Клеточный цикл митоз
Клетки не возникают каждый раз заново, они образуются только в результате деления материнских кле ток. Промежуток времени от появления клетки в результате деления до ее разделения или гибели называется жизненным циклом клетки.
У эукариотических клеток жизненный цикл делится на две основные стадии: интерфазу и митоз
Интерфаза — это промежуток времени в жизненном цикле, в который клетка не делится и нормально функционирует. Интерфаза делится на три периода: G1, S и G2 периоды.
G1период (пресинтетический, постмитотический) — это период роста и развития клетки, в который происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь об разовавшейся клетки.
В Sпериоде (синтетическом) происходит процесс репликации ДНК. Генетическая формула клетки после удвоения ДНК — 2n4c.
G2 период (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центриолей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо различимыми, целостность ядерной оболочки не нарушается.
Некоторые клетки со временем перестают делиться и погибают, что может быть связано с завершением выполнения определенных функций, как в случае клеток эпидермиса кожи и клеток крови, или с повреждением этих клеток факторами окружающей среды, в частности возбудителями болезней.
Источник: www.sites.google.com
Клеточным циклом (митотическим циклом) называют весь период существования клетки от ее появления в результате деления до элиминации вследствие либо деления, либо апоптоза.
Клеточный цикл подразделяется на четыре периода: пресинтетический G1, синтетический S, постсинтетический G2 и митоз M. Периоды G1, S и G2 составляют в совокупности интерфазу, во время которой клетка сохраняет оформленное ядро, тогда как при делении митозом оно временно исчезает. Эта схема также подразумевает, что синтез ДНК происходит в интерфазе в ограниченный период времени, причем между ним и митозом существуют паузы (gaps).
Во время пресинтетического периода клетка активно синтезирует РНК и белки, восстанавливает органоиды, утраченные при делении, накапливает молекулы и энергию, необходимые для репликации, репарирует повреждения ДНК. Преобладание анаболических процессов в G1-периоде вызывает увеличение объема цитоплазмы, благодаря чему наблюдается рост клетки. Продолжительность периода G1 варьирует в широких пределах. Так, например, у лимфоцитов тимуса она составляет около 3 час., в кишечном эпителии – 10 час., в печени – 48 час., в эпидермисе кожи – 64 час.. При неблагоприятных условиях продолжительность G1-периода возрастает, а при стимуляции клеток гормонами, факторами роста и митогенами – снижается.
В синтетическом периоде происходит синтез ДНК и белков хроматина, что приводит к удвоению хромосом и увеличению размеров клеточного ядра. Репликация ДНК осуществляется в геноме эукариот параллельно во многих участках – репликонах, которым соответствуют петлевые домены хроматина. Число репликонов зависит, прежде всего, от количества ДНК в ядре и поэтому варьирует в пределах от 1 тыс. у дрожжей до 40-60 тыс. у млекопитающих. Репликоны собраны в кластеры, которые содержат от 20 до 80 точек инициации. Все репликоны, принадлежащие данному кластеру, реплицируются одновременно.
В репликации ДНК эукариот принимает участие большое количество ферментов. Наиболее важные из них – ДНК-полимеразы, праймазы, лигазы, топоизомеразы, ДНК-азы и РНК-азы. Репликация ДНК происходит по полуконсервативному механизму, при этом в точке инициации возникают две репликативные вилки, которые двигаются в противоположных направлениях со скоростью 1-3 тыс. пар нуклеотидов в минуту. Репликация ДНК в клеточном ядре происходит закономерным образом, распространяясь от эухроматина к гетерохроматину. Позже всех реплицируется сателлитная ДНК, которая локализуется в районе первичных перетяжек хромосом – центромер.
