Целлюлоза придает прочность клеточной стенке клеток
КЛЕТОЧНАЯ СТЕНКА, клеточная оболочка одноклеточных, колониальных и многоклеточных грибов и растений. У многоклеточных растений все клеточные стенки и межклетники образуют единую общеорганизменную структуру (апопласт), обеспечивающую передвижение веществ от клетки к клетке. Толщина клеточной стенки варьирует у разных клеток (у растительных, например, от 0,1 мкм до многих десятков мкм). Формирующие её вещества представлены преимущественно полисахаридами. Они образуют матрикс (аморфное вещество) и включённые в него тонкие нити разного размера – микро и макрофибриллы, которые практически нерастяжимы и обеспечивают прочность клеточной стенки на разрыв. Такие микро и макрофибриллы сложены параллельными линейными неразветвлёнными молекулами хитина (у большинства грибов), целлюлозы (у некоторых грибов и большинства растений), ксиланов (у ряда зелёных и красных водорослей), маннанов (у некоторых зелёных водорослей). На их долю приходится от «4% до «90% сухой массы клеточной стенки, что зависит от таксономической принадлежности организма, типа ткани и возраста клетки. Основу матрикса составляют разветвлённые и неразветвлённые молекулы пептидогликанов, маннанов (грибы), пектинов и гемицеллюлоз (высшие растения), альгинатов и фукоидана (бурые водоросли), которые соединены различными связями в ажурную трёхмерную сеть, проницаемую для водных растворов. Матрикс включает также белки (в том числе гликопротеины), роль которых в клеточных стенках плохо изучена.
Образование клеточной стенки начинается с отложения на поверхности плазматической мембраны слоя матрикса, биополимеры которого синтезируются внутри клетки и путём экзоцитоза выделяются на её поверхность. Синтез микро и макрофибрилл происходит из выделяемых протопластом мономерных предшественников с помощью подвижных белковых комплексов. Последние пронизывают насквозь мембрану и связаны с подстилающими её микротрубочками цитоскелета. У одноклеточных и колониальных организмов изначальный слой матрикса лишён микро и макрофибрилл, часто ослизняется; у спор он инкрустируется минеральными веществами и/или полифенолами либо замещается так называемым спорополленином; у многоклеточных организмов изначальный матрикс становится межклеточной пластинкой. В дальнейшем протопласт формирует собственно клеточные стенки, матрикс которой является непосредственным продолжением изначального слоя. У грибов в толще клеточной стенки образуется срединный слой из пептидогликанов без фибрилл. У растений различают первичную и вторичную клеточные стенки.
рвая образуется во время роста клетки и завершается вскоре после его окончания; содержит небольшое число беспорядочно ориентированных микро и макрофибрилл, что позволяет стенке растягиваться по мере клеточного роста. Клетки некоторых типов высших растений на первичной клеточной стенке формируют изнутри вторичную клеточную стенку с микро и макрофибриллами, располагающимися плотно, параллельно друг другу, с хорошо выраженной слоистой структурой. Фибриллы различных слоёв вторичной клеточной стенки ориентированы в разных направлениях, вследствие чего она становится нерастяжимой. Клеточные стенки могут подвергаться обызвествлению (например, у «известковых» зелёных и красных водорослей), окремнению (например, у хвощей), одревеснению (у сосудистых растений), опробковению (у высших растений), замещению пектиново-гемицеллюлозного матрикса каллозой (в ситовидных полях элементов флоэмы).
Клеточная стенка выполняет функции внешнего скелета клетки, защитной оболочки, регулятора водного режима, участвует в обменных процессах, передаче сигналов внутрь клетки из окружающей среды. Гемицеллюлозы клеточной стенки в эндосперме семян являются запасным веществом для зародыша (хурма, пальмы). Некоторые клетки высших растений в зрелом состоянии лишаются протопласта и представлены исключительно клеточной стенкой (например, клетки феллемы, проводящие элементы ксилемы); в основном из клеточной стенки отмерших клеток состоит древесина. Фактически именно клеточные стенки служат сырьём для деревообрабатывающей и целлюлозно-бумажной промышленности, производства вискозы, а также получения агара и альгинатов для медицинской, пищевой и косметической промышленности. О клеточной стенке бактерий смотри в статьях Бактерии, Грамотрицательные и грамположительные бактерии.
