Что происходит с атф в световую стадию
Фотосинтез
Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:
6СО2 + 6Н2О + Qсвета → С6Н12О6 + 6О2.
У высших растений органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты (строение хлоропластов — лекция №7). В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру, является гидрофильной и поэтому лежит на той поверхности мембраны, которая обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.

Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН—.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН— → •ОН + е—.
Радикалы •ОН объединяются, образуя воду и свободный кислород:
4НО• → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н+ заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е— + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез
Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать.
поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки. Углекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
Строение С4-растений: |
|
|
С4-фотосинтез: |
Значение фотосинтеза
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м2 поверхности в час.
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Источник: licey.net
Что такое световая фаза фотосинтеза
Фотосинтез – сложный процесс, состоящий из 2 фаз: световой и темновой. Реакции протекающие в световой фазе фотосинтеза могут проходить только при освещении. Для темновой фазы свет не важен, она может проходить в любое время. Ниже представлена схема световой и темновой фазы фотосинтеза.
В течение световой стадии, растение захватывает фотоны света с помощью специальных светособирающих комплексов. Энергия фотонов необходима для прохождения процесса распада воды на кислород и водород.
Этот процесс, который происходит в световую фазу фотосинтеза называется фотолизом воды. Далее происходит образование конечных продуктов световой фазы, которые необходимы для прохождения реакций темновой фазы. Световая энергия накапливается в виде АТФ – аденозинтрифосфата.
Водород после фотолиза воды соединяется с ферментом НАДФ (никотинамидадениндинуклеотидфосфат), образуя НАДФН, который является источником водорода для дальнейших химических реакций.
Кислород при фотолизе воды выделяется в атмосферу. Таким образом, результатом световой фазы фотосинтеза является: распад воды под воздействием световой энергии с образованием конечных продуктов АТФ и НАДФН, использующихся для синтеза органики и свободного кислорода.
Где происходит световая фаза
Все процессы происходящие в световой стадии и темновой фазе фотосинтеза протекают в специальных клеточных структурах, называемых хлоропластами. Хлоропласт – зеленая пластида, внутри которой содержится хлорофилл. В растительной клетке содержится большое количество хлоропластов, необходимых для прохождения химических реакций фотосинтеза.
Различные фазы процесса проходят в разных частях хлоропласта. Эта пластида имеет сложную структуру, в ее состав входит большое количество тилакоидов. Тилакоиды – особые структуры внутри хлоропласта, отвечающие за преобразование световой энергии. Тилакоиды, расположенные рядом, образуют стопки – граны.
Световая фаза фотосинтеза происходит в гранах тилакоидов, на их мембранах и во внутритилакоидном пространстве. В этом особенность световой фазы фотосинтеза и этим она отличается от темновой, во время которой химические реакции протекают в строме хлоропласта – плотном веществе между тилакоидами.
Фотохимическая суть процесса
Основным процессом световой фазы фотосинтеза является фотолиз воды, представленный следующим уравнением:
2H2O + Qсвета → 4H+ + 4e— + O2
Молекулы хлорофилла имеют свойство возбуждаться, и терять электроны при попадании на них кванта света. Эти электроны оседают на внешней стороне мембран, заряжая их отрицательно.
Сами молекулы хлорофилла, потерявшие электрон, восстанавливаются, отбирая электроны у воды, находящейся внутри тилакоида. При фотолизе вода расщепляется на следующие компоненты:
- Протоны водорода;
- Электроны водорода;
- Кислород.
Кислород является побочным продуктом фотосинтеза и не участвует в дальнейших процессах. Он выводится наружу из ткани растения и поступает в атмосферу. Протоны накапливаются в специальном протонном резервуаре, находящемся внутри тилакоида.
Протоны заряжают внутреннюю часть тилакоидной мембраны положительно. Таким образом, мембраны тилакоидов имеют отрицательный заряд с внешней стороны, а положительный – с внутренней. Эти заряды постепенно увеличиваются по мере накопления протонов и электронов.
Разность потенциалов между внешней и внутренней поверхностью мембраны должна достигать не менее 200mВ, чтобы начался процесс образования конечных продуктов световой стадии. Только в этом случае протоны начнут проходить сквозь каналы АТФ-синтазы, находящиеся в тилакоидных мембранах.
