Пластинчатый комплекс
Аппарат Гольджи – одномембранная, микроскопическая органелла эукариотической клетки, которая предназначена для завершения процессов синтеза клетки и обеспечивает вывод образовавшихся веществ.
Исследование структурных компонентов комплекса Гольжи началось еще в 1898 итальянским ученым-гистологом Камилло Гольджи, в честь него органелла и была названа. Изучение органоида проходило впервые в составе нервной клетки.

Строение комплекса Гольджи
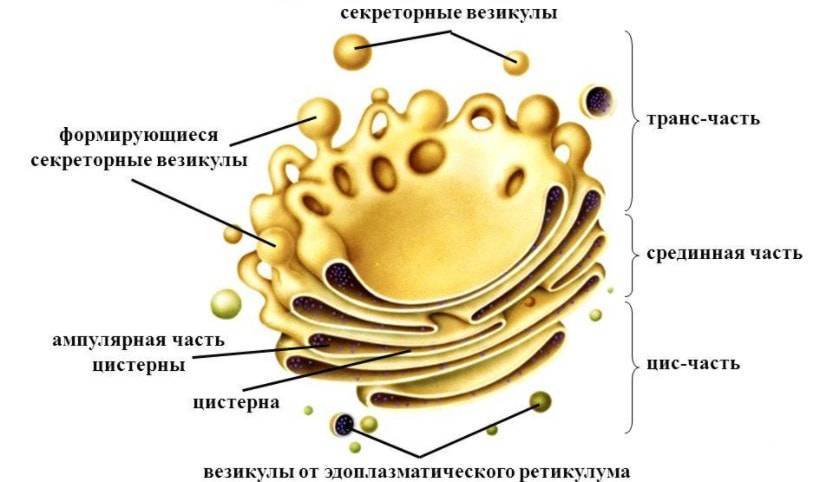
В пластинчатом комплексе (аппарат Гольджи) имеется три части:
- Цис-цистерна — находится вблизи ядра, постоянно взаимодействует с гранулярной эндоплазматической сетью;
- медиал-цистерна или промежуточная часть;
- транс-цистерна — отдаленная от ядра, дает трубчатые разветвления, формируя транс-сеть Гольджи.
Пластинчатый комплекс в клетках разной природы и даже на различных этапах дифференцировки одной клетки, иногда имеет отличительные черты в строении.
Характерные признаки аппарата Гольджи
Имеет вид стопки, которая состоит от трех до восьми цистерн, толщиной около 25 нм, они уплощены в центральной части и расширяются в направлении к периферии, напоминают стопку перевернутых тарелок. Поверхности цистерн примыкают друг к другу очень плотно. От периферической части отпочковываются небольшие мембранные пузырьки.
Клетки человека имеют одну, реже пару стопок, а клетки растений могут содержать несколько таких образований. Совокупность цистерн (одна стопка) совместно с окружающими ее пузырьками называется диктиосомой. Несколько диктиосом могут связываться между собой, формируя сеть.
Полярность – наличие цис-стороны, направленной к ЭПС и ядру, где происходит слияние везикул, и транс-стороны, устремленной к клеточной оболочке (это особенность хорошо прослеживается в клетках секретирующих органов).
Асимметричность – сторона расположенная ближе к ядру клетки (проксимальный полюс) вмещает «незрелые» белки, к ней постоянно присоединяются везикулы, отсоединившиеся от ЭПС, транс-сторона (дистальный, зрелый полюс) содержит уже модифицированные белки.
При разрушении чужеродными агентами пластинчатого комплекса, происходит разделение аппарата Гольджи на отдельные части, но его основные функции при этом сохраняются. После возобновления системы микротрубочек, которые были хаотично разбросаны в цитоплазме, части аппарата собираются, и снова превращаются в нормально функционирующий пластинчатый комплекс. Физиологическое разделение происходит и в обычных условиях жизнедеятельности клеток, во время непрямого деления.
ЭПС и комплекс Гольджи
ЭПС – это часть комплекса Гольджи?
Однозначно нет. Эндоплазматическая сеть – это самостоятельная мембранная органелла, которая построена из системы замкнутых канальцев, цистерн, сформированных непрерывной мембраной. Основная функция – синтез белков, с помощью рибосом, размещенных на поверхности гранулярной ЭПС.
