Клеточный цикл и его периоды
Клеточный цикл — это период жизнедеятельности клетки от конца одного деления до конца следующего, который состоит, таким образом, из стадии относительного покоя, или интерфазы, и деления клетки. В интерфазе хромосомы находятся в деспирали- зированном (раскрученном) состоянии и поэтому не видны под световым микроскопом. Именно поэтому поначалу исследователи полагали, что ядро, пребывающее в неделящемся состоянии, находится в состоянии покоя. На самом деле именно в интерфазе в ядре отмечается наибольшая активность метаболических (обмен-
 |
| Дочерняя |МЕТАФАЗА| 1ТЕЛОФАЗА1 |
| центриоль |
|
I АНАФАЗА I Рис. II. 1. Клеточный цикл у эукариот |
ных и синтетических) процессов, а клетка выполняет свои обычные для нее функции или подготавливается к последующему делению (рис. 11.1).
В интерфазе выделяют три стадии: С^* — пресинтетическая, 8 — синтетическая и 02 — постсинтетическая. Если число хромосом в гаплоидном (одиночном) наборе обозначить, как «п», а количество ДНК, как «с», то в диплоидном (двойном) наборе генетического материала будет соответственно 2п2с. Сразу после предшествующего деления клетка вступает в период Это период, в течение которого клетка готовится к началу синтеза ДНК, происходящего в течение фазы 8; клетка растет, синтезируются РНК, различные белки, происходит накопление продуктов, необходимых для удвоения хромосом, увеличивается количество рибосом и митохондрий. Набор генетического материала будет 2п2с. Главное событие, происходящее в 8-фазе, — это репликация (удвоение) количества ДНК (2п4с).
В постсинтетический период (Сг2) происходит активное накопление энергии и ферментов, необходимых для последующего деления (2п4с). После Сг2-фазы наступает непрямое деление клетки, или митоз. Собственно митотическое деление занимает лишь незначительную часть клеточного цикла.
С* — читается как «джи» (название первой буквы англ. слова §ар — промежуток, интервал).
Наиболее универсальным способом деления соматических клеток, т.е. клеток тела (от греч. кота — тело), является митоз. Этот вид деления клеток был впервые описан немецким гистологом
В.Флемингом в 1882 г., который наблюдал возникновение и описал поведение нитчатых структур в ядре в период деления. Отсюда происходит и название процесса деления — митоз (от греч. тко5 — нить).
При митотическом делении ядро клетки претерпевает ряд строго упорядоченных последовательных изменений с образованием специфических нитчатых структур. В митозе выделяют несколько фаз: профазу, прометафазу, метафазу, анафазу и телофа- зу (рис. II.2).
|
а — интерфаза; б, в, г, д — различные стадии профазы; е, ж — прометафаза; метафаза; к — анафаза; л, м — телофаза; н — образование двух дочерних
2 Асанов
Профаза — первая стадия подготовки к делению. В профазе сетчатая структура ядра постепенно превращается в видимые (хромосомные) нити за счет спирализации, укорочения и утолщения хромосом. В этот период можно наблюдать двойную природу хромосом, т. к. каждая хромосома выглядит продольно удвоенной. Эти половинки хромосом (результат редупликации (удвоения) хромосом в 8-фазе), называемые сестринскими хроматидами, удерживаются вместе одним общим участком — центромерой. Начинается расхождение ценгриолей к полюсам и образование веретена деления (2п4с).
В прометафазе продолжается спирализация хромосомных нитей, происходит исчезновение ядерной оболочки, смешение кариолим- фы и цитоплазмы с образованием миксоплазмы, которая облегчает движение хромосом к экваториальной плоскости клетки (2п4с).
В метафазе все хромосомы располагаются в зоне экватора клетки, образуя так называемую «метафазную пластинку». На стадии метафазы хромосомы имеют самую малую длину, поскольку в это время они наиболее сильно с парализованы и конденсированы. Эта стадия наиболее пригодна для подсчета числа хромосом в клетке, изучения и описания их строения, определения размеров и т.д. Расположение хромосом по отношению друг к другу является случайным. Веретено деления полностью сформировано, и нити веретена прикрепляются к центромерам хромосом (2п4с).
Анафазой называют следующую фазу митоза, когда делятся центромеры хромосом. Нити веретена деления растаскивают сестринские хроматиды, которые с этого момента можно называть дочерними хромосомами, к различным полюсам клетки. Этим обеспечивается согласованное и точное распределение хромосомного материала в дочерние клетки (2п2с).
