изогнуты в цис-сторону. Цис-сторона расположена со стороны шероховатого ЭПР и тесно связана с ним. Транс-сторона направлена в сторону плазматической мембраны. Две стороны аппарата Гольджи отличаются не только пространственным расположением, но и по биохимическим свойствам. С помощью специальных гистохимических тестов и электронной микроскопии было установлено, что различные виды ферментов концентрируются в различных частях аппарата Гольджи.
Функции аппарата Гольджи
По своим функциям аппарат Гольджи – главный регулятор, который управляет движением макромолекул в клетке.
Во время прохождения макромолекул в аппарате Гольджи происходит их ковалентная модификация (процессинг). Особенно значительные перестройки происходят в углеводных частях гликопротеидов. Так, в клетках растений формируются фрагменты целлюлозной клеточной стенки, а в клетках членистоногих – хитиновые кутикулы внешнего скелета.
Цистерны шероховатого эндоплазматического ретикулума и аппарата Гольджи являются основными транспортными артериями, которыми белки движутся из клетки за ее пределы.
Аппарат Гольджи есть во всех клетках.
В некоторых специализированных клетках он может занимать значительную часть клеточного объема (например, в бокаловидных клетках кишечника, которые секретируют слизь).
Вакуоли и пузырьки, которые отщепляются от аппарата Гольджи, выносят вещества за пределы плазматической мембраны путем экзоцитоза. Такие пузырьки и вакуоли – это не только средство транспорта, но и кусочки мембраны. Для сохранения целостности и устойчивого размера мембран в клетке существует противоположно направленный эндоцитоз. Он удаляет из плазматической мембраны избыток площади. В растительных клетках формируется одна большая вакуоль, которая называется центральной. Это место накопления продуктов фотосинтеза в растворимом виде. Специфические функции выполняют вакуоли одноклеточных организмов. Некоторые из них содержат комплекс литических ферментов и переваривают питательные вещества. Это пищеварительные вакуоли. В случае, когда содержание солей в клетке превышает таковое в среде обитания, избыток воды из одноклеточного организма выводится через специальные сократительные вакуоли.
Источник: studwork.org