Пиноцитозные вакуоли функции
Инфузория туфелька: где обитает, строение и функции
Инфузория-туфелька – что это?
Инфузория-туфелька – это простейший одноклеточный микроорганизм, который получил свое название за внешнее сходство с обувной подошвой. Ее размеры колеблются от 10 мкм до 4,5 мм, но такие крупные особи встречаются крайне редко. В основном можно встретить в пресных и стоячих водах.
Невооруженным глазом этот микроорганизм невозможно рассмотреть. Однако при большом скоплении в загрязненной и мутной воде можно увидеть продолговатые точки белого цвета – это и есть инфузории-туфельки. Они находятся в постоянном движении.

Инфузория-туфелька – это бактерия или нет?
Бактерия – одноклеточный организм, отличающийся отсутствием ядра, а инфузория-туфелька обладает двумя ядрами. Из этого можно сделать вывод, что данный представитель фауны не является бактерией.
Где обитает инфузория-туфелька?
Как уже говорилось выше, инфузория-туфелька обитает в пресноводных водоемах. Изучив воду из домашнего аквариума под микроскопом, можно заметить большое количество микроорганизмов, в том числе и инфузорий.
Можно самостоятельно создать искусственный водоем, в котором будет обитать этот простейший одноклеточный организм, для этого будет достаточно залить водой обычное сено, и дать настояться несколько дней.
Строение и функции
Внешнее покрытие этого представителя фауны представляет собой тонкую эластичную оболочку, которая называется мембраной. Она на протяжении всего жизненного цикла сохраняет форму тела. Это связано с наличием в слое цитоплазмы развитых опорных волокон. Эти волокна расположены плотно к оболочке. Инфузория-туфелька обладает двумя ядрами. За пищеварение отвечает большое ядро, а за размножение – малое.
На всей поверхности инфузории-туфельки расположены органы, отвечающие за ее передвижение. Эти органы называются ресничками, и их количество превышает 15 000. Их движения напоминают движения весел. Перемещение инфузории осуществляется тупым концом вперед со скоростью до 3 мм/с. Во время передвижения этот микроорганизм вращается вокруг продольной оси своего тела. Это происходит за счёт медленных волнообразных движений ресничек.
Инфузория-туфелька – это высокоорганизованный простейший организм, который выполняет множество процессов для поддержания своей жизнедеятельности.
Дыхание организма осуществляется за счет попадания кислорода в цитоплазму через мембрану. Благодаря двум сократительным вакуолям, происходит газообмен, осуществляющийся за счет специальных канальцев. Удаление лишней жидкости, которая представляет собой результат процесса жизнедеятельности, происходит каждые 30 секунд. При неблагоприятной окружающей среде происходит замедление работы сократительных вакуолей, и инфузория-туфелька перестает питаться.
Размножение этого высокоорганизованного микроорганизма может быть как половым, так и бесполым.
Бесполое размножение у инфузорий-туфелек представляет собой обычный процесс деления клеток. Примерно раз в день большое и малое ядра расходятся в разные стороны организма и делятся на два. В результате деления образуется две инфузории-туфельки с таким же набором органов, как и у родительского организма.
Половое размножение свойственно только тем инфузориям, которые многократно проходили бесполое размножение или же при неблагоприятных условиях. В результате этого размножения не образуется двух особей. Два микроорганизма соединяются, создавая между собой соединительный мост.
Чем питается?
Рацион питания инфузории-туфельки состоит из бактерий и микроводорослей, которые содержатся в большом количестве в мутной застоявшейся воде. Питание происходит с помощью клеточного рта, по кругу которого расположены реснички. С их помощью микроорганизм может с легкостью захватывать как можно больше еды в рот. Изо рта пища проходит по клеточной глотке, попадая в вакуоли, в которых и происходит процесс пищеварения. Он может происходить в нескольких вакуолях сразу, и может длиться более часа.
Инфузория туфелька может питаться непрерывно, особенно когда температура воды более 17 градусов, прерываясь только для размножения.
Опасна ли для человека?
Существует множество инфузорий, которые могут паразитировать в организме рыб и даже человека. Например, заболевание балантидиаз связано с присутствием балантидия кишечного в организме человека. А ихтиофтириус паразитирует в аквариумных рыбах, вызывая их массовую гибель. Однако инфузория-туфелька не представляет никакой угрозы для их здоровья. Она служит основной пищей для беспозвоночных и мальков рыб.
Заключение
Строение и внешний вид инфузорий-туфелек одинаковы для каждой особи. Они могут отличаться размерами. Жизненный цикл при благоприятных условиях у них тоже одинаковый. Эти микроорганизмы остро реагируют на температуру воды, освещение и содержание солей в водоеме. При неблагоприятных условиях они впадают в анабиоз, и их длительность жизни увеличивается в сотни, а то и в тысячи раз.