В телофазе дочерние хромосомы деспирализуются и постепенно утрачивают видимую индивидуальность. Образуется оболочка ядра, начинается симметричное разделение тела клетки с формированием двух независимых клеток (2п2с), каждая из которых вступает в период О] интерфазы. И цикл повторяется снова.
Биологическое значение митоза состоит в следующем.
1. События, происходящие в процессе митоза, приводят к образованию двух генетически идентичных дочерних клеток, каждая из которых содержит точные копии генетического материала пред- ковой (материнской) клетки.
2. Митоз обеспечивает рост и развитие организма в эмбриональном и постэмбриональном периоде. Организм взрослого человека состоит примерно из 1014 клеток, для чего требуется приблизительно 47 циклов клеточного деления единственной оплодотворенной спермием яйцеклетки (зиготы).
3. Митоз является универсальным, эволюционно закрепленным механизмом регенерации, т. е. восстановления утраченных или функционально устаревших клеток организма.
Возникновение многоклеточности сопровождается специализацией тканей организма: наряду с появлением соматических тканей (костная, мышечная, соединительная и т.д.) обособляется I ткань, дающая начало половым клеткам, — генеративная ткань.
ловое размножение возникло в процессе эволюции как высшая , форма воспроизведения организмов, позволяющая многократно (увеличивать численность потомства, и, что самое главное, половое размножение явилось необходимой предпосылкой возникновения многих форм наследственной изменчивости. Эти два фактора во многом способствовали естественному отбору наиболее приспособленных особей и тем самым существенно определяли скорость эволюционных преобразований.
При половом размножении растений и животных (в том числе Й человека) преемственность между поколениями обеспечивается •Только через половые клетки — яйцеклетку и сперматозоид. Если ‘ы яйцеклетка и сперматозоид обладали полным набором генетических характеристик (2п2с), свойственных клеткам тела, то при Их слиянии образовался бы организм с удвоенным набором (4п4с). Йапример, в соматических клетках организма человека содержится 46 хромосом. Если бы яйцеклетка и сперматозоид человека содержали по 46 хромосом, то при их слиянии образовалась бы зиго- *№с92 хромосомами. В следующем поколении проявились бы по- (Дтаоаи со 184 хромосомами и т.д.
1 Вместе с тем хорошо известно, что количество хромосом является строгой видовой характеристикой, а изменение их числа при- IВодит либо к гибели организма на ранних этапах эмбрионального (развития, либо обусловливает тяжелые заболевания. Таким образом, при образовании половых клеток должен существовать меха- рнизм, приводящий к уменьшению числа хромосом точно в два раза. Этим процессом является мейоз (от греч. шё10818 — уменьшение).
, Мейоз включает два последовательных деления. В результате Первого деления происходит уменьшение числа хромосом в ядре ровно в два раза. Именно поэтому первое деление мейоза иногда азывают редукционным делением, т. е. уменьшающим. Второе де- ние мейоза в основных чертах повторяет митоз и носит название квационного (уравнительного) деления. Мейоз состоит из ряда оследовательных фаз, в которых хромосомы претерпевают спе- фические изменения (рис. II.3). Фазы, относящиеся к первому ^Делению, обозначаются римской цифрой I, а относящиеся ко вто- 1рому — цифрой II.
В каждом делении мейоза по аналогии с митозом различают Профазу, метафазу, анафазу и телофазу.
|
|||||||||
|
|||||||||
|
|||||||||
|
|||||||||
| |
|||||||||
ч / /
7 7 ‘ ‘ -X
3 3 1/3
‘/ /^§4 11
Гс^з»4
/
Телофаза II
Рис. 11.3. Схема мейотического деления клетки
В результате мейоза образуются четыре гаплоидные клетки — гаметы На рисунке представлены три пары хромосом
К первому делению относят изменения ядра от профазы I до телофазы I.
Профаза I имеет принципиальные отличия от профазы митоза Она состоит из пяти основных стадии, лептотены, зиготены, пахитены, диплотены и диакинеза.
Самая ранняя стадия профазы I — лептотена. На этой стадии появляются тонкие перекрученные нити хромосом. Число видимых в световом микроскопе нитей равно диплоидному числу хромосом. Двойственное строение хромосомных нитей (сестринские хроматиды) постепенно выявляется по мере усиления спирализации.