Лит.: Ботаника с основами фитоценологии: анатомия и морфология растений. М., 2007.
А. К. Тимонин.
Источник: knowledge.su
Клеточные стенки высших растений
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жесткость клетки для структурной и механической поддержки, придают форму клетке, направление ее роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургора, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают против патогенов, проникающих из окружающей среды, и запасают углеводы для растения. Растительные клеточные стенки строятся прежде всего с углеводного полимера целлюлозы.
В течение цитокинеза сначала с клеточной пластинки формируется промежуточная ламела, и первичная клеточная стенка расширяется внутри промежуточной ламели. Фактическая структура клеточной стенки не всегда четко определена, существуют несколько моделей ее структуры: модель перекрестных ковелентних связей, модель привязи, модель диффузного слоя и модель стратифицированного слоя. Однако, первичная клеточная стенка может быть определена как сеть волокон целлюлозы направленных в случайных направлениях.
локна воздержатся вместе водородными связями, обеспечивает достаточно высокую прочность. Клетки держатся вместе и разделяют желатиновую мембрану, называет его промежуточная ламела, которая содержит пектаты (соли пектиновой кислоты) магния и кальция. Клетки взаимодействуют через плазмодесмы, то есть цитоплазмени канальцы, соединяющие цитоплазмы клеток через клеточные стенки.
В некоторых типах клеток некоторых растений, после того как достигается максимальный размер или определенная точка развития, между растительным клеткой и первичной стенкой формируется вторичная стенка. В отличие от первичной стенки, волокна выравниваются в основном в одном направлении, и с каждым дополнительным слоем их ориентация слегка меняется. Клетки с вторичными клеточными стенками очень жесткие. Межклеточное взаимодействие все еще возможна через отверстия во вторичной клеточной стенке, позволяющие плазмодкрми проникать через обе клеточные стенки.
Основные углеводы, составляющих первичную клеточную стенку, это целлюлоза, гемицеллюлоза и пектин. Целлюлозные микроволокна связываются через хемицелюлозни мостики, формируя целлюлозно-хемицелюлозну сеть, которая окружена матрицей пектина. Самый общий тип хемицелюлозы в первичной клеточной стенке — ксилоглюкан.
Растительные клеточные стенки также содержат ряд белков, распространенные включают богатые гидропролин гликопротеины (HRGP), также известные как екстенсины, арабиногалактан протеины (AGP), богатые глицин протеины (GRP) и богатые пролин протеины (PRP).
исключением богатых глицин протеинов, все эти белки гликозилированные и содержат гидроксипролин (Hyp). Каждый класс гликопротеинов определяется характерной, часто повторяющейся последовательности аминокислот. Причудливые протеины содержат два или более областей, каждая из последовательностью от другого класса гликопротеинов. Большинство белков клеточной стенки перекрестных связаны с углеводами клеточной стенки и, возможно, имеют структурные функции.
Вторичные клеточные стенки могут содержать лигнин и суберин, делая стенки твердыми. Относительный состав углеводов, дополнительных компонентов и белков зависит от вида растения, типа и возраста клеток.
Клеточные стенки водорослей
Подобно высших растений, большинство водорослей, независимо от происхождения, имеют клеточные стенки. Клеточные стенки водорослей содержат целлюлозу и многие другие гликопротеинов. Дополнительные полисахариды в клеточных стенках водорослей раньше часто использовали как один из признаков для установления их таксономии, но некоторые несвязанные группы имеют те же полисахариды.
