Где происходит фотосинтез у цианобактерий
- Световая фаза фотосинтеза
- Циклический транспорт электронов
- Фотофосфорилирование и окислительное фосфорилирование
- Темновая фаза фотосинтеза
- Цикл Кальвина
- Фотодыхание
- C4-фотосинтез
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
6CO2 + 6H2O → C6H12O6 + 6O2
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.

Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
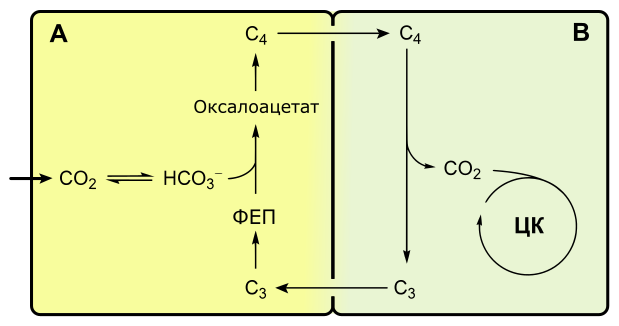
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su
Цианобактерии — изобретатели оксигенного фотосинтеза и создатели кислородной атмосферы Земли — оказались еще более универсальными «биохимическими фабриками», чем ранее считалось. Выяснилось, что они могут совмещать в одной и той же клетке фотосинтез и фиксацию атмосферного азота — процессы, ранее считавшиеся несовместимыми.
Цианобактерии, или, как их раньше называли, синезеленые водоросли, сыграли ключевую роль в эволюции биосферы. Именно они изобрели наиболее эффективный вид фотосинтеза — оксигенный фотосинтез, идущий с выделением кислорода. Более древний аноксигенный фотосинтез, идущий с выделением серы или сульфатов, может происходить только в присутствии восстановленных соединений серы (таких как сероводород) — веществ достаточно дефицитных. Поэтому аноксигенный фотосинтез не мог обеспечить производство органики в количестве, необходимом для развития разнообразных гетеротрофов (потребителей органики), включая животных.
Цианобактерии научились использовать вместо сероводорода обычную воду, что обеспечило им широкое распространение и огромную биомассу. Побочным результатом их деятельности стало насыщение атмосферы кислородом. Без цианобактерий не было бы и растений, ведь растительная клетка — результат симбиоза нефотосинтезирующего одноклеточного организма с цианобактериями. Все растения осуществляют фотосинтез при помощи особых органелл — пластид, которые суть не что иное, как симбиотические цианобактерии. И не ясно еще, кто главный в этом симбиозе. Некоторые биологи говорят, пользуясь метафорическим языком, что растения — всего лишь удобные «домики» для проживания цианобактерий.
Цианобактерии не только создали биосферу «современного типа», но и по сей день продолжают ее поддерживать, производя кислород и синтезируя органику из углекислого газа. Но этим не исчерпывается круг их обязанностей в глобальном биосферном круговороте. Цианобактерии — одни из немногих живых существ, способных фиксировать атмосферный азот, переводя его в доступную для всего живого форму. Азотфиксация абсолютно необходима для существования земной жизни, а осуществлять ее умеют только бактерии, и то далеко не все.
Главная проблема, с которой сталкиваются азотфиксирующие цианобактерии, состоит в том, что ключевые ферменты азотфиксации — нитрогеназы — не могут работать в присутствии кислорода, который выделяется при фотосинтезе. Поэтому у азотфиксирующих цианобактерий выработалось разделение функций между клетками. Эти виды цианобактерий образуют нитевидные колонии, в которых одни клетки занимаются только фотосинтезом и не фиксируют азот, другие — покрытые плотной оболочкой «гетероцисты» — не фотосинтезируют и занимаются только фиксацией азота. Эти два типа клеток, естественно, обмениваются между собой производимой продукцией (органикой и соединениями азота).
До недавнего времени считалось, что совместить фотосинтез и азотфиксацию в одной и той же клетке невозможно. Однако 30 января Артур Гроссман и его коллеги из Института Карнеги (Вашингтон, США) сообщили о важном открытии, показывающем, что ученые до сих пор сильно недооценивали метаболические способности цианобактерий. Оказалось, что живущие в горячих источниках цианобактерии рода Synechococcus (к этому роду относятся примитивные, древние, чрезвычайно широко распространенные одноклеточные цианобактерии) ухитряются совмещать в своей единственной клетке оба процесса, разделяя их во времени. Днем они фотосинтезируют, а ночью, когда концентрация кислорода в микробном сообществе (циано-бактериальном мате) резко падает, переключаются на азотфиксацию.
Открытие американских ученых не стало полной неожиданностью. В прочтенных за последние годы геномах нескольких разновидностей Synechococcus были обнаружены гены белков, связанных с азотфиксацией. Не хватало только экспериментальных подтверждений того, что эти гены действительно работают.