На стадии зиготены происходит взаимное притяжение (конъюгация) парных или гомологичных хромосом, одна из которых была привнесена отцовской половой клеткой, другая — материнской. В митозе подобного процесса нет. Конъюгировавшая пара хромосом называется бивалентом. В нем четыре хроматиды, но они еще не различимы под микроскопом.
Стадия пахитены — самая продолжительная стадия профазы первого деления. Дальнейшая спирализация приводит к утолщению хромосом. Двойственное строение хромосом становится четко различимым: каждая хромосома состоит из двух хроматид, объединенных одной центромерой. Четыре хроматиды, объединенные попарно двумя центромерами, образуют тетраду. На стадии пахитены можно видеть ядрышки, прикрепленные к определенным участкам хромосом (области вторичных перетяжек).
В следующей стадии — диплотене — начинается процесс отталкивания друг от друга ранее конъюгировавшихся хромосом.
от процесс начинается с области центромер. Точки соприкосновения несестринских хроматид как бы сползают к концам хромосом, образуя Х-образные фигуры, называемые хиазмами. Образование хиазм сопровождается обменом гомологичных участков хроматид. Образование хиазм существенно увеличивает наследственную изменчивость благодаря появлению хромосом с новыми комбинациями аллелей за счет кроссинговера.
Последняя стадия профазы I — диакинез. В диакинезе усиливается спирализация хромосом, уменьшается число хиазм вследствие их передвижения к концам хромосом. Биваленты перемещаются в экваториальную плоскость. Исчезают оболочка ядра и ядрышки. Окончательное формирование веретена деления завершает профазу I.
В метафазе I биваленты выстраиваются в экваториальной плоскости клетки, образуя метафазную пластинку. Хромосомы при этом сильно спирализованы — утолщены и укорочены. Число бивалентов вдвое меньше, чем число хромосом в соматической клетке организма, т. е. равно гаплоидному числу.
В анафазе I гомологичные хромосомы, каждая из которых состоит из двух сестринских хроматид, расходятся к противоположным полюсам клетки. В результате этого число хромосом в каждой дочерней клетке уменьшается ровно вдвое. При этом как «отцовская», так и «материнская» хромосомы бивалента с равной вероятностью могут попадать в любую из дочерних клеток.
Телофаза I очень короткая Она характеризуется формированием новых ядер и ядерной мембраны.
Затем следует особый период — интеркинез. В интеркинезе в отличие от интерфазы митоза отсутствует 8-период и, следовательно, не происходит репликации ДНК и удвоения числа хромосом. Сестринские хроматиды перед профазой II уже удвоены.
За интеркинезом наступает второе мейотическое деление — эквационное, которое состоит из таких же фаз, как и митоз. Уже в начале второго мейотического деления клетка содержит 23 хромосомы, каждая из которых состоит из двух сестринских хроматид. В профазе II формируется новое веретено деления, в метафазе
II хромосомы вновь располагаются в экваториальной плоскости клетки. Во время анафазы II за счет деления центромеры к полюсам расходятся сестринские хроматиды, и в телофазе II образуются дочерние клетки с гаплоидным числом хромосом.
Таким образом, диплоидная клетка, вступившая в мейоз, образует четыре-дочерние клетки с гаплоидным набором хромосом.
Биологическое значение мейоза состоит в следующем.
1. Мейоз обеспечивает преемственность в ряду поколений организмов, размножающихся половым путем, в то время как митоз выполняет ту же задачу в ряду клеточных поколений.
2. Мейоз является одним из важнейших этапов процесса полового размножения.
3. В процессе мейоза происходит редукция числа хромосом от диплоидного числа (46 у человека) до гаплоидного (23).
4. Мейоз обеспечивает комбинативную наследственную изменчивость, являющуюся предпосылкой генетического разнообразия людей и генетической уникальности каждого индивида. Комбина- тивная генетическая изменчивость в процессе мейоза возникает в результате двух событий: случайного распределения негомологичных хромосом и кроссинговера, т. е. взаимного обмена гомологичных районов хроматид при образовании хиазм.
5. Мейоз называют делением созревания, поскольку формирование половых клеток (гамет) человека, как и других эукариот, связано с редукцией числа хромосом.
Источник: zakon.today
Биологическое значение деления клеток. Новые клетки возникают в результате деления уже существующих. Если делится одноклеточный организм, то из него образуются два новых. Многоклеточный организм также начинает свое развитие чаще всего с одной-единственной клетки. Путем многократных делений образуется огромное количество клеток, которые и составляют организм. Деление клеток обеспечивает размножение и развитие организмов, а значит, непрерывность жизни на Земле.