- Манозилови микроволокна зучтричаються в клеточных стенках многих зеленых водорослей (примитивних растений), в том числе родов Codium, Dasycladus и Acetabularia, и некоторых красных водорослей например, родов Porphyra и Bangia.
- Ксилан
- Альгиновая кислота — обычный полисахарид в клеточных стенках бурых водорослей
- Сульфоновани ролисахариды встречаются клеточных стенках большинства водорослей, например, в красных водорослях находим агарозу, каррагинан, порфиран, фурселеран и фуноран.
Другие вещества, которые могут быть найдены в водорослевых клеточных стенках, включают спорополенин и кальций.
Группа диатомовых водорослей (диатомей) синтезируют клеточные стенки (также известно как фрустулы или клапаны) с Кремнев кислоты (особенно ортокремневои кислоты, H 4 SiO 4). Кислота полимеризуется внутриклеточно, затем стенка вытесняется наружу для защиты клетки. Такие стенки требуют меньше энергии на свой синтез, позволяя сохранить много энергии, что, возможно, частично отвечает за быстрый рост этих организмов.
Клеточные стенки грибов
Не все виды грибов имеют клеточные стенки, но если стенка присутствует, она состоит из глюкозамина и хитина, того же углерода, что придает прочность экзоскелета насекомых. Они служат подобной цели, что и клеточные стенки растений, предоставляя грибным клеткам твердость и поддерживать свою форму и предотвращения осмотическому лизису. Они также ограничивают вход молекул, потенциально ядовитых для гриба, например некоторым фунгициды растительного и искусственного происхождения. Состав, свойства и форма грибной клеточной стенки меняются на протяжении жизненного цикла и зависят от условий произрастания.
Другие эукариоты
Оомицеты, патогены растений похожи на грибы, также имеют целлюлозные клеточные стенки. До недавнего времени за этот признак их относили к грибам, структурные и молекулярные свидетельства заставили отнести их к гетероконтив, как и бурые водоросли.
Клеточные стенки бактерий
Как в других организмах, бактериальные клеточные стенки обеспечивают структурную целостность клеткам. У прокариот, первичная функция клеточной стенки — защитить ячейку от внутреннего тургора, вызванным высокой концентрацией белков и инших молекулы внутри клетки по сравнению с окружающей средой. Бактериальная клеточная стенка отличается от других тем, что ее основным компонентом является пептидогликан, слой которого размещается немедленно вне цитоплазматической мембраны. Пептидогликан отвечает за твердость стенки и придания формы клетке. Он относительно пористый и не мешает потока растворимых молекул сквозь него. Существует два основных типа бактериальных клеточных стенок, по этому признаку бактерии делятся на грам-отрицательные и грам-положительные.
Грам-положительные бактерии
Грам-положительные бактерии характеризуется присутствием очень толстого слоя пептидогликана, который отвечает за содержание красителя кристал-виолет течение процедуры окрашивания по Граму. Такая стенка находится исключительно в организмах, принадлежащих к типам Actinobacteria (или грамм-положительные организмы с высоким содержанием G + C) и Firmicutes (или грамм-положительные организмы с низким содержанием G + C).
ктерии в пределах типа Deinococcus-Thermus также могут положительно краситься по Граму, но содержат некоторые структуры клеточной стенки, типичные от грамотрицательных организмов. В клеточную стенку грамположительных бактерий встроенные полимерного спирты, тейхоевая кислота, некоторые из которых связываются с липидами, формируя липотейхоеву кислоту. Ци вещества отвечают за соединение пептидогликана с цитоплазматической мембраной. Тейхоевая кислота придает клетке отрицательный электрический заряд благодаря наличию фосфодиестерних связей между мономерами тейхоевая кислоты.