Источник: internat-barnaul.ru
ный транспорт множества органических и неорганических молекул, участвует в генерации потенциала действия нервных и мышечных элементов. Благодаря работе натрий-калиевого насоса в клетках поддерживается высокая концентрация ионов калия (140мМ) и низкая – ионов натрия (12мМ), между тем как в крови и межклеточной жидкости соотношение этих концентраций – обратное. Работа Na+/K+ насоса создает разность потенциалов в 50-70мB (плюс
– снаружи плазматической мембраны, минус – внутри). Существуют многочисленные белки-переносчики и ионные каналы. Хорошим примером может служить белок-переносчик, обеспечивающий двунаправленное (из клетки и в клетку) движение анионов (Cl— и HCO—3) через эритроцитарную мембрану. Перенос глюкозы в клетку также осуществляют интегральные гликопротеины. Транспорт аминокислот также осуществляется с помощью белковпереносчиков. Обратное всасывание фосфатов происходит в проксимальных канальцах нефрона при помощи двух кодируемых генами NPТ2 переносчиков. Дефекты этих генов являются причиной некоторых болезней, в том числе рахита, который невозможно вылечить витамином Д.
Активный транспорт позволяет клетке концентрировать питательные вещества, находящиеся во внешней среде в весьма низких концентрациях.
В ряде случаев макромолекулы и их агрегаты, а часто и крупные частицы попадают в клетку, либо покидают ее путем эндоили экзоцитоза. Эндоцитоз и экзоцитоз – это два активных процесса, посредством которых различные материалы транспортируются через мембрану либо в клетки (эндоцитоз), либо из клеток (экзоци-
тоз).
Эндоцитоз – это везикулярный перенос жидкостей, макромолекул или небольших частиц и микроорганизмов (вирусов, бактерий) в клетку. Существует несколько механизмов эндоцитоза:
1.Пиноцитоз – («клеточное питье»).
2.Рецепторно-опосредованный эндоцитоз или клатрин-
зависимый эндоцитоз.
3.Фагоцитоз – («клеточная еда»).
Пиноцитоз – это постоянный процесс поглощения жидкости и растворенных веществ с образованием небольших пузырьков (пиноцитозные пузырьки). Эти пузырьки переносят небольшие моле-
кулы, воду и растворимые белки, то есть вещества, относящиеся к жидкой фазе внеклеточной среды. Благодаря такому процессу, клетки могут поглощать как крупные молекулы, так и ионы, неспособные проникнуть через мембрану. В цитоплазме клетки многие пиноцитозные пузырьки сливаются друг с другом и формируют ранние эндосомы, которые смещаются в глубь клетки и сливаются с лизосомами; в их полости начинается разрушение веществ. В зависимости от размеров пиноцитозных пузырьков выделяют макро- и микропиноцитоз. Несмотря на маленькие размеры пиноцитозных пузырьков, их многочисленность позволяет им доставлять в клетку большое количество веществ (особенно интенсивно пиноцитоз протекает в эндотелиальных клетках кровеносных капилляров).
Рецепторно-опосредуемый эндоцитоз характеризуется по-
глощением из внеклеточной жидкости конкретных макромолекул. Для этой цели клетка экспрессирует на свою поверхность специфические поверхностные рецепторы, что обеспечивает избирательное связывание молекул во внеклеточном растворе. Рецепторы, способные связывать лиганд (специфическую сигнальную молекулу), накапливаются в специфических участках поверхности клетки, называемых окаймленными ямками. Поверхность этих небольших углублений покрыта с цитозольной стороны плазматической мембраны белком клатрином. При связывании лиганда с рецептором под плазматической мембраной формируется пузырек – ранняя эндосома, содержащий комплекс лиганд-рецептор, покрытый снаружи клатрином. Белок клатрин предотвращает слияние эндосом с лизосомами и переваривание лиганда ферментом. В пузырьках такого типа лиганд доставляется в различные отделы в пределах одной клетки или переносится в другой слой клеток. Такой тип переноса называют трансцитозом. Известны четыре типа трансцитоза. Первый тип – рецептор возвращается в плазматическую мембрану, а лиганд разрушается (инсулин, липопротеиды низкой плотности, некоторые вирусы и др.). Второй тип – рецептор возвращается и лиганд возвращается (белок, переносящий железо и др.). Третий тип трансцитоза характеризуется разрушением лиганда и рецептора (эпидермальный фактор роста и др.). Четвертый тип – его механизм связан с транспортом и лиганда, и рецептора (материнские антитела переносятся в клетки молочной железы, а затем у новорожден-
ного из молока они проходят через эпителий кишечника и попадают в лимфатические протоки и кровь).
Нарушения различных типов рецепторно-опосредованного эндоцитоза сопровождаются серьезными заболеваниями человека (семейная гиперхолестеринемия, приводящая к образованию атеросклеротических бляшек в сосудах, является примером неполноценного трансцитоза первого типа).
Фагоцитоз – поглощение крупных твердых частиц (микроорганизмов, остатков разрушающихся клеток и др.). К фагоцитозу способны все клетки, но наиболее активно фагоцитоз осуществляют профессиональные фагоциты (моноциты, макрофаги, нейтрофилы). В ходе фагоцитоза образуются большие эндоцитозные пузырьки – фагосомы. Фагосомы сливаются с лизосомами и формируют фаголизосомы, в которых происходит переваривание поглощенного материала. Фагоцитоз является важнейшим механизмом защиты организма-хозяина от микроорганизмов.
Экзоцитоз – перенос частиц и крупных соединений из клетки. Наиболее распространенный способ экзоцитоза – секреция. Специализированные клетки хранят секретируемые молекулы (пищеварительные ферменты, гормоны, нейромедиаторы) в пузырьках, расположенных вблизи плазматической мембраны), а при получении внешнего сигнала происходит слияние пузырьков с плазматической мембраной и освобождение их содержимого.