Клеточный цикл — жизнь клетки с момента ее образования в процессе деления материнской клетки до собственного деления (включая это деление) или гибели.
В течение этого цикла каждая клетка растет и развивается таким образом, чтобы успешно выполнять свои функции в организме. Далее клетка функционирует определенное время, по истечении которого либо делится, образуя дочерние клетки, либо погибает.
У различных видов организмов клеточный цикл занимает разное время: например, у бактерий он длится около 20 мин, у инфузории туфельки — от 10 до 20 ч. Клетки многоклеточных организмов на ранних стадиях развития делятся часто, а затем клеточные циклы значительно удлиняются. Например, сразу после рождения человека клетки головного мозга делятся огромное число раз: 80 % нейронов головного мозга формируется именно в этот период. Однако большинство этих клеток быстро теряет способность к делению, а часть доживает до естественной смерти организма, вообще не делясь.
Клеточный цикл состоит из интерфазы и митоза (рис. 54).
Интерфаза — промежуток клеточного цикла между двумя делениями. В течение всей интерфазы хромосомы неспирализованы, они находятся в ядре клетки в виде хроматина. Как правило, интерфаза состоит из трех периодов: пре-синтетического, синтетического и постсинтетического.
Пресинтетический период (G,) — наиболее продолжительная часть интерфазы. Он может продолжаться у различных типов клеток от 2—Зч до нескольких суток. Во время этого периода клетка растет, в ней увеличивается количество органоидов, накапливается энергия и вещества для последующего удвоения ДНК- В течение Gj-периода каждая хромосома состоит из одной хроматиды, т. е. количество хромосом (п) и хроматид (с) совпадает. Набор хромосом и хро-
матид (молекул ДНК) диплоидной клетки в Grпериоде клеточного цикла можно выразить записью 2п2с.
В синтетическом периоде (S) происходит удвоение ДНК, а также синтез белков, необходимых для последующего формирования хромосом. В этот же период происходит удвоение центриолей.
Удвоение ДНК называют репликацией. В ходе репликации специальные ферменты разъединяют две цепи исходной материнской молекулы ДНК, разрывая водородные связи между комплементарными нуклеотидами. С разошедшимися цепями связываются молекулы ДНК-полимеразы — главного фермента репликации. Затем молекулы ДНК-полимеразы начинают двигаться вдоль материнских цепей, используя их в качестве матриц, и синтезировать новые дочерние цепи, подбирая для них нуклеотиды по принципу комплементарности (рис. 55). Например, если участок материнской цепи ДНК имеет последовательность нуклеотидов А Ц Г Т Г А, то участок дочерней цепи будет иметь вид ТГЦАЦТ. В связи с этим репликацию относят к реакциям матричного синтеза. В результате репликации образуются две идентичные двуцепочечные молекулы ДНК- В состав каждой из них входит одна цепочка исходной материнской молекулы и одна вновь синтезированная дочерняя цепочка.
К концу S-периода каждая хромосома состоит уже из двух идентичных сестринских хроматид, соединенных друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2п4с.
Постсинтетический период (G2) наступает после удвоения ДНК- В это время клетка накапливает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего С2-периода набор хромосом и хроматид в клетке остается неизменным — 2п4с.
Интерфаза завершается, и начинается деление, в результате которого образуются дочерние клетки. В ходе митоза (основного способа деления клеток эукариот) сестринские хроматиды каждой хромосомы отделяются друг от друга и попадают в разные дочерние клетки. Следовательно, молодые дочерние клетки, вступающие в новый клеточный цикл, имеют набор 2п2с.
Таким образом, клеточный цикл охватывает промежуток времени от возникновения клетки до ее полного разделения на две дочерние и включает интерфазу (Gr, S-, С2-периоды) и митоз (см. рис. 54). Такая последовательность периодов клеточного цикла характерна для постоянно делящихся клеток, например для клеток росткового слоя эпидермиса кожи, красного костного мозга, слизистой оболочки желудочно-кишечного тракта животных, клеток образовательной ткани растений. Они способны делиться каждые 12—36 ч.