Для этого используется энергия протона, проходящего тилакоидную мембрану. Прошедший мембрану протон и электрон, находящийся на внешней ее стороне вступают в реакцию с находящейся в строме хлоропласта молекулой НАДФ с образованием НАДФН.
2Н+ + 2е— + НАДФ → НАДФН
Светособирающие комплексы
Только небольшая часть молекул хлорофилла поглощает энергию, отдавая электроны. Они находятся в реакционных центрах и называются молекулами-ловушками. Остальная же часть этого пигмента собрана в светособирающие комплексы, задачей которых является не поглощение, а передача энергии.
Для чего нужны светособирающие комплексы? Если бы каждая молекула хлорофилла улавливала свет, то такая работа была бы крайне неэффективной. Процесс возбуждения и потери электрона проходил бы очень редко, а структура передачи электронов была бы слишком сложной из-за очень большого количества молекул.
На самом деле существует очень мало молекул, поглощающих энергию и отдающих электроны. На каждую из них приходится до 300 молекул, собранных в светособирающие комплексы по антенному типу. Они расположены на нескольких уровнях.
Конечно, происходит передача не квантов света, а только энергии, полученной при поглощении света. Таким образом, хлорофилл может не только поглощать световую энергию, но и передавать ее.
На самом нижнем уровне светособирающего комплекса находится 1 молекула-ловушка. Энергия поступает к ней со всего антенного комплекса. Передача энергии происходит с определенными потерями ее количества.
Но молекула-ловушка получает энергию в десятки или даже в сотни раз чаще, чем молекулы, расположенные на самом высоком уровне. Молекулы-ловушки входят в состав фотосистем, которые участвуют в транспорте электронов во время световой стадии фотосинтеза.
Основные компоненты цепи переноса электронов
В течение световой стадии происходит перенос электронов от тилакоидных фотосистем с помощью промежуточных веществ-переносчиков до образования конечных продуктов фазы. Электрон-транспортная цепь имеет сложную структуру и множество компонентов.
Основными компонентами цепи переноса электронов являются:
Фотосистема 1 способна поглощать свет с длиной волны 700 нм, фотосистема 2 – 680 нм. Фотосистемы работают параллельно. При поглощении света фотосистемы отдают электроны на вещества-переносчики или акцепторы. В электрон-транспортной цепи задействовано множество акцепторов, которые захватывают электроны и отдают их другому компоненту цепи.
Когда фотосистема 2 теряет электрон под воздействием света, он сначала попадает на акцептор феофитин. Далее в его транспорте принимает участие целый ряд акцепторов, последним из которых является пластоцианин. Далее электрон попадает в фотосистему 1, восполняя электрон, отданный этой фотосистемой под воздействием кванта света.
Фотосистема 1 отдает электрон на акцептор ферредоксин. Отсюда он поступает в последний компонент цепи НАДФ-редуктазу. Здесь образуется в световую фазу фотосинтеза вещество НАДФН. Недостачу электронов фотосистема 1 восполняет за счет электронов, приходящих от фотосистемы 2.
Особое значение в электрон-транспортной цепи имеет комплекс цитохромов b6-f. Электроны, проходя через этот комплекс, многократно взаимодействуют с акцептором пластохиноном. При этом комплекс цитохромов увеличивает количество, не только электронов, но также и протонов, что повышает эффективность световой стадии.
Продукты световой стадии
При прохождении этапа световой фазы фотосинтеза образуются следующие продукты, необходимые для синтеза органики в дальнейших темновых реакциях: АТФ и НАДФН. АТФ – источник биохимической энергии. Эта молекула синтезируется из АДФ при поглощении энергии движущегося протона.
Формулу синтеза АТФ во время световой фазы фотосинтеза можно представить в следующем формуле:
АДФ + ортофосфорная кислота + энергия → АТФ + Н2О
Синтезированный АТФ может участвовать во всех химических реакциях, для прохождения которых необходима энергия. При взаимодействии с водой происходит обратная реакция с выделением энергии.
АТФ вновь расщепляется на АДФ и ортофосфорную кислоту:
АТФ + H2O → АДФ + ортофосфорная кислота + энергия
Для образования органических веществ при фотосинтезе такая энергетическая составляющая крайне необходима, так как синтез органики требует поглощения большого количества энергии. НАДФН – восстановленный фермент, который является источником водорода. Он используется в химических процессах темновой фазы, где отдает водород и превращается в фермент НАДФ.