Одновременно с репликацией ДНК в ядро поступают гистоны и другие белки хроматина, которые синтезируются рибосомами в цитоплазме. Они взаимодействуют со вновь синтезированной ДНК и воспроизводят многоуровневую структуру хроматина, существовавшую в ядре до начала репликации. К концу S-периода количество ДНК и гистонов в клеточном ядре увеличивается ровно в два раза. При этом однохроматидные хромосомы, которые остались от материнской клетки, превращаются в двухроматидные. Это можно наблюдать непосредственно, вызывая конденсацию хромосом в интерфазе осмотическим шоком.
Продолжительность S-периода составляет для лимфоцитов тимуса 6 час., клеток кишечного эпителия – 8 час., печени – 16 час. В течение S-периода происходит также удвоение центриолей.
В постсинтетическом периоде клетка начинает непосредственную подготовку к делению. При этом в ней синтезируются тубулины, которые необходимы для формирования микротрубочек веретена деления, а также белки, участвующие в конденсации хромосом и других процессах митоза. В G2-периоде может также происходить незначительный репаративный синтез ДНК. Для этого периода клеточного цикла характерен высокий уровень посттрансляционных модификации белков, в особенности фосфорилирования и ацетилирования. Основными субстратами для соответствующих ферментов являются белки цитоскелета и ядерного матрикса. В G2-периоде происходит также активация тканеспецифических генов. Продолжительность этого периода у разных клеток меняется мало, составляя в большинстве случаев 2-4 часа.
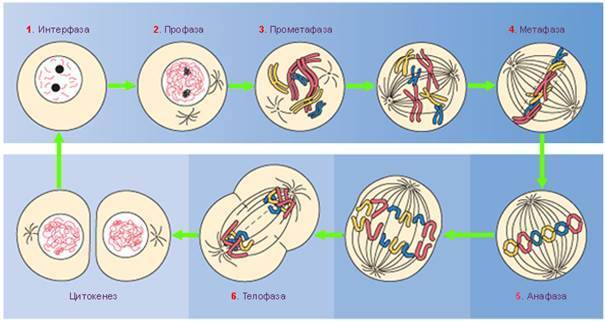
После окончания G2-периода клетка приступает к делению. Митотическое деление клетки состоит из четырех фаз – профазы, метафазы, анафазы и телофазы.
Переход из G2-периода в профазу митоза сопровождается повышением вязкости цитоплазмы и округлением клеток. Одновременно наблюдаются изменения в хроматине, мелкозернистая структура которого меняется на глыбчатую. В ранней профазе в ядре более отчетливыми становятся интенсивно окрашенные теломерные участки хромосом, которые располагаются по периметру ядра у нуклеолеммы. В поздней профазе видно, что от теломерных участков к центру ядра отходят плечи хромосом.
В профазе митоза происходит расхождение двух диплосом клеточного центра в противоположные концы – полюса клетки. Между ними формируется состоящее из микротрубочек веретено деления (митотическое,илиахроматиновое веретено). Число радиально расположенных вокруг диплосом микротрубочек увеличивается, и они образуют астросферу.
В конце профазы наступает диссоциация ядрышка, а содержимое цитоплазмы и ядра сливаются в миксоплазму. Одновременно происходит разрушение пластинчатого комплекса. Продолжительность профазы составляет от 15 до 60 мин.
В начале метафазы (прометафаза,илиметакинез) хромосомы перемещаются в плоскость экватора, которая располагается в середине веретена деления перпендикулярно линии, соединяющей полюса клетки. В результате образуется структура, которая называется метафазной пластинкой, или материнской звездой.
Митотические хромосомы в составе метафазной пластинки отличаются характерной морфологией. Обычно они имеют форму вытянутого цилиндра диаметром 1-2 мкм и длиной 1-50 мкм. В центре хромосомы находится первичная перетяжка, или центромера, которая делит ее на два плеча. В районе центромеры располагается прицентромерный гетерохроматин. Хромосомы с равными по длине плечами называются метацентрическими, с неравными плечами – субметацентрическими, а с очень коротким вторым плечом – акроцентрическими.