В противоположность этому большинство клеток многоклеточного организма встают на путь специализации и после прохождения части Gj-периода могут переходить в так называемый период покоя (Go-период). Клетки, пребывающие в Gn-периоде, выполняют свои специфические функции в организме, в них протекают процессы обмена веществ и энергии, но не происходит подготовка к репликации. Такие клетки, как правило, навсегда утрачивают способность к делению. Примерами могут служить нейроны, клетки хрусталика глаза и многие другие.
Однако некоторые клетки, находящиеся в Gn-периоде (например, лейкоциты, клетки печени), могут выходить из него и продолжать клеточный цикл, пройдя все периоды интерфазы и митоз. Так, клетки печени могут снова приобретать способность к делению спустя несколько месяцев пребывания в периоде покоя.
Клеточная гибель. Гибель (смерть) отдельных клеток или их групп постоянно встречается у многоклеточных организмов, так же как гибель одноклеточных организмов. Гибель клеток можно разделить на две категории: некроз (от греч. некрос — мертвый) и ап о птоз, который часто называют программируемой клеточной смертью или даже клеточным самоубийством.
Некроз — отмирание клеток и тканей в живом организме, вызванное действием повреждающих факторов. Причинами некроза может быть воздействие высоких и низких температур, ионизирующих излучений, различных химических веществ (в том числе токсинов, выделяемых болезнетворными микроорганизмами). Некротическая гибель клеток наблюдается также в результате их механического повреждения, нарушения кровоснабжения и иннервации тканей, при аллергических реакциях.
В повре>вденных клетках нарушается проницаемость мембран, останавливается синтез белков, прекращаются другие процессы обмена веществ, происходит разрушение ядра, органоидов и, наконец, всей клетки. Особенностью некроза является то, что такой гибели подвергаются целые группы клеток (например, при инфаркте миокарда из-за прекращения снабжения кислородом отмирает участок сердечной мышцы, содержащий множество клеток). Обычно отмирающие клетки подвергаются атаке лейкоцитов, и в зоне некроза развивается воспалительная реакция.
Апоптоз — запрограммированная гибель клеток, регулируемая организмом. В ходе развития и функционирования организма часть его клеток погибает без непосредственного повреждения. Этот процесс протекает на всех стадиях жизни организма, даже в эмбриональный период.
Во взрослом организме также постоянно происходит запланированная гибель клеток. Миллионами гибнут клетки крови, эпидермиса кожи, слизистой оболочки желудочно-кишечного тракта и др. После овуляции погибает часть фолликулярных клеток яичника, после лактации — клетки молочных желез. В организме взрослого человека ежедневно в результате апоптоза гибнет 50—70 миллиардов клеток. При апоптозе клетка распадается на отдельные фрагменты, окруженные плазмалеммой. Обычно фрагменты погибших клеток поглощаются лейкоцитами или соседними клетками без запуска воспалительной реакции. Восполнение утраченных клеток обеспечивается путем деления.
Таким образом, апоптоз как бы прерывает бесконечность клеточных делений. От своего «рождения» до апоптоза клетки проходят определенное количество нормальных клеточных циклов. После каждого из них клетка переходит либо к новому клеточному циклу, либо к апоптозу.
Источник: botana.biz
Живые организмы состоят из клеток, рост и деление которых нуждается в запрограммированных последовательных событиях и процессах, составляющих клеточный цикл. Некоторые из этих процессов происходят непрерывно, как, например, синтез белков и липидов. Другие, как, например, синтез ДНК, носят прерывистый характер и связаны с процессом клеточного деления.
В каждом клеточном цикле объединяются два динамических и качественно различных периода:
- интерфаза,
- митоз
Необходимо подчеркнуть, что процесс клеточного размножения основан на репликации ДНК, синтез которой происходит в периоде S интерфазы клеточного цикла. Интерфаза является наиболее длительным периодом клеточного цикла (90%), этот период представляет собой этап, когда клетка имеет интенсивную биосинтетическую активность (синтез ДНК, РНК, белков), обеспечивая необходимые условия для осуществления клеточного деления. Интерфаза подразделяется на три периода, известные как:
- Период G1 – пресинтетический или постмитотический период, во время которого: — возобновляется процесс транскрипции и белкового синтеза, блокированные во время митоза; — происходит деконденсация хроматина – важный процесс для активации транскрипции генов; — происходит реорганизация ядрышек; — клетка содержит диплоидный набор монохроматидных хромосом (2n = 2с).
- Период S – синтетический период, характеризуется: — полуконсервативной асинхронной репликацией молекул ДНК; — удвоением количества ДНК; — двухроматидными хромосомами (2n = 4с); — параллельным синтезом гистоновых и негистоно- вых белков, участвующих в синтезе и упаковке ДНК; — удвоением центриолей.