Источник: florist-club.com
Световая фаза фотосинтеза: введение Форум Биологические базы данных Большая часть солнечной энергии испускается в виде фотонов — квантов видимого света. Энергия света поглощается хлорофиллом и переводит его в возбужденное состояние. Электрон в составе хлорофилла поглощает квант света определенной длины волны и перемещается на более высокий энергетический уровень этой молекулы. Такой электрон можно сравнить с камнем, поднятым на высоту, который приобретает потенциальную энергию и теряет ее, падая с высоты. Возбужденный электрон, как по ступеням, перемещается по цепи переносчиков электронов. При этом он теряет энергию, которая служит для фосфорилирования АДФ в АТФ и запасания ее в высокоэнергетической связи этого соединения. Часть возбужденных светом электронов используется также для восстановления НАДФ+ в НАДФН. Под действием солнечного света в хлоропластах происходит также расщепление молекулы воды — фотолиз, при этом образуются электроны, которые возмещают потери их хлорофиллом; в качестве побочного продукта при этом образуется кислород. В 1939г. Роберту Хиллу впервые удалось показать, что в изолированных хлоропластах под действием света, при добавлении акцептора электронов, происходит выделение кислорода. Эта реакция, описываемая приведенным ниже уравнением, получила название реакции Хилла : hv 2Н20+2А = 2АН2+02 где А — акцепторы электронов и протонов (или атомов водорода) ; такими акцепторами в хлоропластах являются молекулы НАДФ + ; hv — квант световой энергии (h — постоянная Планка, a v-частота излучения) . В составе хлоропластов описаны две фотосистемы разного строения и состава: фотосистема I и фотосистема II . На рис. 36 приведена схема, упрощенно изображающая просходящие в этих двух фотосистемах процессы. В фотосистеме I имеется реакционный центр, представляющий собой молекулы хлорофилла в комплексе с особым белком. Этот комплекс поглощает красный свет с длиной волны в 700 нм, поэтому его называют фотохимическим центром Р700 (буква Р от англ. pigment — пигмент) . Под действием света электрон в этом реакционном центре переходит в возбужденное состояние, "перескакивая" на высокий энергетический уровень молекулы хлорофилла. Возбужденный электрон хлорофилла переносится на ферредоксин, который представляет собой содержащий железо белок и является сильным биологическим восстановителем. От ферредоксина высокоэнергетические электроны через фермент — редуктазу ферредоксина переносятся на НАДФ + ,восстанавливая его в НАДФН. В молекулах хлорофилла фотосистемы I остаются при этом дыры — незаполненные места электронов, перешедших в НАДФН. Эти дыры заполняются электронами, которые образуются в фотосистеме II . В фотосистеме II также имеется реакционный центр — комплекс хлорофилла с белком, который поглощает свет с длиной волны 680 нм (его называют Р680 ). Под действием света электрон хлорофилла в фотохимическом центре Р680 также переходит в возбужденное состояние и захватывается первым переносчиком в длинной цепи, обозначаемым буквой Z. От этого акцептора электрон спускается по цепи переносчиков "вниз" и в конечном итоге заполняет дыру, образовавшуюся в хлорофилле фотосистемы I . Таким образом, фотосистема II поставляет электроны для фотосистемы I, которые расходуются в ней на восстановление НАДФ + в НАДФН. Однако это не единственная функция фотосистемы II. Возбужденные светом электроны фотосистемы II от акцептора Z "перескакивают" вниз по цепи переносчиков, куда входят пластохинон (PQ) , цитохром b 563, цитохром f, пластоцианин и конечный акцептор — хлорофилл фотосистемы I . На этом пути энергия электрона расходуется на "зарядку" универсального биологического аккумулятора: на фосфорилирование АДФ в богатую энергией АТФ . Таким образом, энергия света запасается в молекулах АТФ и расходуется далее для синтеза углеводов, белков, нуклеиновых кислот и для иных жизненных процессов растений, а через них и иных живых организмов, населяющих нашу планету.