кой процесс называют стимулированной секрецией. Удаление из клетки твердых частиц называют экскрецией. При такой форме экзоцитоза удаляемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазмолеммой. В клетках иммунной системы осуществляется рекреция – перенос твердых веществ через клетку. При этом процессе с одной стороны клетки происходит фагоцитоз, а с другой – экскреция. Так специализированные макрофаги (дендритные клетки, клетки Лангерганса и др.), локализованные в слизистых оболочках и коже захватывают бактериальные клетки, разрушают их до мелких обломков и представляют (презентируют) эти антигены иммуннокомпетентным лимфоцитам.
ЦИТОПЛАЗМА
СТРУКТУРНЫЕ КОМПОНЕНТЫ ЦИТОПЛАЗМЫ КЛЕТКИ
I. ГИАЛОПЛАЗМА
II. ОРГАНЕЛЛЫ
III. ВКЛЮЧЕНИЯ
I. ГИАЛОПЛАЗМА
Гиалоплазма – это матрикс цитоплазмы, представляющий собой ее истинную внутреннюю среду. Он имеет вид гомогенного или тонкозернистого вещества с низкой электронной плотностью. Является сложной коллоидной системой, включающей в себя различные биополимеры: глобулярные белки (20-25% от общего содержания белков в эукариотической клетке), нуклеиновые кислоты, полисахариды, промежуточные продукты их обмена, воду, неорганические вещества и др. Эта система способна переходить из золеобразного (жидкого состояния) в гелеобразное и обратно.
II. ОРГАНЕЛЛЫ
Органеллы – это постоянные структурные компоненты цитоплазмы клеток, имеющие строго определенное строение и выполняющие жизненно важные специфические функции.
Классификации органелл
I.По строению:
|
А. Мембранные |
Б. Немембранные |
|
— Гранулярная ЭПС |
— Рибосомы |
|
— Гладкая ЭПС |
— Центриоли |
|
— Комплекс Гольджи |
— Микротрубочки и |
|
— Митохондрии |
микрофиламенты |
|
— Лизосомы |
— Протеасомы |
|
— Пероксисомы |
|
II. По распространенности в клетках:
А. Общие органеллы (присутствуют во всех эукариотических клетках): (все мембранные и немембранные органеллы);
Б. Специальные органеллы (присутствуют только в отдельных, высокоспециализированных клетках):
—миофибриллы (мышечные клетки)
—нейрофибриллы (нервные клетки)
—микроворсинки (эпителиоциты кишечника)
—реснички (эпителиоциты дыхательных путей)
—жгутики (сперматозоиды)
А. МЕМБРАННЫЕ ОРГАНЕЛЛЫ Мембранными называются органеллы, в структуре которых
имеются элементарные клеточные мембраны.
Гранулярная эндоплазматическая сеть представлена системой многочисленных мембран, образующих уплощенные мешки, цистерны и трубочки на поверхности которых содержатся рибосомы. Комплекс мембран гранулярной эндоплазматической сети связан с наружной мембраной ядерной оболочки и перинуклеарным пространством. В полостях эндоплазматической сети содержится много различных молекул-компонентов, среди которых имеют большое значение белки шапероны. Шапероны – семейство специализированных внутриклеточных белков, обеспечивающих быстрое и правильное сворачивание (фолдинг) вновь синтезированных молекул белка. Связывание с шаперонами препятствует агрегации с другими белками и тем самым создает условия для формирования вторичной и третичной структуры растущего пептида. Шапероны принадлежат к так называемым белкам теплового шока (hsp 60, hsp 70, hsp 90). Синтез данных белков активируется при стрессах, в частности, при тепловом шоке (отсюда и название – белок теплового шока). Шапероны предотвращают денатурацию белков при высокой температуре и др. экстремальных факторах. Связываясь с аномальными белками, шапероны восстанавливают их нормальную конформацию и тем самым повышают выживаемость организма при резком ухудшении физико-химических параметров среды.
Рисунок 2. Схема строения гранулярной эндоплазматической сети
(1- рибосомы; 2- цистерны).
Функции:
1.Синтез экспортных белков.
2.Синтез белков для мембран клетки.
3.Изоляция данных белков от содержимого гиалоплазмы.
4.Транспорт белков.
Гладкая эндоплазматическая сеть – в основе строения лежат элементарные клеточные мембраны, образующие систему мелких вакуолей, трубок и канальцев. Гладкая сеть является продолжением гранулярной, однако не содержит рибофоринов – гликопротеиновых рецепторов, с которыми соединяется большая субъединица рибосом и поэтому не связана с рибосомами.
Функции:
1.Метаболизм (синтез) липидов.
2.Метаболизм (синтез) углеводов (гликоген в клетках печени).
3.Депонирование ионов кальция (в скелетной и сердечной мышечной ткани).
4.Обезвреживание различных вредных веществ (в клетках печени).
5.Транспортная.
Комплекс Гольджи — состоит из набора расширенных по краям (ампулы) уплощенных цистерн, сложенных в стопку и от-
~36 ~
почковывающихся от цистерн микропузырьков и вакуолей. Каждое такое скопление цистерн называется диктиосомой. Количество цистерн в разных клетках варьирует, чаще всего, в пределах 5-12, между которыми находятся тонкие прослойки гиалоплазмы. Комплекс Гольджи располагается обычно между эндоплазматической сетью и плазматической мембраной. Часть комплекса Гольджи, обращенная к эндоплазматической сети, называется цис-полюсом, а удаленная от ЭС – транс-полюсом. В соответствии с полярностью комплекса Гольджи каждая сторона его цистерн имеет цис- и транс-поверхности.