Таким образом, удалось выяснить, откуда берут азот термофильные микробные маты, живущие при температурах, непригодных для роста обычных нитчатых азотфиксирующих цианобактерий с гетероцистами. Кроме того, открытие позволяет по-новому взглянуть на древнейшие этапы развития микробной жизни на нашей планете. Ведь первые известные в ископаемом состоянии остатки живых организмов (их возраст — около 3,5 млрд лет) напоминают одноклеточных цианобактерий, близких к Synechococcus.
Источник: Hot-Spring Bacteria Flip a Metabolic Switch.
См. также:
Б. В. Громов. Цианобактерии в биосфере.
Ископаемые бактерии.
Зарождение жизни. Прокариотная биосфера.
Александр Марков
Источник: elementy.ru
В зависимости от того, какой источник энергии могут использовать прокариоты, их делят на фототрофов (источник энергии — свет) и хемотрофов (источник энергии — окислительно-восстановительные реакции).
Организмы, у которых источниками (донорами) электронов в энергетическом процессе являются неорганические вещества, предложено называть литотрофными , а те, у которых донорами электронов служат органические соединения, — органотрофными .
Фотосинтетический аппарат состоит из трех основных компонентов:
— светособирающих пигментов, поглощающих энергию света и передающих ее в реакционные центры;
— фотохимических реакционных центров, где происходит трансформация электромагнитной формы энергии в химическую;
— фотосинтетических электронтранспортных систем, обеспечивающих перенос электронов, сопряженный с запасанием энергии в молекулах АТФ .
В фотохимической реакции участвуют, как правило, хлорофиллы или бактериохлорофиллы а в модифицированной форме. Эти же виды хлорофиллов наряду с другими, а также пигментами иных типов ( фикобилипротеины , каротиноиды ) выполняют функцию антенны. У некоторых пурпурных бактерий , содержащих только бактериохлорофилл b , он выполняет обе функции. У гелиобактерий бактериохлорофилл g также служит светособирающим пигментом и входит в состав реакционного центра ( табл. 21 ).
Два компонента фотосинтетического аппарата — реакционные центры и электронтранспортные системы — всегда локализованы в клеточных мембранах, представленных ЦПМ и у большинства фотосинтезирующих эубактерий развитой системой внутрицитоплазматических мембран — производных ЦПМ ( рис. 4 ). Локализация светособирающих пигментов в разных группах фотосинтезирующих эубактерий различна ( табл. 22 ). У пурпурных бактерий , гелиобактерий и прохлорофит светособирающие пигменты в виде комплексов с белками интегрированы в мембраны ( рис. 72 , А). В клетках зеленых бактерий и цианобактерий основная масса светособирающих пигментов находится в особых структурах, прикрепленных к поверхности мембраны, но не являющихся ее компонентом. Это хлоросомы зеленых бактерий и фикобилисомы цианобактерий ( рис. 4 ).
В хлоросомах зеленых бактерий содержится весь бактериохлорофилл с, d или е (в зависимости от вида), а также небольшие количества бактериохлорофилла а, служащего промежуточным звеном при переносе энергии света от основного светособирающего бактериохлорофилла к бактериохлорофиллу а, локализованному в ЦПМ . С этой формы пигмента энергия света передается на модифицированную форму бактериохлорофилла а реакционного центра. Локализованные в хлоросомах светособирающие бактериохлорофиллы организованы в виде палочковидных структур диаметром 5-10 нм, расположенных параллельно длинной оси хлоросомы ( рис. 72 , Б), Их высокоупорядоченная организация и упаковка осуществляются с помощью белковых молекул. В основании хлоросомы, примыкающем к ЦПМ, расположен слой молекул бактериохлорофилла а.
Фотосинтез у цианобактерий: характеристика фототрофов, морфология и локализация фотосинтетического аппарата, механизм фотосинтеза.
Цианобактерии:
Oxyphotobacteria
У них грамотриц. кл. стенка, много слизи или чехол, нет жгутиков, бывают газовые вакуоли, соседние клетки могут быть соединены микроплазмодесмами.
Фотосинтетический аппарат:
Рисунок:
Участие микроорганизмов в круговороте азота в природе. Азотфиксация.
Встречается в воздухе (80%), в почве, в воде, органический азот в белках и нуклеотидах.
Этапы цикла: азотфиксация, аммонификация, нитрификация, денитрофикация, ассимиляция.
Азотфиксация.
1. Свободноживущие азотфиксаторы: цианобактерии, строгие анаэробы, Azotobacter Beijerinkia, цианобактерии, фототрофы (строгие аэробы).
Микроорганизмы, живущие в симбиозе с высшими растениями.
А.Бобовые Rhizobium: аэробы, нуждающиеся в микроэлементах, неспорообразующие.
Обладают экзоферментом (выдается наружу галактоуронедаза).
Разрушают клеточную стенку волоска. В корневолоске образуют бактероидную нить. Внутри клубенька бактероидная зона, снаружи клетки корня. Там протекает азотфиксация. Фермент нитрогеназа.