Источник: touch.otvet.mail.ru
Комментарий специалиста
Конкурс «био/мол/текст», цитирую, «ежегодно собирает более сотни участников, отважившихся весело, но корректно рассказать о сложнейших проблемах современной биологии для широкого круга читателей». В представленных статье и видео о фотосинтезе первый пункт выполнен замечательно — рассказ ведется весело и задорно, однако научная часть нуждается в серьезной корректировке. Основные ошибки в представлениях о фотосинтезе кочуют по разным источникам, в том числе их, к сожалению, можно отыскать и в школьных учебниках по биологии за 11 класс.
Итак, попытаемся самые грубые ошибки исправить.
«Для световой фазы необходимо наличие солнечного света, а вот для темновой солнышко роли не играет. Поэтому темновая фаза может проходить и днем, и ночью».
Световая фаза фотосинтеза действительно зависит от света, а вот темновая — это скорее историческое название. Во-первых, темновая фаза фотосинтеза полностью зависит от наличия продуктов световой фазы, и, во-вторых, в темноте ключевые ферменты темновой фазы ингибируются, а в строме хлоропласта идут совсем другие процессы.
«Если света мало, фотосинтез идет одинаково при любой температуре».
Это все же художественное преувеличение — не при любой, но в некоторых температурных пределах, приемлемых для растительного организма. Так, при низкой интенсивности света фотосинтез будет идти одинаково при температурах 15 °С и 25 °С, однако при 5 °С интенсивность фотосинтеза значительно снизится из-за снижения скорости ферментативных реакций.
«Познакомимся с таким термином, как фотолиз. Фотолиз — это расщепление молекулы воды под действием фотонов. Если взять молекулу воды (H2O) и воздействовать на нее квантами света, то она распадется на катион водорода (H+) и анион гидроксида (OH−). Дело в том, что изначально молекула воды электронейтральна, но один из атомов водорода решает отдать свой электрон паре ОН. Электрон заряжен отрицательно, поэтому его потеря приводит к появлению “+” на водороде. А ОН, присоединив лишний электрон, оказывается с “−”».
Термин «фотолиз» абсолютно неприемлем для описания процессов, происходящих с водой во время световой фазы фотосинтеза. «Фотолиз» означает распад вещества непосредственно под действием света, однако с водой этого не происходит, иначе фотолиз шел бы у нас в каждом стакане воды… Вода — довольно устойчивое соединение, для ее расщепления на O2 и H2 необходимо либо действие электрического тока, либо очень высокие температуры (выше 1000 °С); ни то, ни другое не характерно для биологических систем. Поэтому термин «фотолиз» при рассказе о световой фазе фотосинтеза некорректен и не должен употребляться. Фотолиз, наверное, одно из самых распространенных заблуждений, связанных с фотосинтезом.
«Здесь содержится пигмент хлорофилл, который по своему составу очень похож на гемоглобин крови человека, но вместо атома железа содержит магний».
Гемоглобин — это белок, содержащий гем, а вот гем, в свою очередь, содержит центральный атом железа. Хлорофилл по своей структуре немного похож на гем, а не на гемоглобин.
«В это же время происходит фотолиз воды. Электрон движется к катиону водорода и молекуле аденозиндифосфата (АДФ). АДФ содержит два остатка фосфорной кислоты, а при встрече с электроном присоединяет еще один остаток и превращается в аденозинтрифосфат (АТФ). Молекула АТФ — это энергетическое депо клетки, в ней запасается энергия для всех процессов жизнедеятельности.
Кроме того, электрон присоединяется к катиону водорода и делает из него нейтральный атом, который затем переходит в строму.
Гидроксид-анион жертвует своим электроном, и тот занимает место ушедшего ранее. При этом образуется нейтральный ОН.
Четыре ОН-группы в ходе химической реакции дают две молекулы воды и кислород».
К сожалению, в описании световой фазы фотосинтеза в этой части статьи нет ни слова правды… Электрон не движется ни к протону, ни к молекуле АДФ. Атомарного водорода и «нейтрального ОН» не образуется. ОН-группы не вступают в реакцию, результатом которой будет образование кислорода и воды.
«Темновая фаза идет в строме хлоропласта. В ней участвуют атомы водорода, пришедшие из тилакоида, и молекулы углекислого газа».
И еще раз повторим, атомарного водорода при фотосинтезе не образуется. Во время работы световой фазы протоны (Н+) закачиваются во внутреннее пространство тилакоидов и выходят обратно, в строму, в результате работы АТФ-синтазы — фермента, синтезирующего АТФ.