Некоторые хромосомы, кроме первичной перетяжки, имеют вторичную перетяжку. Она расположена ближе к концу хромосомы, отделяя от нее небольшой участок – спутник. В районе вторичной перетяжки локализован ядрышковый организатор, который содержит кластер генов рРНК и служит в интерфазном хроматине местом формирования ядрышка. Вторичная перетяжка окружена гетерохроматином.
Концы хромосом содержат особые участки – теломеры, которые состоят из гетерохроматина. Теломеры обеспечивают в интерфазном ядре прикрепление хромосом к нуклеолемме.
Между центромерой и теломерой с помощью методов дифференциального окрашивания в хромосомах выявляются различной ширины вставки из гетерохроматина — полоски, или бенды. Они структурируют хромосому на отдельные блоки, расположение которых видоспецифично.
Каждая митотическая хромосома состоит из двух идентичных сестринских хроматид, связанных между собой центромерой. У большинства клеток в организме хромосомы представлены двойным диплоидным набором в отличие от одинарного гаплоидного набора у гамет.
Метафазная пластинка представляет собой сложный пространственный комплекс, который состоит из центриолей, отходящих от них микротрубочек веретена деления и хромосом.
кротрубочки веретена присоединены к центромерам с помощью особых белковых структур – кинетохоров.В кинетохоре имеются три слоя – прилежащий к хромосоме плотный внутренний слой, рыхлый средний слой и плотный внешний слой. От внешнего слоя отходят многочисленные микротрубочки, образующие корону кинетохора.Белки плотного внешнего слоя CENP способны специфически связываться с сателлитной ДНК. Белки внутреннего слоя кинетохора обеспечивают соединение сестринских хроматид. В состав кинетохора входят также механохимические белки динеины, которые участвуют в перемещении хромосом по микротрубочкам. Продолжительность метафазы составляет 5-30 мин.
Переход клетки из метафазы в анафазупроисходит очень быстро. При этом сестринские хроматиды в хромосомах метафазной пластинки утрачивают связь между собой и начинают синхронное движение по направлению к противоположным полюсам клетки. Скорость движения хроматид, которые теперь превратились в самостоятельные хромосомы, может достигать 2 мкм/мин. В процессе движения ориентация хромосом меняется, и, поскольку центромеры опережают теломеры, они приобретают V-образную форму.
У животных анафаза слагается из двух процессов – движения хромосом к полюсам (анафаза А) и расхождения полюсов (анафаза В). У растений расхождения полюсов не наблюдается.
Движение хромосом в анафазе связано со скольжением микротрубочек веретена и моторными функциями динеина. Оно зависит от АТФ, кальция и температуры и может блокироваться колхицином и другими веществами, которые подавляют полимеризацию микротрубочек. Анафаза – самая короткая стадия митоза, продолжительность ее варьирует от 5 до 20 мин.
Телофазамитоза в общих чертах напоминает профазу, но события в ней происходят в обратном порядке. Она начинается с остановки хромосом, которые, не меняя ориентации, начинают деконденсироваться. Одновременно происходит восстановление нуклеолеммы из мембранных пузырьков, контактирующих с боковыми поверхностями хромосом. После полной реконструкции нуклеолеммы в ядре формируются новые ядрышки. Параллельно с восстановлением клеточного ядра происходит разборка веретена деления. Она начинается у полюсов клетки и заканчивается у экватора. Сохраняющиеся некоторое время в телофазе микротрубочки веретена могут формировать остаточные тельца.
Митоз завершается цитотомией,илицитокинезом — распределением цитоплазмы между двумя дочерними клетками. У животных цитотомия обеспечивается инвагинацией плазмолеммы между двумя ядрами и формированием перетяжки. У растений цитотомия происходит путем построения дополнительной внутриклеточной перегородки. Общая продолжительность митоза составляет обычно от 1 до 4 час.