Рисунок 3. Схема ультраструктурного строения комплекса Гольджи (1- цистерна; 2- вакуоли, 3- микропузырьки; первичные лизосомы)
Функции:
1.Накопление, сортировка и созревание веществ, синтезированных на гладкой и гранулярной эндоплазматической сети.
2.Синтез и сегрегация сложных соединений (полисахаридов, гликопротеидов, липопротеидов).
3.Транспортная.
4.Участвует в гидроосмотическом ответе клетки (в случае больших водных потоков цитоплазма обводняется, и вода частично собирается
вкрупных вакуолях комплекса Гольджи).
5.Образование первичных лизосом.
Митохондрии – это мембранные органеллы размером около 1–2 мкм, обладающие, в отличие от других органелл клеток, сдвоенной липидной мембраной: гладкой – наружной митохондриальной мембраной, отделяющей содержимое митохондрий от гиало-
~37 ~
плазмы, и внутренней митохондриальной мембраны, складчатой. Складки внутренней митохондриальной мембраны образуют поперечные перегородки – кристы, глубоко проникающие внутрь митохондрий и отграничивающие собственное внутреннее содержимое митохондрий – ее матрикс.
Рисунок 4. Схема строения митохондрии (1-
наружная митохондриальная мембрана; 2-внутренняя митохондриальная мембрана; 3-кристы; 4-матрикс митохондрий (рибосомы, кольцевые молекулы ДНК); 5-межмембранное пространство). Из кн. Э. Хадорн, Р. Венер «Общая зоо-
логия», 1989
Различные типы клеток отличаются друг от друга как по количеству и форме митохондрий, так и по количеству крист (много крист имеют митохондрии в сердечной мышце). Обычно в клетке, например, гепатоците, содержится около 2000 митохондрий. Продолжительность жизни митохондрий около 10 дней. Состарившиеся митохондрии узнаются и разрушаются в аутофагосомах. Митохондрии размножаются путем деления, при этом не исключается их образование de novo.
Мембраны митохондрий содержат интегральные мембранные белки. Во внешнюю мембрану входят порины, которые образуют поры и делают мембраны проницаемыми для веществ с молекулярной массой до 10кДа. Внутренняя же мембрана митохондрии непроницаема для большинства молекул; исключение составляют О2, СО2, Н2О. Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%): транспортные белки, ферменты (NO-синтетаза), компоненты дыхательной цепи и АТФсинтаза. Пространство, образованное внешней и внутренней мембранами, обычно называют межмембранным пространством; оно служит резервуаром протонов. Матрикс митохондрий имеет тонкозернистое строение. Он представлен тонкими нитями (молекулы ДНК) и мелкими гранулами (митохондриальные рибосомы) и за-
полнен концентрированным раствором ферментов цикла Кребса. Митохондриальная ДНК у млекопитающих представляет собой кольцевую молекулу, состоящую из 16569 пар нуклеотидов; в каждой митохондрии может быть 5-10 копий ДНК. Митохондриальная хромосома включает 37 генов. Митохондриальный геном всех людей, кроме родственников по женской линии, различен. Это связано с тем, что в митохондриальных генах нет интронов и отсутствуют системы репарации ДНК, вследствие этого мутации мтДНК возникают примерно в 10 раз чаще, чем в ядерных генах. Различия митохондриальной ДНК разных людей дают возможность использовать анализ этой ДНК для генетической идентификации личности и установления родства. В последнее время обнаружена обширная группа болезней, связанных с патологией митохондрии. Сегодня известно более 200 крупных делеций и дупликаций в митохондриальной ДНК, которые оказывают негативное влияние на функционирование этих важнейших органелл. Известно, что инактивация митохондриальных генов в результате мутаций служит причиной различных патологических состояний от наследственной слепоты и глухоты до диабета и старческого слабоумия. Некоторые дефекты митохондрий являются причиной врожденного бесплодия женщин. Все вызванные митохондриальными мутациями болезни передаются по материнской линии. Некоторые ученые считают, что по мере накопления мутаций мтДНК в соматических клетках индивидуума в них идет процесс разбалансирования, который является одной из основных причин старения организма.
Функции:
6.Синтез АТФ — «энергетические блоки».
7.Синтез митохондриальных белков.
8.Образование оксида азота (NO) – в малых концентрациях он стимулирует образование новых митохондрий, а в больших – подавляет их работу.
9.Образование первичных лизосом.
Лизосомы (от греч. lysis – разложение, распад и греч. soma – тело) – окруженные мембраной органеллы (диаметром 0,2- 0,8 мкм), присутствующие в цитоплазме всех эукариотических клеток. Лизосомы образно называют емкостями с «оружием массового поражения», так как внутри них находится целый набор гидроли-
~39 ~
тических ферментов (кислая фосфатаза и др.), способных разрушить любой компонент клетки. Клетку спасает от разрушения только лизосомальная мембрана.