Существует ряд сходных признаков между ЭПС и аппаратом Гольджи:
- Это внутриклеточные образования, отграниченные от цитоплазмы мембраной;
- отделяют мембранные пузырьки, которые наполнены органическими продуктами синтеза;
- вместе формируют единую синтезирующую систему;
- в секретирующих клетках имеют наибольшие размеры и высокий уровень развития.
Чем образованы стенки эндоплазматической сети и комплекса Гольджи?
Стенки ЭПС и аппарата Гольджи представлены в виде однослойной мембраны. Эти органеллы вместе с лизосомами, пероксисомами и митохондриями объединены в группу мембранных органоидов.
Что происходит в комплексе Гольджи с гормонами и ферментами?
За синтез гормонов отвечает эндоплазматическая сеть, на поверхности ее мембраны идет производство гормональных веществ. В комплекс Гольджи поступают синтезированные гормоны, здесь они накапливаются, затем идет переработка и выведение их наружу. Поэтому в клетках эндокринных органов встречаются комплексы больших размеров (до 10 мкм).
Функции комплекса Гольджи
Протеолиз белковых веществ, что приводит к активации белков, так проинсулин переходит в инсулин.
Обеспечивает транспорт из клетки продуктов синтеза ЭПС.
Самой важной функцией комплекса Гольджи считают выведение из клетки продуктов синтеза, поэтому его еще называют транспортным аппаратом клетки.
Синтез полисахаридов, таких как пектин, гемицеллюлоза, которые входят в состав мембран растительных клеток, образование гликозаминогликанов, одного из составляющих межклеточной жидкости.
В цистернах пластинчатого комплекса идет созревание белковых веществ, необходимых для секреции, трансмембранных протеинов клеточной мембраны, ферментов лизосом и др. В процессе созревания белки постепенно перемещаются по отделам органоида, в которых завершается их формирование и происходит гликозилирование и фосфорилирование.
Формирование липоптротеидных веществ. Синтез и накопление слизистых веществ (муцина). Образование гликолипидов, которые входят в состав мембранного гликокаликса.
Передает белки в трех направлениях: к лизосомам (перенос контролируется ферментом – маннозой- 6-фосфат), к мембранам или внутриклеточной среде, и к межклеточному пространству.
Вместе с зернистой ЭПС образует лизосомы, путем слияния отпочковавшихся везикул с автолитическими ферментами.
Экзоцитозный перенос – везикула, подойдя к мембране, встраивается в нее и оставляет свое содержимое с наружной стороны клетки.
Источник: animals-world.ru
Эндоплазматическая сеть в разных клетках может быть представлена в форме уплощенных цистерн, канальцев или отдельных везикул. Стенка этих образований состоит из билипидной мембраны и включенных в нее некоторых белков и отграничивает внутреннюю среду эндоплазматической сети от гиалоплазмы.
Различают две разновидности эндоплазматической сети:
На наружной поверхности мембран зернистой эндоплазматической сети содержатся прикрепленные рибосомы. В цитоплазме могут быть обе разновидности эндоплазматической сети, но обычно преобладает одна форма, что и обуславливает функциональную специфичность клетки. Следует помнить, что названные две разновидности являются не самостоятельными формами эндоплазматической сети, так как можно проследить переход зернистой эндоплазматической сети в гладкую и наоборот.
Функции зернистой эндоплазматической сети:
Гладкая эндоплазматическая сеть представлена цистернами, более широкими каналами и отдельными везикулами, на внешней поверхности которых отсутствуют рибосомы.
Функции гладкой эндоплазматической сети:
Пластинчатый комплекс Гольджи (сетчатый аппарат) представлен скоплением уплощенных цистерн и небольших везикул, ограниченных билипидной мембраной. Пластинчатый комплекс подразделяется на субъединицы — диктиосомы. Каждая диктиосома представляет собой стопку уплощенных цистерн, по периферии которых локализуются мелкие пузырьки. При этом, в каждой уплощенной цистерне периферическая часть несколько расширена, а центральная сужена.