После завершения митоза две дочерние клетки могут опять вступить в G1-период клеточного цикла. Однако такое поведение клеток характерно только для быстрорастущих тканей и культур клеток в экспоненциальной фазе роста. В остальных случаях часть клеток после митоза переходят из клеточного цикла в состояние пролиферативного покоя — G0. Остающиеся в цикле клетки составляют пролиферативный пул.
Величина пролиферативного пула определяется как отношение пролиферирующих клеток к общему числу клеток в популяции. Пролиферирующие клетки идентифицируются по включению радиоактивно меченых предшественников ДНК или с помощью моноклональных антител к ядерному антигену пролиферирующих клеток — PNCA. Величина пролиферативного пула отражает характер роста тканей и клеточных культур. В стабильных, не растущих и не обновляющихся клеточных популяциях, например, в мышечной и нервной тканях, величина пролиферативного пула составляет менее 1 %. В растущих или обновляющихся тканях величина пролиферативного пула варьирует в пределах от 10 до 30 %. Наиболее высокие значения пролиферативного пула характерны для быстро растущих опухолей — более 90 %.
Источник: studopedia.ru
В основе индивидуального развития всех организмов лежит клеточное деление. Время существования клетки от деления до деления называется клеточным (митотическим) циклом. Величина его может сильно различаться для разных организмов и для разных стадий развития. Типичный митотический цикл эукариотической клетки состоит из 4 периодов (рис. 3.3).
Пресинтетический период (G1) – наиболее длительный период клеточного цикла. Он характеризуется ростом клетки, накоплением РНК, АТФ, белков, необходимых для образования клеточных структур, подготовкой клетки к синтезу ДНК.
Рис. 3.3. Клеточный (митотический) цикл
Синтетический период (S) – период синтеза ДНК и репликации хромосом. В этот период происходит также интенсивный синтез гистонов, их перемещение в ядро, где они связываются с реплицированной ДНК. К концу периода каждая хромосома состоит из двух хроматид, имеющих идентичные копии молекулы ДНК. Таким образом, именно во время S-периода генетический материал клетки удваивается.
Постсинтетический период (G2) – период формирования структур, необходимых для процесса деления клетки. Продолжается синтез РНК и белков. Запасается энергия в виде АТФ.
Периоды G1, S, G2 иногда объединяют под названием интерфаза, однако надо заметить, что термин этот несколько устаревший, возникший в далекие времена, когда механизм клеточного деления был не изучен.
Период митоза (М) – период деления генетического материала и образования двух новых клеток. Этот период занимает менее 10 % времени клеточного цикла.
Последовательность периодов клеточного цикла можно представить следующим образом:
Митоз – основной способ деления эукариотической клетки. В нем выделяют 4 следующие друг за другом фазы:
1. Профаза. Идет процесс прогрессивной спирализации хромосом. Исчезают ядрышки, разрушается ядерная мембрана. Образуется веретено деления, состоящее из микротрубочек. К концу про-фазы центриоли клеточного центра расходятся к полюсам клетки.
2. Метафаза. Хромосомы выстраиваются в экваториальной плоскости. В области центромеры они прикреплены к нитям веретена деления, но некоторые нити веретена проходят от полюса до полюса, не прикрепляясь к хромосомам.
3. Анафаза. Центромера делится пополам, и хроматиды начинают синхронно расходиться к полюсам клетки. С этого момента они становятся самостоятельными дочерними хромосомами. Большой теоретический интерес представляет механизм распределения хромосом, случайность или предопределенность этого процесса. Не совсем понятна роль веретена деления и центриолей. В конце анафазы на полюсах клетки группируются два идентичных хромосомных набора.
4. Телофаза. Завершается обособление двух кариотипов. Вокруг них образуются ядерные мембраны. Происходит деспирализация хромосом, формируются ядрышки. Распадается митотическое веретено деления. Завершает телофазу процесс разделения цитоплазмы – цитокинез, в котором главную роль играют структуры цито-скелета.
Данная схема митоза характерна для всех высших эукариот. Некоторые протисты и грибы имеют ряд особенностей процесса, не затрагивающих его сущность.