Рисунок 5. Электронная микро-
фотография (1- лизосомы; 2- митохондрии)
Виды лизосом:
1.Первичные
2. Вторичные — фаголизосомы — аутофагосомы 3. Третичные
(остаточные тельца)
—Первичные лизосомы – отпочковываются от аппарата Гольджи в виде пузырьков, отграниченных мембраной, начиненных гидролитическими ферментами.
—Вторичные лизосомы — отграниченные мембраной пузырьки, начиненные гидролитическими ферментами, внутри которых находятся объекты, подлежащие разрушению. Если такими объектами являются состарившиеся или поврежденные другие органеллы (митохондрии), такие пузырьки (вторичные лизосомы) называют аутофагосомами. Если объектами, подлежащими уничтожению, будут являться захваченные извне частицы, такие пузырьки (вторичные лизосомы) называют фаголизосомами (фагосомами).
—Третичные лизосомы (остаточные тельца) — отграниченные мембраной пузырьки, начиненные незначительным количеством гидролитических ферментов, внутри которых накапливаются непереваренные продукты. В этих лизосомах происходит уплотнение содержимого и его перестройка. Пример: у человека при старении организма в некоторых клетках происходит отложение “пигмента старения” – липофусцина.
~40 ~
Источник: studfile.net
Источник: naturalpeople.ru
ЦЕНТРАЛЬНАЯ ВАКУОЛЬ РАСТИТЕЛЬНОЙ КЛЕТКИ.
I. ВВЕДЕНИЕ
Современный этап развития биологии характеризуется исследованиями в ряде приоритетных научных направлений, одним из которых является изучение механизмов мембранного транспорта. Менее всего в настоящее время изучены эти механизмы применительно к специфичной для растений мембране — вакуолярной.
Наличие вакуоли является отличительной особенностью растительной клетки. Её роль в жизнедеятельности растений достаточно велика. В настоящее время показано, что это полифункциональный компартмент, который занимает особое место в структурной и функциональной организации растительного организма. Трудно переоценить роль вакуоли в регуляции клеточного объёма, тургора, а также pH и ионного гомеостаза цитозоля, что обеспечивает работу ферментов цитоплазмы, в изоляции вторичных продуктов метаболизма и метаболических ядов, в трансдукции сигналов различной природы, в процессах апоптоза, в ответных защитных реакциях растения на стрессовое воздействие и т.д. Одна из важнейших функций вакуоли состоит в запасании метаболитов.
Основным метаболитом, запасаемым в растении (в отличие от глюкозы в животной клетке), является сахароза. Процессы синтеза и транспортировки сахарозы по растению в настоящее время изучаются. Но каким бы способом ни транспортировалась сахароза (по симпласту или по апопласту), конечный этап дальнего транспорта — вакуолярная мембрана, которая содержит переносчик сахарозы и протонные помпы, обеспечивающие процесс транспорта. Путь к тонопласту идёт по донорно-акцепторным градиентам, а скорость закачивания сахарозы в вакуоль, которая зависит от активности протонных помп тонопласта, может являться в этом процессе одним из определяющих звеньев.
Различают пищеварительные и сократительные (пульсирующие) вакуоли, регулирующие
осмотическое давление и служащие для выведения из организма продуктов распада.
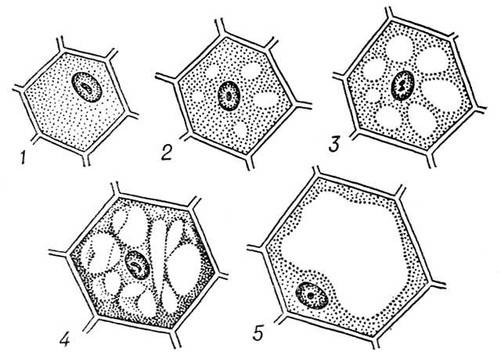
В клетках животных – пиноцитозная вакуоль.
Внутренняя мембрана клетки – тонопласт — окружает вакуоль. Царство одноклеточные, тип простейшие. – группа животных большинство из которой питаются органической пищей (бактерии, одноклеточные водоросли). Захваченные ими оформленные частицы пищи перевариваются в пищеварительных вакуолях – органоидах пищеварения. После чего растворенная часть переваренной пищи поступает в цитоплазму. У простейших, поглощающих
из окружающей среды жидкие органические вещества, пищеварительные вакуоли,
как правило, отсутствуют.
Так же для простейших характерны сократительные вакуоли – органоиды, основная функция которых заключается в регуляции осмотического давления в клетке. Эти вакуоли свойственны преимущественно пресноводным простейшим, так как в их теле осмотическое давление благодаря образованию солей, всегда выше, чем в окружающей среде, в результате чего через покровы внутрь тела непрерывно поступает вола. Удаление избытка воды с помощью сократительных вакуолей предохраняет простейших от гибели. Вместе с водой через сократительные вакуоли из тела простейших частично выводятся продукты обмена веществ.
Для клеток растений характерна хорошо развитая система вакуолей, в значительной степени обеспечивающая их соматические свойства.
Почти во всех клетках, особенно в старых, хорошо заметны полости – вакуоли. Они заполнены клеточным соком.
Клеточный сок – это вода с растворенными в ней сахарами и другими органическими веществами. В клеточном соке могут содержаться красящие вещества, придающие синюю, фиолетовую, малиновую окраску лепесткам и другим органам растений. Осенняя окраска листьев также обусловлена окрашенным клеточным соком. Разрезая спелый плод или другую сочную часть растения, мы повреждаем клетки, и из их вакуолей вытекает сок. Цитоплазма в каждой из клеток оттеснена вакуолью к оболочке. Молодые клетки, в отличии от старых, неспособных делиться, содержат много вакуолей. В старой клетке обычно имеется одна большая вакуоль.