- Период G2 – постсинтетический или премитотический, в котором процессы транскрипции и белкового синтеза происходят с той же интенсивностью, что и в периоде G1, это обеспечивает необходимые условия для митоза.
Митоз и цитокинез. Клеточное деление начинается делением ядра (митоз) и завершается делением цитоплазмы (цитокинез). Митоз и цитокинез занимают короткий период клеточного цикла (10%) и носят название митотического периода. Митоз, однажды начавшись, представляет собой непрерывный процесс. Однако для изучения и описания его условно подразделяют на четыре этапа:
- профазу,
- метафазу,
- анафазу,
- телофазу.
Профаза характеризуется наличием в ядре двухроматидных хромосом. Эти хроматиды, соединенные на уровне центромеры, представляют собой цитологическую картину процесса репликации ДНК. Хромосомы сильно конденсируются, утолщаются и становятся видимыми; по обе стороны центромеры образуются по одному кинетохору. В конце профазы исчезает ядрышко и диссоциирует ядерная оболочка. Одновременно организуется аппарат деления: центриоли перемещаются к противоположным полюсам клетки, формируется веретено деления путем сборки микротрубочек.
Метафаза. Нити веретена деления связывают центриоль и хромосому с помощью кинетохоров. Хромосомы, соединенные с веретеном деления, располагаются в экваториальной плоскости, образуют метафазную пластинку. На этой стадии хромосомы максимально спирализованы и представляют собой оптимальную форму для цитологического изучения.
Анафаза начинается с продольного разделения центромеры каждой хромосомы, расхождения сестринских хроматид и завершается одновременной их миграцией к противоположным полюсам клетки. На этом этапе хромосомы становятся монохроматидными, и клетка содержит тетраплоидный набор хромосом (4n = 4с).
Телофаза. В телофазе заканчивается миграция хромосом к полюсам клетки, у каждого полюса содержится 2n монохроматидных хромосом (диплоидный набор). Начинается прогрессирующая деспирализация хромосом и возврат наследственного материала в состояние интерфазного хроматина. Нити веретена деления диссоциируют, вновь появляются ядерные мембраны вокруг каждой группы хромосом, вновь организуются ядрышки.
Цитокинез завершает процесс деления. Происходит разделение цитоплазматической массы на две половины и разделение клеточных органелл. Каждая дочерняя клетка наследует в результате цитокинеза набор клеточных компонентов. Увеличение количества всех компонентов клетки не требует точного контроля. Если в клетке имеется много молекул или органелл определенного типа, то достаточно того, чтобы число их приблизительно удвоилось за цикл, и они затем примерно поровну разделились между двумя дочерними клетками. Рост органелл происходит задолго до начала цитокинеза. Увеличение количества клеточных органелл реализуется различными путями. Митохондрии растут и делятся полуавтономно, аппарат Гольджи и ЭПС фрагментируются на пузырьки, которые служат для образования новых клеточных органелл, в то время как рибосомы размножаются путем образования комплекса рРНК и рибосомальных белков. В отношении ДНК такое удвоение и распределение должно быть совершенно точно, и для этого нужен специальный механизм: репликация ядерной ДНК. Этот механизм обеспечивает образование генетически идентичных клеток, как между собой, так и с материнской клеткой.
Митотическое деление является основой размножения соматических клеток, обеспечивает эмбриогенез, рост многоклеточного организма, определяет биомассу организма и регенерацию тканей.
В настоящее время цитометрию принято подразделять на:
- проточную,
- статическую.
Первый вариант цитометрии осуществляется с помощью специальных приборов – проточных цитометров и сортеров. Для статической цитометрии могут быть использованы конфокальные микроскопы, а также более простые и дешевые системы анализа изображений, смонтированные на обычных люминесцентных микроскопах.
Существует много методик, которые с одинаковым успехом можно воспроизводить как с помощью проточной, так и статической цитометрии. Более того, статическая цитометрия в некоторых случаях позволяет получить более обширную информацию о клетках, причем ее производительность ненамного меньше проточной.
Метод проточной цитометрии сформировался за последние 30 лет на основе отдельных опытов по подсчету числа частиц и определению их размеров.