В диктиосоме различают два полюса:
Установлено, что к цис-полюсу подходят транспортные вакуоли, несущие в пластинчатый комплекс продукты, синтезированные в зернистой эндоплазматической сети. От транс-полюса отшнуровываются пузырьки, несущие секрет к плазмолемме для его выведения из клетки. Однако часть мелких пузырьков, заполненных белками-ферментами, остается в цитоплазме и носит название лизосом.
Функции пластинчатого комплекса:
Среди многочисленных функций пластинчатого комплекса на первое место ставят транспортную функцию. Именно поэтому его нередко называют транспортным аппаратом клетки.
Лизосомы наиболее мелкие органеллы цитоплазмы (0,2-0,4 мкм) и поэтому открытые (де Дюв, 1949 г.) только с использованием электронного микроскопа. Представляют собой тельца, ограниченные липидной мембраной и содержащие электронноплотный матрикс, состоящий из набора гидролитических белков-ферментов (50 гидролаз), способных расщеплять любые полимерные соединения (белки, липиды, углеводы и их комплексы) на мономерные фрагменты. Маркерным ферментом лизосом является кислая фосфатаза.
Функция лизосом — обеспечение внутриклеточного пищеварения, то есть расщепления как экзогенных, так и эндогенных веществ.
Классификация лизосом:
Истинными лизосомами являются мелкие электронноплотные тельца, образующиеся в пластинчатом комплексе.
Пищеварительная функция лизосом начинается только после слияния лизосомы с фагосомой, то есть фагоцитированным веществом, окруженным билипидной мембраной. При этом образуется единый пузырек — фаголизосома, в которой смешивается фагоцитированный материал и ферменты лизосомы. После этого начинается расщепление (гидролиз) биополимерных соединений фагоцитированного материала на мономерные молекулы (аминокислоты, моносахара и так далее). Эти молекулы свободно проникают через мембрану фаголизосомы в гиалоплазму и затем утилизируются клеткой, то есть используются или для образования энергии или на построение биополимерных структур. Но не всегда фагоцитированные вещества расщепляются полностью.
Дальнейшая судьба оставшихся веществ может быть различной. Некоторые из них могут быть выведены из клетки посредством экзоцитоза, по механизму, обратному фагоцитозу. Некоторые вещества (прежде всего липидной природы) не расщепляются лизосомальными гидролазами, а накапливаются и уплотняются в фаголизосоме. Такие образования называются третичными лизосомами или остаточными тельцами.
В процессе фагоцитоза и экзоцитоза осуществляется регуляция мембран в клетке:
Установлено, что некоторые клетки в течение часа полностью обновляют плазмолемму.
Кроме рассмотренного механизма внутриклеточного расщепления фагоцитированных экзогенных веществ, таким же способом разрушаются эндогенные биополимеры — поврежденные или устаревшие собственные структурные элементы цитоплазмы. Вначале такие органеллы или целые участки цитоплазмы окружаются билипидной мембраной и образуется вакуоль аутофаголизосома, в которой осуществляется гидролитическое расщепление биополимерных веществ, как и в фаголизосоме.
Следует отметить, что все клетки содержат в цитоплазме лизосомы, но в различном количестве. Имеются специализированные клетки (макрофаги), в цитоплазме которых содержится очень много первичных и вторичных лизосом. Такие клетки выполняют защитные функции в тканях и называются клетками-чистильщиками, так как они специализированы на поглощение большого числа экзогенных частиц (бактерий, вирусов), а также распавшихся собственных тканей.
Пероксисомы — микротельца цитоплазмы (0,1-1,5 мкм), сходные по строению с лизосомами, однако отличаются от них тем, что в их матриксе содержатся кристаллоподобные структуры, а среди белков-ферментов содержится каталаза, разрушающая перекись водорода, образующуюся при окислении аминокислот.
Источник: www.medkurs.ru
Комплекс Гольджи представляет собой стопку мембранных мешочков (цистерн) и связанную с ней систему пузырьков.
На наружной, вогнутой стороне стопки из пузырьков, отпочковывающихся от глад. ЭПС, постоянно формируются новые цистерны, а на внутренней стороне цистерны превращаются обратно в пузырьки.
Основная функция комплекса Гольджи — транспорт веществ в цитоплазму и внеклеточную среду, а также синтез жиров и углеводов. Комплекс Гольджи участвует в росте и обновлении плазматической мембраны и в формировании лизосом.