Б. Небобовые – актиномицеты.
Участие микроорганизмов в круговороте азота в природе. Денитрификация и ассимиляция.
Важнейший элемент, входящий в состав белков, а следовательно, имеющий исключительное значение для жизни — это азот. В живых существах, населяющих планету, содержится примерно 15—20 млрд. т азота, в почвах (в 30-сантиметровом слое) на каждом гектаре имеется в среднем 5—15 т азота.
Круговорот азота включает следующие процессы:
— фиксация азота;
— аммонификация;
— нитрификация;
— денитрификация.
Денитрификация, протекающая под воздействием микробов, представляет собой восстановление нитратов с образованием в качестве • конечного продукта — молекулярного азота, возвращающегося из почвы в атмосферу. Вызывается этот процесс денитрифицирующими бактериями. Наиболее распространенные из них в природе: Tiolacillus denitrifi-cans — палочка, не образующая спор, факультативный анаэроб; Ps. fluo-rescens — подвижная палочка, выделяет зеленоватый пигмент, быстро разлагает нитраты; Ps. aeruginosa — бактерия сходна с предыдущей; Ps. Stutzeri — небольшая палочка, образующая цепочки, разлагает нитраты в анаэробных условиях.
Ассимиляция – извлечение бактериями азота из нитратов для синтеза азотосодержащих клеточных компонентов нитратредуцирующих в нитратредукторов.
NO3 – NO2 – NH3
Участие микроорганизмов в круговороте азота в природе. Нитрификация и
Аммонификация.
Аммонификация – процесс превращения органического азота (N) в NH4. (Гниение, минерализация)
Белки: экзопротеазы.
Нуклеиновые кислоты: экзонуклеазы.
Бактерии: Bacillus, Pseudomonas, Micromycetes грамполож.
Аммонификация мочевины
Уробактерии образуют фермент уреазу. Под действием фермента уреазы происходит гидролиз мочевины с образованием карбоната аммония, который почти тотчас разлагается на составные компоненты – NH3, H2O, CO2.
H2N-CO-NH2 + H2O → (NH4)2CO3 → 2NH3 + H2O + CO2
Бактерии, разлагающие мочевину, получили название уробактерий. К ним относятся Sporosarcina ureae, Micrococcus ureae, Bacillus pasteuri и Baccilus probatus.
В качестве азота они используют аммиачные соли или свободный аммиак, образующийся при гидролизе мочевины. Углерод из мочевины уробактерии использовать не могут, так как он находится в сильно окисленной форме и при гидролизе не выделяется в виде углерода диоксида. Углерод уробактерии используют из различных органических соединений (соли лимонной, янтарной, яблочной, уксусной и других кислот, а также моносахариды, сахариды и крахмал).
Нитрификация – окисление аммония (NH3) до нитрата (NO3).
В почве является нежелательным процессом.
Проходит в 2 стадии
Первая стадия — окисление аммиака до нитрит-аниона
, которое осуществляют нитрозные бактерии родов Nitrosomonas, Nitrosococcus и Nitrosospira (ранее выделялись также рода Nitrosolobus, Nitrosovibrio, но сейчас их представители включены в Nitrosospira ) по следующему механизму:
1. NH3 + O2 + НАДН2 → NH2OH + H2O + НАД+
2. NH2OH + H2O → HNO2 + 4H+ + 4e−
3. 1/2O2 + 2H+ + 2e− → H2O
Ферментом для первой реакции служит аммиакмонооксигеназа. Гидроксиламин ингибирует работу фермента. В бесклеточных экстрактах восстановителем может служить НАД(Ф)·H.
Следующую реакцию осуществляет гидроксиламиноксидоредуктаза. Окислителем в них служит цитохром c, с него электрон передаётся на убихинон и далее в дыхательную цепь, на цитохромоксидоредуктазу и, в конечном итоге, на кислород. При этом запасается энергия в виде трансмембранного протонного потенциала.
Образование НАД(Ф)·H для фиксации углекислого газа в цикле Кальвина происходит путём обратного переноса части электронов.
Вторая стадия — окисление аниона азотистой кислоты до аниона азотной, производимое нитратными бактериями (почвенный род Nitrobacter и водные Nitrospira, Nitrococcus, Nitrospina). Процесс протекает в одну реакцию:
NO2− + H2O → NO3− + 2H+ + 2e−
катализируемую нитрит:нитрат-оксидоредуктазой, локализованной в ЦПМ. Далее электроны передаются на цитохромы дыхательной цепи, в которой единственным пунктом транслокации протонов является цитохромоксидаза.
Образование НАД(Ф)·H для фиксации углекислого газа также происходит путём обратного переноса электронов.
Участвуют 2 типа бактерий: нитроазотные – грамотриц., полиморфные, подвижные (образуют слизь), аэробы; нитратные – грамотриц., полиморфные.
Источник: lektsia.com