В настоящее время выпускают два основных типа приборов для проточной цитометрии :
- простые в использовании аппараты, которые могут измерять флуоресценцию при двух и более длинах волн и светорассеяние под углом около 10º (малоугловое прямое рассеяние) и 90º;
- большие клеточные сортеры, которые не только измеряют пять и более клеточных или ядерных параметров, но и сортируют частицы с заданным набором этих параметров.
Принципы проточной цитометрии весьма просты. Клетки или ядра поодиночке пересекают сфокусированный световой пучок, обычно лазерный. Свет определенной длины возбуждает молекулы флуоресцирующих красителей, связанных с различными клеточными компонентами, при этом при этом может происходить одновременное возбуждение нескольких разных красителей, что позволяет оценить сразу несколько клеточных параметров. Свет, испускаемый красителями, собирают с помощью системы линз и зеркал и разлагают на компоненты. Световые сигналы детектируют, преобразуют в электрические импульсы и далее в форму, удобную для компьютерной обработки и хранения информации.
Методом проточной цитометрии можно получать самые разные данные:
- определять содержание в клетке ДНК и РНК, суммарное количество белков и количество специфических белков, узнаваемых моноклональными антителами,
- исследовать клеточный метаболизм (например, измерять внутриклеточный рН), изучать транспорт ионов кальция и кинетику ферментативных реакций.
Источник: vseobiology.ru
КЛЕ́ТОЧНЫЙ ЦИКЛ, набор строго скоординированных во времени процессов, приводящих к воспроизведению клеток; важнейший механизм, с помощью которого репродуцируется всё живое. В основе способности клеток к самовоспроизведению лежит уникальное свойство ДНК реплицироваться с образованием двух точных копий, которые в ходе деления распределяются между двумя дочерними клетками. Помимо удвоения ДНК в К. ц. большинства клеток должно удваиваться число их органелл и макромолекул, в противном случае с каждым делением клетки становились бы меньше. Т. о., для поддержания своих размеров клетка должна координировать рост клеточной массы с её делением. У одноклеточных организмов К. ц. совпадает с жизненным циклом. В непрерывно размножающихся клетках многоклеточных организмов он состоит из чётко сменяющих друг друга периодов: интерфазы (подготовки к делению) и митоза (собственно деления).
Именно в интерфазе, а не в митозе, как считали ранее, происходит репликация ДНК (удвоение хромосом). Сама интерфаза подразделяется на три фазы. Пресинтетическая G1-фаза предшествует синтезу ядерной ДНК и следует сразу за предыдущим делением. В этой фазе в клетке синтезируются рибонуклеиновые кислоты (РНК), разл. белки, увеличивается число рибосом, митохондрий. В целом наблюдается активный рост клетки. В фазе синтеза ДНК (S-фазе) продолжается образование РНК и белков, происходит репликация ядерной ДНК. Постсинтетическая G2-фаза характеризуется окончательной подготовкой клетки к делению, в т. ч. сборкой микротрубочек, которые в дальнейшем будут формировать веретено деления. За фазами интерфазы следует митоз (М-фаза). Понятие «К. ц.» включает в себя не только интервал времени от одного клеточного деления до другого. Иногда клетка, завершившая митоз, прекращает подготовку к очередным синтезу ДНК и митозу и входит в состояние «вне цикла», т. е. в состояние пролиферативного покоя, обозначаемое как G0-фаза. Из этой фазы она может вновь вернуться в состояние К. ц. под влиянием стимула к размножению. Существуют также клетки с удлинённой продолжительностью постсинтетической фазы. Их называют G2-популяцией.
Длительность К. ц. варьирует у разных организмов в широких пределах – от нескольких минут до нескольких суток. Показано также, что разные клеточные популяции могут сильно различаться и по продолжительности отд. фаз цикла. Наиболее коротким является К. ц. бластомеров на ранних стадиях эмбрионального развития. В К. ц. этих клеток практически отсутствуют G1- и G2-фазы, S-фаза может длиться всего неск. минут. На заключит. этапе эмбрионального развития животных, напр. у млекопитающих, происходит массовое вступление клеток в состояние покоя, связанное с их дифференцировкой. Длительность К. ц. различается в пределах одного взрослого многоклеточного организма. Напр., клетки регенерирующей печени мыши делятся примерно через каждые 48 ч, а крипт кишки (углублений эпителия в её слизистой) – через 10,5 ч. Лимфоциты человека, помещённые в среду для культивирования клеток и обработанные фитогемагглютинином, начинают размножаться, и их К. ц. оказывается равным примерно 24 часам. Наиболее вариабельной является G1-фаза. При дифференцировке одни типы клеток взрослого организма, выполняя специфич. функции, находятся в G0-фазе, но сохраняют способность к размножению, другие – необратимо утрачивают её и называются терминально дифференцированными.