Комплекс Гольджи был открыт в 1898 г. К. Гольджи. Располагая крайне примитивным оборудованием и ограниченным набором реактивов, он сделал открытие, благодаря которому совместно с Рамон-и-Кахалом получил Нобелевскую премию. Он обработал нервные клетки раствором бихромата, после чего добавил нитраты серебра и осмия. С помощью осаждения солей осмия или серебра с клеточными структурами Гольджи обнаружил в нейронах темноокрашенную сеть, которую назвал внутренним сетчатым аппаратом. При окраске общими методами пластинчатый комплекс не накапливает красителей, поэтому зона его концентрации видна как светлый участок. Например, вблизи ядра плазмоцита видна светлая зона, соответствующая области расположения органеллы.
Чаще всего комплекс Гольджи прилежит к ядру. При световой микроскопии он может распределяться в виде сложных сетей или отдельных диффузно расположенных участков (диктиосом). Форма и положение органеллы не имеют принципиального значения и могут изменяться в зависимости от функционального состояния клетки.
Комплекс Гольджи — это место конденсации и накопления продуктов секреции, вырабатываемых в других участках клетки, в основном в ЭПС. Во время синтеза белков меченные радиоизотопом аминокислоты накапливаются в гр. ЭПС, а затем их находят в комплексе Гольджи, секреторных включениях или лизосомах. Такое явление позволяет определить значение комплекса Гольджи в синтетических процессах в клетке.
При электронной микроскопии видно, что комплекс Гольджи состоит из скоплений плоских цистерн, которые называются диктиосомами. Цистерны плотно прилежат друг к другу на расстоянии 20…25 нм. Просвет цистерн в центральной части около 25 нм, а на периферии образуются расширения — ампулы, ширина которых непостоянна. В каждой стопке около 5…10 цистерн. Кроме плотно расположенных плоских цистерн в зоне комплекса Гольджи находится большое количество мелких пузырьков (везикул), особенно по краям органеллы. Иногда они отшнуровываются от ампул.
Со стороны, прилежащей к ЭПС и к ядру, в комплексе Гольджи имеется зона, содержащая значительное количество мелких пузырьков и небольших цистерн.
Комплекс Гольджи поляризован, то есть качественно неоднороден с разных сторон. Он имеет незрелую цис-поверхность, лежащую ближе к ядру, и зрелую — транс-поверхность, обращенную к поверхности клетки. Соответственно органелла состоит из нескольких взаимосвязанных компартментов, выполняющих специфические функции.
Цис-компартмент обычно обращен к клеточному центру. Его внешняя поверхность имеет выпуклую форму. С цистернами сливаются микровезикулы (транспортные пиноцитозные пузырьки), направляющиеся из ЭПС. Мембраны постоянно обновляются за счет пузырьков и, в свою очередь, восполняют содержимое мембранных образований других компартментов. В компартменте начинается посттрансляционная обработка белков, которая продолжается в следующих частях комплекса.
Промежуточный компаргмент осуществляет гликозилирование, фосфорилирование, карбоксилирование, сульфатирование биополимерных белковых комплексов. Происходит так называемая посттрансляционная модификация полипептидных цепочек. Идет синтез гликолипидов и липопротеидов. В промежуточном компартмснте, как и в цис-компартменте, формируются третичные и четвертичные белковые комплексы. Часть белков подвергается частичному протеолизу (разрушению), что сопровождается их трансформацией, необходимой для созревания. Таким образом, цис — и промежуточный компартменты необходимы для созревания белков и других сложных биополимерных соединений.
Транс-компартмент располагается ближе к периферии клетки. Внешняя поверхность его обычно вогнутая. Частично транс-компартмент переходит в транс-сеть — систему везикул, вакуолей и канальцев.
В клетках отдельные диктиосомы могут быть связаны друг с другом системой везикул и цистерн, примыкающих к дистальному концу скопления плоских мешков, так что образуется рыхлая трехмерная сеть — транс-сеть.
В структурах транс-компартмента и транс-сети происходят сортировка белков и других веществ, образование секреторных гранул, предшественников первичных лизосом и пузырьков спонтанной секреции. Секреторные пузырьки и прелизосомы окружают белки — клатрины.