Грамотрицательные бактерии
В отличие от грамположительных бактерий, грамотрицательные бактерии содержат очень тонкий слой пептидогликана, отвечающий за неспособность клеточных стенок содержать краситель кристал-виолет течение процедуры окрашивания по Граму. В дополнение к слою пептидогликанов, грамотрицательные бактерии имеют вторую, так называемую внешнюю мембрану, находится кнаружи от клеточной стенки и компонует фосфолипиды и липополисахариды на своей внешней стороне. Отрицательно заряженные липополисахариды также предоставляют клетке отрицательный электрический заряд. Химическая структура липополисахаридив внешней мембраны часто уникальная для отдельных штаммов бактерий и часто отвечает за реакцию антигенов с представителями этих штаммов.
Как любой двойной слой фосфолипидов, внешняя мембрана достаточно непроницаема для всех заряженных молекул. Однако, белковые каналы (погрузится) присутствуют во внешней мембране, позволяют пассивный транспорт многих ионов, сахара и аминокислот через внешнюю мембрану. Таким образом, эти молекулы присутствуют в периплазматическое, слое между внешней и цитоплазматической мембранами. Периплазматическое содержит слой пептидогликана и много белков, шо отвечают за гидролиз и прием внеклеточных сигналов. Читается, что перивлазма гелеобразная, а не жидкая, из-за высокого содержания белка и пептидогликана. Сигналы и живильни вещества с периплазматическое попадают в цитоплазму клетки используя транспортные белки в цитоплизматичний мембране.
Клеточные стенки архей
Хотя они и не уникальны, клеточные стенки архей несколько отличаются от бактериальных. Например, клеточные стенки большинства архей образованные внешними слоями белков или S-слоем. S-слоя распространенные в бактериях, где они служат единственным компонентом клеточной стенки в некоторых организмах (например в Planctomyces) или внешним слоем во многих организмах с пептидогликана. За исключением одной группы метаногенов, археи не имеют пептидогликаннои стенки. Даже в данном случае, пептидогликаны очень отличается от типа, найденного в бактериях.
Источник: info-farm.ru
Состав клеточной стенки растений
У растений стенка дочерних клеток образуется уже во время деления родительской. Впоследствии она называется первичной. У многих клеток позже образуется вторичная оболочка.
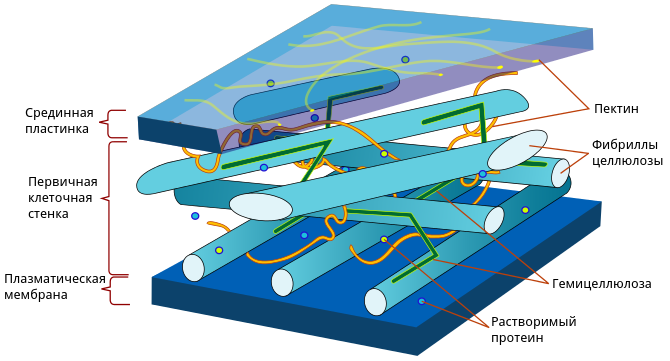
Первичная клеточная оболочка состоит из микрофибрилл целлюлозы, погруженных в матрикс из других полисахаридов. Отличительной особенностью волокон целлюлозы является их прочность. Молекула целлюлозы представляет собой длинную полисахаридную цепь. Отдельные молекулы соединяются друг с другом водородными связями в пучок, который называется микрофибриллой. Такие фибриллы образуют каркас клеточной стенки.
Матрикс клеточной стенки составляют полисахариды пектины и гемицеллюлозы, а также ряд других веществ (например, белков). Пектиновые вещества представляют собой группу кислых полисахаридов, их молекулы могут быть не только линейными, но и разветвленными. Гемицеллюлозы также смешанная группа полисахаридов. Длина их линейных молекул короче, чем у целлюлозы.
Оболочки соседних клеток растений соединены между собой срединной пластинкой, состоящих из пектатов магния и кальция, для которых характерна клейкость.

В состав стенок растений входит вода (составляет более половины массы), обуславливая ряд физических и химических свойств полисахаридов.