Основное биологическое значение митоза заключается в точном распределении генетического материала между дочерними клетками.
Следующая глава >
Источник: bio.wikireading.ru
Клетка в своей жизни проходит разные состояния: фазу роста, фазу подготовки к делению и деление. Обычно делению клеток предшествует редупликация их хромосомного аппарата, синтез ДНК. Это правило является общим для прокариотических и эукариотических клеток. Время существования клетки от деления до деления или от деления до смерти, обычно называют клеточным циклом (cyclus cellularis). Во взрослом организме высших позвоночных клетки различных тканей и органов имеют неодинаковую способность к делению. Встречаются популяции клеток, полностью потерявшие свойство делиться. Это специализированные, дифференцированные клетки (зернистые лейкоциты крови). В организме есть постоянно обновляющиеся ткани — различные эпителии, кроветворные ткани. В таких тканях существует часть клеток, которые постоянно делятся, заменяя отработавшие или погибающие клеточные типы (клетки базального слоя покровного эпителия, клетки крипт кишечника, кроветворные клетки костного мозга). Многие клетки, не размножающиеся в обычных условиях, приобретают вновь это свойство при процессах репаративной регенерации органов и тканей. Размножающиеся клетки обладают разным количеством ДНК в зависимости от стадии клеточного цикла. Клеточный цикл – переход от деления к синтезу веществ, составляющих клетку, а затем опять к делению – можно представить на схеме в виде цикла, в котором выделяют несколько фаз.
После деления клетка вступает в фазу синтеза белков и роста, эту фазу называют G1. Часть клеток из этой фазы переходит в фазу G0, эти клетки функционируют и потом погибают без деления (эритроциты). Но большинство клеток, накопив необходимые вещества и восстановив свой размер, а иногда и без изменения размеров после предыдущего деления, начинают подготовку к следующему делению. Эта фаза называется фаза S – фаза синтеза ДНК, затем, когда хромосомы удвоились, клетка переходит в фазу G2 – фазу подготовки к митозу. Затем происходит митоз (деление клетки), и цикл повторяется заново. Фазы G1, G2, S вместе называются интерфазой(т.е. фазой между делениями клетки). Весь клеточный цикл состоит из 4 отрезков времени: собственно митоза (M), пресинтетического (G1), синтетического (S) и постсинтетического (G2) периодов интерфазы. Буква G- от английского Gap – интервал, промежуток. В G1-периоде, наступающим сразу после деления, клетки имеют диплоидное содержание ДНК на одно ядро (2c) (с – количество ДНК на клетку). В период G1 начинается рост клетокглавным образом за счет накопления клеточных белков, что определяется увеличением количества РНК на клетку. В этот период начинается подготовка клетки к синтезу ДНК (S-периоду).
В следующем, S-периоде происходит удвоение количества ДНК на ядро и соответственно удваивается число хромосом. В разных клетках, находящихся в S-периоде, можно обнаружить разные количества ДНК — от 2c до 4c. Это связано с тем, что исследованию подвергаются клетки на разных этапах синтеза ДНК (только приступившие к синтезу и уже завершившие его). S-период является узловым в клеточном цикле. Без прохождения синтеза ДНК неизвестно ни одного случая вступления клеток в митотическое деление. Постсинтетическая (G2) фаза еще называется премитотической. В данной фазе происходит синтез иРНК, необходимый для прохождения митоза. Несколько ранее этого синтезируется рРНК рибосом, определяющих деление клетки. Среди синтезирующихся в это время белков особое место занимают тубулины — белки микротрубочек митотического веретена.