«Сначала к CO2 присоединяется фермент рибулозобисфосфаткарбоксилаза»
Фермент рибулозобисфосфаткарбоксилаза захватывает молекулу СО2, но не присоединяется к ней, а наоборот присоединяет СО2 к молекуле пятиуглеродного сахара — рибулозо-1,5-бисфосфата.
«Для синтеза одной молекулы глюкозы необходимо 16 фотонов с длиной волны 680 нм».
Не очень понятно, откуда такая цифра и почему только фотоны с длиной волны 680 нм. По самым скромным подсчетам, на фиксацию одной молекулы СО2 необходимо затратить 8 квантов света, причем не обязательно с одинаковой длиной волны. Для синтеза одной молекулы глюкозы нужно 6 атомов углерода, то есть 6 молекул СО2, а значит, минимум 6×8=48 фотонов.
«Хлорофилл не единственный пигмент, содержащийся в растениях. Выделяют еще каротиноиды и фикобилины».
Здесь необходимо уточнить, что каротиноиды встречаются у всех фотосинтезирующих организмов, а вот фикобилины — только у некоторых водорослей и цианобактерий.
Внимательный читатель, возможно, подумает, что критиковать-то легко, а вот как же описать фотосинтез «на пальцах», без сложных терминов, чтобы было понятно неспециалистам, а еще лучше — старшеклассникам. Попробуем сделать это в рамках комментария.
Если сформулировать очень кратко, цель фотосинтеза — восстановить очень окисленное соединение СО2 до восстановленного соединения — сахара — с помощью энергии солнечных квантов и электронов от воды.
Действительно, традиционно фотосинтез делится на световую и темновую фазы, однако помним, что название «темновая» — историческое.
Световая фаза фотосинтеза происходит в мембране тилакоидов хлоропласта и полностью зависит от света, так как использует энергию фотонов. Основная задача световой фазы — обеспечить энергией (АТФ) и восстановителем (источником электронов) темновую фазу. Как это происходит?
Квант света (он же фотон) переводит молекулу хлорофилла в возбужденное состояние: это значит, что за счет энергии кванта повышается энергия одного из электронов молекулы хлорофилла, и этот возбужденный электрон может уйти (и уходит!) от хлорофилла по цепочке переносчиков. Практически весь дальнейший путь этого электрона будет связан с окислительно-восстановительными реакциями (переносчик, получающий электрон, восстанавливается, а затем отдает электрон следующему переносчику в цепочке, восстанавливая его, а сам при этом окисляется, и так далее).
Отдавший свой электрон хлорофилл (точнее, здесь совместно работает пара молекул хлорофилла, называемая димером хлорофилла) остается со знаком «+» и становится самым сильным окислителем в биологическом мире, настолько сильным, что может отнять электрон у молекулы воды. В этом процессе участвует специальная структура — водоокисляющий комплекс, в состав которого входят четыре атома Mn, связанные с белком. Четыре марганца водоокисляющего комплекса захватывают одномоментно две молекулы воды, а дальше на каждый квант света, попавший на димер хлорофилла и приведший к уходу от хлорофилла одного возбужденного электрона, от одного из атомов марганца на «димер-с-плюсом» приходит следующий электрон. Следующий квант света — еще один возбужденный электрон уходит в цепь переносчиков от димера, и один электрон приходит на димер от марганца. Так от атомов марганца по одному уходят четыре электрона, каждый из них, попадая на димер хлорофилла, получает дополнительную энергию от фотонов и уходит дальше в цепь переносчиков. Лишившиеся четырех электронов марганцы одномоментно отнимают четыре электрона у двух молекул воды, система возвращается в исходное состояние, захватывает две новые молекулы воды и снова может поставлять электроны на димер хлорофилла. Что же останется от воды? Два атома кислорода соединятся, образуя молекулу O2 — побочный продукт фотосинтеза. А четыре протона (4H+) остаются во внутритилакоидном пространстве. Этот процесс можно назвать фотоокислением воды, но очевидно, что он не имеет ничего общего с фотолизом.
Оказывается, энергии одного кванта света недостаточно для того, чтобы сделать и восстановитель, и АТФ, поэтому электрон, путешествуя по цепи переносчиков, в некоторый момент попадает на следующий димер хлорофилла. Здесь электрон получает еще одну порцию световой энергии — еще один квант света, — чтобы в конечном итоге через несколько переносчиков попасть на молекулу-восстановителя, которая необходима для превращения СО2 в сахар.