II. ОСНОВНОЕ СОДЕРЖАНИЕ.
Растительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал.
Вакуоль (франц. vacuole, от лат. vacuus — пустой) – наполненный жидкостью мембранный мешочек. В животных клетках могут наблюдаться небольшие вакуоли, выполняющие фагоцитарную, пищеварительную, сократительную и другие функции. Растительные клетки имеют одну большую центральную вакуоль. Жидкость, заполняющая её, называется клеточным соком. Это концентрированный раствор сахаров, минеральных солей, органических кислот, пигментов и других веществ. Вакуоли накапливают воду, могут содержать красящие пигменты, защитные вещества (например, таннины), гидролитические ферменты, вызывающие автолиз клетки, отходы жизнедеятельности, запасные питательные вещества. У растений вакуоли — производные эндоплазматической сети, окружены полупроницаемой мембраной — тонопластом. Вся система вакуолей растительной клетки называется вакуомом, который в молодой клетке представлен системой канальцев и пузырьков; по мере роста и дифференцировки клетки они увеличиваются и сливаются в одну большую центральную вакуолю, занимающую 70- 95% объема зрелой клетки.
Вакуоли растений, строение и функции.
В
очень молодых клетках вакуолей нет или они почти незаметны по мере роста и дифференцировки клетки вакуоли появляются в разных её участках, а затем, постепенно увеличиваясь, сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 80% объема всей клетки.
Центральные вакуоли отделены от цитоплазмы одинарной липоидно-белковой полупроницаемой мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от эндоплазматической сети. Такие первичные провакуоли растут в объеме, сливаются друг с другом и, в конце концов, образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки.
Полость вакуоли заполнена так называемым клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Одной из главных ее функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны, тонопласта, так и плазмалеммы способствуют тому, что вакуоль функционирует в
качестве осмометра и придает клетке необходимую прочность и тургисцентность (напряженность).
Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от метаболизирующей гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через который может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. Так выводятся, секретируются из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например антоцианы. Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5).
Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких, как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях происходит запасание белков, что характерно для семян. Поступление белков в вакуоли связано со способностью вакуолей ЭР и АГ сливаться с тонопластом. Запасание белков семян происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов, протеазы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян. Стенки вакуолей могут образовывать впячивания внутрь, затем они отщепляются от тонопласта и растворяются внутри вакуоли.
Вакуолярная мембрана и роль вакуоли в растительной клетке.
Отличительной особенностью растительной клетки является наличие в ней центральной вакуоли, которая в ряде случаев может занимать до 90% объема зрелой растительной клетки и играет особую роль в структурной и функциональной организации растительного организма. Вакуоли активно участвуют во внутриклеточном перераспределении веществ, а также выполняют функции лизосом. Вакуоли аккумулируют большие количества запасных веществ и
других метаболитов, составляющих пищевую и лекарственную ценность растений (сахара, аминокислоты, органические кислоты, витамины, белки и др.).
При отложении веществ в запас вакуолярная мембрана выступает в качестве конечного барьера, регулирующего процессы накопления и удержания питательных веществ в запасающих органах растений. Механизмы формирования вакуолей при дифференциации клеток подробно рассматривались Матилем, который показал, что центральную вакуоль нужно рассматривать как скомбинированную органеллу, образованную путём объединения и слияния мембран различных клеточных органелл. Она является конечным продуктом дифференциации эндомембранной системы растений.
Образование центральной вакуоли стало решающем условием и основой выживания при выходе растений из воды на сушу. Формирование центральной вакуоли со специализированной мембраной — тонопластом, имеет фундаментальное значение в эволюции сосудистых растений и, видимо, является генетически закреплённым способом поддержания гомеостаза целого растения. То, что центральные вакуоли имеются практически во всех растительных организмах, свидетельствует о фундаментальной значимости этих органелл, поскольку в противном случае они бы элиминировали в процессе эволюции, по крайней мере, у некоторых видов или в некоторых тканях. Возникновение вакуоли способствовало ускорению эволюции растений и, в свою очередь, привело к изменению функциональной нагрузки и дифференциации вакуолей и их мембран в связи со специализацией клеток в системе целого растения.
В выполнении указанных функций ключевая роль принадлежит уникальной клеточной мембране — тонопласту. На сегодня известно, что вакуолярная мембрана содержит ряд специализированных систем пассивного и активного переноса веществ: каналы, переносчики, протонные помпы. Основной вклад в транспорт на тонопласте вносят две протонные помпы: Н^-АТФаза и ЕГ-пирофосфатаза. Оба фермента способны преобразовывать освобождённую при гидролизе АТФ и неорганического пирофосфата энергию в перенос протонов через тонопласт. Генерируемая разность электрохимического потенциала расходуется на вторичный транспорт ионов, Сахаров, аминокислот, который осуществляется через каналы и переносчики. Таким образом, Н^-АТФаза и К^-пирофосфтаза являются ключевыми ферментами в системе переноса углеводов и других соединений в антипорте с протоном.
Функции вакуоли в клетках высших растений.