Жесткий каркас растения во многих местах пронизан каналами (плазмодесмами), по которым цитоплазма одной клетки соединяется с цитоплазмой соседних.
Клетки мезофилла листа (а также некоторые другие) на протяжении всей своей жизни имеют только первичную стенку. У большинства же клеток на первичную оболочку с внутренней стороны отлагается вторичная стенка, составленная из дополнительных слоев целлюлозы. Обычно в это время клетка уже дифференцирована и не растет (исключение составляют, например, клетки колленхимы).
В каждом отдельном слое вторичного утолщения микрофибриллы целлюлозы располагаются под одним углом (параллельно друг другу). Однако разные слои имеют разный угол, что обеспечивает большую прочность.
Часть клеток растений одревесневают (трахеальные элементы ксилемы, склеренхима и др.). В основе этого процесса лежит интенсивная лигнификация стенок (в небольших количествах лигнин есть во всех оболочках). Лигнин не является полисахаридом, а представляет собой сложное полимерное вещество. Отложения лигнина могут иметь различную форму (сплошную, кольцевую, спиральную, сетчатую). Он скрепляет целлюлозу, не дает ей смещаться. Лигнин не только обеспечивает прочность, но и дает дополнительную защиту от неблагоприятных физических и химических факторов.
Функции клеточной стенки
Оболочки разных клеток совместно обеспечивают всему растению и его отдельным частям механическую прочность и опору. Это функция клеточной стенки аналогична одной из функций скелета животных. Однако она не единственная.
Жесткость стенок препятствует растяжению клеток и их разрыву. В результате по физическим законам в клетки может путем осмоса поступать вода. Для травянистых растений тургоцентричность клеток является единственной их опорой.
Микрофибриллы целлюлозы ограничивают рост клеток и определяют их форму. Если микрофибриллы окольцовывают клетку, то она будет расти в длину (поперек направления волокон).
Связанные клеточные стенки образуют апопласт, по которому передвигается вода и минеральные вещества. Плазмодесмы связывают содержимое разных клеток в единую систему — симпласт.
Стенки сосудов ксилемы, трахеид, ситовидных трубок выполняют транспортную функцию.
Наружные клеточные стенки эпидермальных клеток покрыты воском (кутикулой). С одной стороны, он препятствует испарению воды, с другой – проникновению вредных микроорганизмов.
У некоторых растений в определенных клетках оболочки видоизменяются и служат местом запаса питательных веществ.
Источник: biology.su
Клеточная стенка (оболочка) является неотъемлемым компонентом клеток растений и грибов и представляет собой продукт их жизнедеятельности. Она придает клеткам механическую прочность, защищает их содержимое от повреждений и избыточной потери воды, поддерживает форму клеток и их размер, а также препятствует разрыву клеток в гипотонической среде. Клеточная стенка участвует в поглощении и обмене различных ионов, т. е. является ионообменником. Через клеточную оболочку осуществляется транспорт веществ.
Клеточная стенка, формирующаяся во время деления клеток и их роста путем растяжения, называется первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои, и образуется прочная вторичная клеточная оболочка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений и хитин у грибов), компоненты матрикса (гемицеллюлоза, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности оболочки (кутин и воск).
Молекулы целлюлозы за счет водородных связей объединяются в пучки —микрофибриллы. Переплетенные микрофибриллы составляют каркас клеточной оболочки. У большинства грибов микрофибриллы клеточной стенки состоят из хитина.
Микрофибриллы погружены в матрикс клеточной стенки. Матрикс состоит из смеси различных химических веществ, среди которых преобладают полисахариды (гемицеллюлозы и пектиновые вещества).
Гемицеллюлозы — это группа полисахаридов (полимеры пен-тоз и гексоз — ксилозы, галактозы, маннозы, глюкозы и др.). Молекулы гемицеллюлоз, как и целлюлозы, имеют форму цепи, но в отличие от последней их цепи короче, менее упорядочены и сильно разветвлены. Они легче растворяются и разрушаются ферментами.