Как известно, половые мужские и женские клетки несут единичный (гаплоидный,1n1c) набор хромосом и, следовательно, содержат в 2 раза меньше ДНК, чем все остальные клетки организма. Такие половые клетки (сперматозоиды и ооциты) с единичным набором хромосом называют гаплоидными. Плоидность обозначают буквой n. Количество ДНК на клетку (c) зависит от ее плоидности: клетки с 2n числом хромосом содержат 2c количества ДНК. При оплодотворении происходит слияние двух клеток, каждая из которых несет 1n набор хромосом, поэтому образуется исходная диплоидная (2n, 2c) клетка — зигота. В дальнейшем в результате деления диплоидной зиготы и последующего деления диплоидных клеток разовьется организм, клетки которого (кроме зрелых половых) будут диплоидными. Каждая хромосома является парной, а также каждая имеет две одинаковые части – левую и правую хроматиды. Такой набор хромосом характерен для клетки, уже начавшей свое деление, т.е. клетки, в которой прошел процесс удвоения ДНК. Удвоение количества ДНК называют синтетическим периодом, или S-периодом, клеточного цикла. Говорят, что количество хромосом в клетке остается прежним (2n), а число хроматид в каждой хромосоме – удвоенным (4c – 4 хроматиды на одну пару хромосом) — 2n4c. При делении в дочерние клетки от каждой хромосомы попадет одна хроматида и клетки получат полный диплоидный набор 2n2c.
При подготовке к делению происходит репликация ДНК, на каждой хромосоме синтезируется ее копия. После репликации ДНК конденсируется, хромосомы приобретают более компактную укладку, и в таком состоянии их можно увидеть в световом микроскопе. Между делениями эти хромосомы не столь конденсированы и в большей степени расплетены. Понятно, что в конденсированном состоянии им трудно функционировать. Хромосома имеет вид в виде буквы Х только во время одной из стадий митоза.
Среди разновидностей клеточного деления можно выделить: прямое бинарное деление, митоз, или непрямое деление (характерное для большинства клеток); амитоз, или прямое деление клеток; мейоз, или редукционное деление (при образовании половых клеток); эндорепродукция (образование полиплоидных и многоядерных клеток).
Бактериальные клетки содержат только одну кольцевую молекулу ДНК, прикрепленную к клеточной мембране. Перед делением клетки ДНК реплицируется и образуются две идентичные молекулы ДНК, каждая из которых также прикреплена к клеточной мембране. При делении клетки мембрана врастает между двумя молекулами ДНК так, что в конечном итоге в каждой дочерней клетке оказывается по одной идентичной молекуле ДНК. Такой процесс получил название прямого бинарного деления.
МИТОЗ Митоз (mitosis), кариокинез, или непрямое деление, — универсальный, широко распространенный способ деления клеток. При этом конденсированные и уже редуплицированные хромосомы переходят в компактную форму митотических хромосом, образуется веретено деления, участвующее в сегрегации и переносе хромосом (ахроматиновый митотический аппарат), происходит расхождение хромосом к противоположным полюсам клетки и деление тела клетки (цитокинез, цитотомия). Процесс непрямого деления клеток принято подразделять на несколько основных фаз: профаза, прометафаза, метафаза, анафаза, телофаза.
Источник: studopedia.info Клетки не возникают каждый раз заново, они образуются только в результате деления материнских кле ток. Промежуток времени от появления клетки в результате деления до ее разделения или гибели называется жизненным циклом клетки. У эукариотических клеток жизненный цикл делится на две основные стадии: интерфазу и митоз
Интерфаза — это промежуток времени в жизненном цикле, в который клетка не делится и нормально функционирует. Интерфаза делится на три периода: G1, S и G2 периоды. G1период (пресинтетический, постмитотический) — это период роста и развития клетки, в который происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь об разовавшейся клетки. В Sпериоде (синтетическом) происходит процесс репликации ДНК. Генетическая формула клетки после удвоения ДНК — 2n4c. G2 период (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центриолей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо различимыми, целостность ядерной оболочки не нарушается. Некоторые клетки со временем перестают делиться и погибают, что может быть связано с завершением выполнения определенных функций, как в случае клеток эпидермиса кожи и клеток крови, или с повреждением этих клеток факторами окружающей среды, в частности возбудителями болезней.
Источник: www.sites.google.com |