Клатрины осаждаются на мембране формирующегося пузырька, постепенно отщепляя его от дистальной цистерны комплекса. Окаймленные пузырьки отходят от транс-сети, их перемещение гормонозависимое и контролируется функциональным состоянием клетки. Процесс транспортировки окаймленных пузырьков находится под влиянием микротрубочек. Белковые (клатриновые) комплексы вокруг пузырьков распадаются после отщепления пузырька от транс-сети и вновь формируются в момент секреции. В момент секреции белковые комплексы пузырьков взаимодействуют с белками микротрубочек, и пузырек транспортируется к наружной мембране. Пузырьки спонтанной секреции не окружены клатринами, их формирование происходит непрерывно и они, направляясь к клеточной мембране, сливаются с ней, обеспечивая восстановление цитолеммы.
В целом комплекс Гольджи участвует в сегрегации — это разделение, отделение определенных частей от основной массы, и накоплении продуктов, синтезированных в ЭПС, в их химических перестройках, созревании. В цистернах происходит синтез полисахаридов, их соединение с белками, что приводит к образованию сложных комплексов пептидогликанов (гликопротеинов). С помощью элементов комплекса Гольджи выводятся готовые секреты за пределы секреторной клетки.
Мелкие транспортные пузырьки отщепляются от гр. ЭПС в зонах, свободных от рибосом. Пузырьки восстанавливают мембраны комплекса Гольджи и доставляют в него полимерные комплексы, синтезируемые в ЭПС. Пузырьки транспортируются в цис-компартмент, где сливаются с его мембранами. Следовательно, в комплекс Гольджи поступают новые порции мембран и продуктов, синтезированных в гр. ЭПС.
В цистернах комплекса Гольджи происходят вторичные изменения в белках, синтезированных в гр. ЭПС. Эти изменения связаны с перестройкой олигосахаридных цепочек гликопротеинов. Внутри полостей комплекса Гольджи с помощью трансглюкозидаз модифицируются лизосомальные белки и белки секретов: происходит последовательная замена и наращивание олигосахаридных цепочек. Модифицирующиеся белки переходят от цистерны цис-компартмента в цистерны транс-компартмента за счет транспорта в пузырьках, содержащих белок.
В транс-компартменте белки сортируются: на внутренних поверхностях мембран цистерн располагаются белковые рецепторы, которые узнают секреторные белки, белки мембран и лизосом (гидролазы). В результате от дистальных транс-участков диктиосом отщепляются три типа мелких вакуолей: содержащие гидролазы — прелизосомы; с секреторными включениями, вакуоли, восполняющие клеточную мембрану.
Секреторная функция комплекса Гольджи заключается в том, что синтезированный на рибосомах экспортируемый белок, отделяющийся и накапливающийся внутри цистерн ЭПС, транспортируется в вакуоли пластинчатого аппарата. Затем накопленный белок может конденсироваться, образуя секреторные белковые гранулы (в поджелудочной, молочной и других железах), или оставаться в растворенном виде (иммуноглобулины в плазматических клетках). От ампулярных расширений цистерн комплекса Гольджи отщепляются пузырьки, содержащие эти белки. Такие пузырьки могут сливаться между собой, увеличиваться в размерах, образуя секреторные гранулы.
После этого секреторные гранулы начинают двигаться к поверхности клетки, соприкасаются с плазмолеммой, с которой сливаются их собственные мембраны, и содержимое гранул оказывается за пределами клетки. Морфологически этот процесс называется экструзией, или экскрецией (выбрасывание, экзоцитоз) и напоминает эндоцитоз, только с обратной последовательностью стадий.
Комплекс Гольджи может резко увеличиваться в размерах в клетках, активно осуществляющих секреторную функцию, что обычно сопровождается развитием ЭПС, а в случае синтеза белков — ядрышка.
Во время деления клетки комплекс Гольджи распадается до отдельных цистерн (диктиосом) и/или пузырьков, которые распределяются между двумя делящимися клетками и в конце телофазы восстанавливают структурную целостность органеллы. Вне деления происходит непрерывное обновление мембранного аппарата за счет пузырьков, мигрирующих из ЭПС и дистальных цистерн диктиосомы за счет проксимальных компартментов.
Источник: www.activestudy.info