Итак, восстановитель готов! А как же АТФ? Во время путешествия нашего электрона по цепи переносчиков при некоторых окислительно-восстановительных реакциях из стромы во внутреннее пространство тилакоида переносятся протоны (Н+). Тут надо вспомнить два важных факта, во-первых, внутреннее пространство тилакоида — замкнутое и полностью отделено от стромы мембраной, а во-вторых, в этом же пространстве накапливаются протоны, оставшиеся от воды. Таким образом, внутри тилакоида накапливается много протонов, гораздо больше, чем в строме. И оказывается, что этот «запас» протонов — это одна из форм запасания энергии, так как каждая система стремится к равновесию, и протоны из внутритилакоидного пространства будут стремиться обратно в строму, чтобы сравнять концентрации и «восстановить справедливость». Это стремление протонов восстановить равновесие использует фермент АТФ-синтаза. Понять, как работает АТФ-синтаза, нам поможет великолепный образ — представьте себе гидроэлектростанцию: вода падает с огромной высоты и крутит турбину, энергия падающей воды превращается в механическую энергию вращения турбины, а эта механическая энергия, в свою очередь, превращается в электрическую, которую мы используем на самые разные нужды. Примерно так же работает АТФ-синтаза, только не на воде, а на протонах. Протоны, стремясь вырваться из внутритилакоидного пространства, попадают в специальный канал АТФ-синтазы и, проходя его, раскручивают вращающуюся часть фермента. Энергия, запасенная в разнице концентраций протонов между внутренним пространством тилакоида и стромой, превращается в механическую энергию вращения. Вращение передается на другую часть АТФ-синтазы, которая за счет этой механической энергии присоединяет фосфат к молекуле АДФ, образуя АТФ.
Таким образом, в результате работы световой фазы фотосинтеза благодаря энергии света получаются два основных продукта:
- Восстановитель — источник электронов.
- АТФ — источник энергии.
Оба продукта световой фазы используются на следующем этапе при восстановлении СО2. Не забудем и побочный продукт световой фазы фотосинтеза — кислород, благодаря которому жизнь на нашей планете такая, какой мы ее знаем.
Темновая фаза фотосинтеза, во время которой происходит фиксация СО2, носит также название цикла Кальвина — в честь его первооткрывателя, лорда Мелвина Кальвина, который получил за это открытие Нобелевскую премию по химии в 1961 году.
Цикл Кальвина начинается с того, что фермент рибулозобисфосфаткарбоксилаза (РуБисКО или РБФК) присоединяет молекулу СО2 к пятиуглеродному сахару рибулозо-1,5-бисфосфату. Этот цикл удобно рассчитывать сразу на шесть молекул СО2, и, соответственно, 6 молекул рибулозо-1,5-бисфосфата (см. рис). Итак, в результате реакции образуется нестабильное шестиуглеродное соединение (помним, что у нас их получается шесть штук!), которое распадается на два одинаковых трехуглеродных фрагмента (у нас их будет 2×6=12 трехуглеродных фрагментов). Эти трехуглеродные соединения необходимо восстановить — здесь используем АТФ и восстановитель из световой фазы, а затем 10 (из 12-и) восстановленных трехуглеродных соединений вернутся обратно в цикл, специальный набор ферментов сделает из них снова шесть пятиуглеродных сахаров, которые мы видели в самом начале цикла. При этом еще раз придется потратить АТФ. Оставшиеся два восстановленных трехуглеродных соединения дадут нам в итоге желанный сахар.
Источник: biomolecula.ru
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
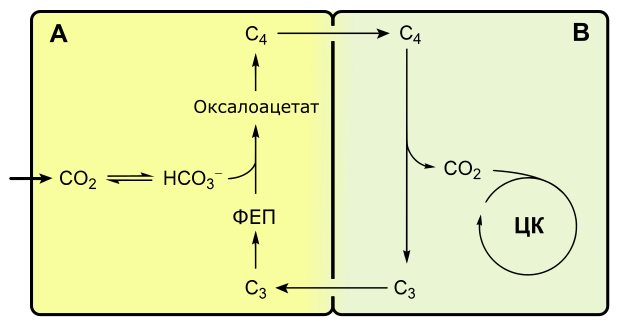
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su