Пектиновые вещества — это полимеры, построенные из моносахаридов (арабинозы и галактозы), галактуроновой кислоты (сахарной кислоты) и метилового спирта. Длинные молекулы пектиновых веществ могут быть линейны ми или разветвленными. Молекулы пектиновых веществ содержат большое количество карбоксильных групп и поэтому способны соединяться с ионами Mg2+ и Са2-. При этом образуются клейкие, студнеобразные пектаты магния и кальция, из которых затем складываются срединные пластинки, скрепляющие стенки двух соседних клеток.
Ионы двухвалентных металлов могут обмениваться на другие катионы (Н—, К+ и т. д.). Это обусловливает катионообменную способность клеточных оболочек.
Пектиновыми веществами и пектатами богаты оболочки клеток многих плодов. Так как при их извлечении из оболочек и добавлении сахара образуются гели, пектины используют как желе-образующие вещества для изготовления мармелада и др.
Помимо углеводных компонентов, в состав матрикса клеточной стенки входит структурный белок экстенеин —гликонроте-ин, который по своему составу близок к межклеточным белкам животных —коллагенам.
На долю матрикса приходится до 60% сухого вещества клеточной оболочки. Матрикс оболочки не просто заполняет промежутки между микрофибриллами, а образует прочные химические (водородные и ковалентные) связи между макромолекулами и микрофибриллами, что обеспечивает прочность клеточной стенки, ее эластичность и пластичность.
Основным инкрустирующим веществом оболочки клеток растений является лигнин — полимер с неразветвленной молекулой, состоящей из ароматических спиртов.
Интенсивная лигнификация (пропитка слоев целлюлозы лигнином) клеточных оболочек начинается после прекращения роста клетки. Лигнин может откладываться отдельными участками — в виде колец, спиралей или сетки, как это наблюдается в оболочках клеток проводящей ткани — ксилемы, или сплошным слоем, за исключением тех мест, где осуществляются контакты между соседними клетками в виде плазмодесм.
Лигнин скрепляет целлюлозные волокна и действует как очень твердый и жесткий каркас, усиливающий прочность клеточных стенок на растяжение и сжатие. Он же обеспечивает клеткам дополнительную защиту от физических и химических воздействий, снижает водопроницаемость. Содержание лигнина в оболочке достигает 30%. Инкрустация им клеточных оболочек приводит к их одревеснению, которое часто влечет за собой отмирание живого содержимого клетки.
Лигнин в сочетании с целлюлозой придает особые свойства древесине, которые делают ее незаменимым строительным материалом.
На клеточную оболочку могут откладываться также жиропо-добные вещества — суберин, кутин и воск.
Суберин откладывается на оболочку изнутри и делает ее практически непроницаемой для воды и растворов. В результате протопласт клетки отмирает и клетка заполняется воздухом. Такой процесс называется опробковением. Наблюдается опробковение оболочки клеток в покровных тканях многолетних древесных растений — перидерме, корке, а также в эндодерме корня.
Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками. Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами (целлюлозой и пектином) и образует кутикулу. Воск часто откладывается в кристаллической форме на поверхности частей растений (листьев, плодов), образуя восковой налет.
Кутикула и восковой налет защищают клетки от повреждений и проникновения инфекции, уменьшают испарение воды с поверхности органов.
В оболочках эпидермальных клеток некоторых растений (злаков, осок и др.) накапливается большое количество минеральных веществ (минерализация), в первую очередь карбоната кальция и кремнезема. При минерализации листья и стебли растений становятся жесткими, твердыми и в меньшей степени поедаются животными.
Таким образом, клеточная стенка играет важную роль в жизни клеток растений и грибов и выполняет ряд специфических функций.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
Источник: sbio.info