В настоящее время убедительно доказано, что вакуоль является полифункциональным компартментом растительных клеток. Она играет основную роль в запасании метаболитов, регуляции клеточного объёма, тургора, а также pH и ионного гомеостаза цитозоля, что обеспечивает работу ферментов цитоплазмы. Вакуоль изолирует вторичные продукты метаболизма и метаболические яды, в ней происходят процессы детоксикации и катаболической деградации целого ряда соединений эндогенного и экзогенного происхождения, переваривание «стареющих» или повреждённых органелл.
В последние годы выявлена важная роль вакуоли в трансдукции сигналов различной природы, в процессах апоптоза и в ответных защитных реакциях растений на стрессовое воздействие. Это перечисление наиболее значимых функций вакуоли скорее всего не является исчерпывающим.
Трудно сказать, какая из этих функций является наиболее важной для растения. Одной из первых была сформулирована концепция о функции растительных вакуолей как о литическом компартменте, который, по аналогии с лизосомами животных клеток, переваривает «стареющие» или повреждённые органеллы. Кислые гидролазы были обнаружены в вакуолях, изолированных из всех изученных растительных объектов. Многие клеточные белки, имеющие дефект в своей структуре, способны накапливаться внутри вакуоли и там претерпевать протеолитическую деградацию под действием вакуолярных гидролаз. Интенсивная деградация белков внутри вакуоли свойственна также процессу вакуолярной аутофагии растительных клеток, который происходит в условиях сахарозного или углеводного голодания, когда содержание Сахаров и крахмала в клетках, а также их метаболическая активность заметно снижаются. Установлено, что вакуолярная аутофагия свойственна также стареющим клеткам растений. Из-за наличия внутриклеточных пулов углеводов и способности контролировать процесс аутофагии растительные клетки в условиях голодания могут некоторое время сохранять свою жизнеспособность. Литическая функция вакуолей тесно связана с апоптозом, который, как было показано в последние годы, является важным фактором развития и дифференциации многоклеточного растительного организма. В частности, он имеет место в клетках листьев как ответная, защитная реакция на действие патогена или во время старения таких клеток, способствуя транслокации тех или иных компонентов к другим, более молодым, растущим растительным клеткам.
Доминирующая роль центральной вакуоли в регуляции внутриклеточного осмотического давления и клеточного тургора доказана результатами многочисленных исследований. Она
находится в соответствии с интенсивным накоплением внутри вакуоли неорганических ионов, а также целого ряда других соединений, в том числе органических. Осмотический баланс в клетке может достигаться благодаря аккумуляции внутри вакуолярного и цитозольного компартментов осмолитов различной природы, что особенно важно в условиях осмотического стресса, когда в цитоплазме накапливаются осмолиты, способные защитить ферменты и другие биополимеры от инактивации в результате обезвоживания или сопутствующего ему окислительного стресса. Осморегуляторная функция вакуоли не ограничивается участием в регуляции клеточного тургора и, по-видимому, связана с включением в регуляцию объёма или оводнённости цитозоля растительных клеток. Это предположение подтверждается обнаружением на вакуолярной мембране аквапоринов -интегральных белков, облегчающих пассивный трансмембранный перенос молекул воды.
Одной из основных функций вакуоли является гомеостатическая регуляция ионного состава цитозоля растительных клеток, связанная с поддержанием оптимальных условий для активной работы ферментов цитоплазмы. Её потенциальная способность к выполнению этой важной функции в клетке обусловлена тем, что она может накапливать внутри себя значительные концентрации как основных физиологически важных ионов, таких как Н4″, К+, Na+, Ca2+, Mg2+, СГ, N03″, так и ионов, обладающих токсическим действием (Cd , Zn ). Недавно была убедительно продемонстрирована гомеостатическая регуляция уровня К+ в цитозоле растительных клеток за счёт его вакуолярной компартментации. Включение вакуоли в регуляцию уровня свободного Са2+ также является хорошо известным примером участия этой органеллы во внутриклеточном ионном гомеостазе растительных клеток. Регуляция уровня Са2+ в цитозоле с участием вакуоли включает в себя не только аккумуляцию Са2+ внутри этой органеллы, но и мобилизацию его из вакуоли в цитозоль. Экспорт Са из вакуоли выполняет также очень важную роль в трансдукции сигналов в растительной клетке. В этом случае вакуоль выступает в роли донора вторичных посредников, обусловливающих передачу сигналов к метаболическим системам клетки.
Одна из важных функций вакуоли состоит в изоляции вторичных продуктов метаболизма и метаболических ядов. К настоящему времени достигнут значительный прогресс в изучении обусловленных активностью вакуоли механизмов детоксикации экзогенных токсинов растительными клетками. Решающую роль в этом сыграло недавно обнаруженная способность изолированных вакуолей к Mg-АТФ-зависимой аккумуляции разнообразных глютатион-S-коньюгатов, в том числе и с гербицидами. Экспорт из цитозоля в вакуоль соответствующих GS-коньюгатов предотвращает токсическое действие на клетку тех или иных ксенобиотиков. В настоящее время найдено, что через GS-коньюгат-транслоцирующую систему могут
переноситься GS-коньюгаты антоцианина, некоторые фенольные соединения с аллелопатическими свойствами, а также некоторые фитоалексины. В животных клетках через эту транспортную систему могут изолироваться соединения, образуемые в ходе перекисного окисления липидов клеточных мембран или окислительного повреждения ДНК под действием активных форм кислорода. Поэтому есть основание предполагать, что эти же функции будут обнаружены и в растительных клетках.
Одной из наиболее важных функций вакуоли является запасающая. В некоторых случаях в вакуолях накапливаются значительные количества аминокислот и водорастворимых углеводов, причём их содержание нередко достигает 70-80% от суммы всех аминокислот или углеводов клетки. Нередко в вакуолях обнаруживают значительный пул органических кислот. Так, на листьях Bryophylum показано, что всего 2% всей внутриклеточной изолимонной кислоты участвует в метаболизме, остальные 98% изолированы в вакуолях. Состав углеводов, накапливаемых в большом количестве в вакуолях, значительно различается. Если вакуоли из лепестков и листьев тюльпана содержат в основном глюкозу и фруктозу, то корнеплоды столовой свёклы накапливают сахарозу. В вакуолях накапливаются большие количества фенолов, нитратов, витаминов и других вторичных соединений, которые нередко являются антиоксидантами или относятся к лекарственным соединениям.
В выполнении всех этих функций важная роль принадлежит уникальной клеточной мембране — тонопласту, которая разделяет содержимое вакуоли и цитоплазму клетки.
1.2. Тонопласт: особенности химического состава и физиологическая роль белков
Изолированная вакуолярная мембрана обладает типичным трёхслойным строением при толщине 9,5-10 нм, присущим для пограничных мембран in suti, и характеризуется достаточно сложной надмолекулярной структурой. При этом тонопласт имеет чёткую асимметрию, которая выражается в различной электронной плотности периферических слоев мембраны на поперечных срезах и в неодинаковой концентрации внутримембранных глобулярных частиц на вакуолярных и цитоплазматических сколах. Коэффициент распределения глобулярных частиц между вакуолярной и цитоплазматической сторонами составлял 725/1006 = 0.361. Известно, что высокая насыщенность глобулярными частицами (до 1000 — 3500 частиц на 1мкм2) характерна для мембран, проявляющих особенно высокую функциональную активность.
Внутримембранные глобулы, как правило, представляют собой глобулы трансмембранных (интегральных) белков, которые могут являться ферментами, ионными каналами и переносчиками. Суммарная площадь, занятая частицами на продольных сколах, составляла около 23,5% от общей площади поверхности тонопласта, а объём, занимаемый глобулярными частицами, составлял около 22% от объёма мембраны. Это удовлетворительно
коррелирует с результатами прямых биохимических анализов тонопласта, согласно которым во фракции вакуолярных мембран, тщательно очищенных от периферических белков, около 20% приходится на долю интегральных белков, извлекаемых из мембран только с помощью детергентов. Белок / липидное соотношение в тонопласте столовой свёклы составляло 0,69. 6-8% белков тонопласта были отнесены к периферическим, остальные 92% -к интегральным, пронизывающим мембранный матрикс. При электрофоретическом разделении в ПААГ с ДС в спектре белков тонопласта выявлялось около 20 полипетидов, большинство из которых имели молекулярные массы менее 70 кД. Спектр белков тонопласта, солюбилизированных Тритоном Х-100, состоял из 15 белковых полос, 8 из которых были гликопротеинами.
Такие биохимические характеристики, как белок/липидное соотношение, количественный и качественный спектр белков изменялись при использовании других объектов для получения вакуолярных мембран. Так, например, во фракции тонопласта, выделенной из корнеплодов столовой свёклы и из лепестков амариллиса обнаружены лишь следы углеводов, тогда как в тонопласте, выделенном из дрожжей, на 100 мг белка приходилось 79 мг углеводов. Но все исследователи, независимо от объекта исследования, отмечают высокую текучесть и эластичность вакуолярной мембраны, которая обусловлена большим содержанием липидов, составляющих до 80% от её общего веса.
III. ЛИТЕРАТУРА.
1. К.М.Суханова «Цитология» Москва,1970г.
2. Л.Я.Кулинич «Справочник по биологии» Москва,1986г.
3. Б.М.Медников «Аксиомы биологии» Москва,1985г.
4. В. Азерников. Тайнопись жизни. Москва, 1973г.
5. Н. Н. Воронцов, Л. Н. Сухорукова. Эволюция органического мира. Москва,1991г.
6. Э. Рис, М. Стернберг. От клеток к атомам. Москва, 1988г.
7. А. С. Трошин, А. Д. Браун, Ю. Б. Вахтин, Л. Н. Жинкин, С. Штрбанова. Кто мы? Книга о жизни, клетках и ученых. Москва, 1984г
8. Руководство по цитологии, т. 1, М.—Л., 1965, гл. 2;
9. Робертис Э. де, Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967;
10. Робертсон Дж., Мембрана живой клетки, в сборнике: Структура и функция клетки, пер. с англ., М., 1964; Finean J. В., The molecular organization of cell membranes, «Progress in Biophysics and Molecular Biology», 1966, v. 16, p. 143—70.
11. http://www.zubreshka.ru/?id=27090&from=350
12. http://www.college.ru/biology/course/content/chapter9/section1/paragraph5/theory.html
13. http://www.medkurs.ru/lecture1k/med_biology/qm24/2480.html
14. http://all-referats.ru/alls/10/6497.html
15. http://fundamed.narod.ru/Shpori/Cytology_3.doc
Источник: infourok.ru