Программа осуществления процессов К. ц. включает сложную систему контроля, которая обеспечивается работой комплексов регуляторных белков. При этом контролирующие системы действуют как внутри клетки (эндогенные регуляторы), так и вне её (экзогенные регуляторы). Эндогенная регуляция клеточного цикла основана на циклич. активации протеинкиназ, известных как циклин-зависимые киназы. Активность последних увеличивается или падает в ходе К. ц. Такие колебания активности ведут к циклич. изменениям степени фосфорилирования внутриклеточных белков, инициирующих или регулирующих важнейшие события К. ц.: репликацию ДНК, сегрегацию хромосом и цитокинез (собственно деление тела клетки на две). У разных организмов в разных фазах К. ц. активируются разл. варианты комплексов циклинов и киназ. Прекращение работы таких комплексов связано с действием ингибирующих белков или с протеолизом циклинового компонента комплекса. В целом эндогенная регуляция в ходе К. ц. обеспечивается сложной системой процессов синтеза и деградации определённых субъединиц регуляторных комплексов, процессами фосфорилирования и дефосфорилирования этих и др. регуляторых белков. Экзогенная регуляция включает действие на клетки внеклеточных регуляторных сигналов. Для одноклеточных организмов такими сигналами являются компоненты окружающей среды, для многоклеточных организмов – синтезируемые разл. клетками небольшие белковые молекулы, к которым относятся факторы роста и цитокины. Нарушение механизмов эндогенной и экзогенной регуляции К. ц. может приводить к неконтролируемому размножению клеток, что служит причиной образования опухолей.
Источник: bigenc.ru
Введение
Для того чтобы клетка смогла полноценно разделиться, она должна увеличиться в размерах и создать достаточное количество органоидов. А для того чтобы не растерять наследственную информацию при делении пополам, она должна изготовить копии своих хромосом. И, наконец, для того чтобы распределить наследственную информацию строго поровну между двумя дочерними клеткам, она должна в правильном порядке расположить хромосомы перед их распределением по дочерним клеткам. Все эти важные задачи решаются в процессе клеточного цикла.
Клеточный цикл имеет важное значение, т.к. он демонстрирует важнейшие свойства клетки: способность к размножению, росту и дифференцировке. Обмен тоже идёт, но его не рассматривают при изучении клеточного цикла.
Определение понятия
Клеточный цикл — это период жизни клетки от рождения до образования дочерних клеток.
У животных клеток клеточный цикл, как промежуток времени между двумя делениями (митозами), длится в среднем от 10 до 24 часов.
Клеточный цикл состоит из нескольких периодов (синоним: фазы), которые закономерно сменяют друг друга. В совокупности первые фазы клеточного цикла (G1, G0, S и G2) носят название интерфазы, а последняя фаза называется митозом.
Рис. 1. Клеточный цикл.
Периоды (фазы) клеточного цикла
1. Период первого роста G1 (от английского Growth — рост), составляет 30-40% цикла, и период покоя G0
Синонимы: постмитотический (наступает после митоза) период, пресинтетический (проходит перед синтезом ДНК) период.
Клеточный цикл начинается от рождения клетки в результате митоза. После деления дочерние клетки уменьшены в размерах и органоидов в них меньше, чем в норме. Поэтому «новорожденная» маленькая клетка в первом периоде (фазе) клеточного цкла (G1) растёт и увеличивается в размерах, а также формирует недостающие органоиды. Идёт активный синтез белков, необходимых для ввсего этого. В результате клетка становится полноценной, можно сказать, «взрослой».
Чем обычно заканчивается для клетки период роста G1?
- Вступллением клетки в процесс дифференцировки. За счёт дифференцировки клетка приобретает специальные особенности для выполнения функций, необходимых всему органу и организму. Запускается дифференцировка управляющими веществами (гормонами), воздействующими на соответствующие молекулярные рецепторы клетки. Клетка, завершившая свою дифференцировку, выпадает из круговорота делений и находится в периоде покоя G0. Требуется воздействие активирующих веществ (митогенов) для того, чтобы она претерпела дедифференцировку и вновь вернулась в клеточный цикл.
- Гибелью (смертью) клетки.
- Вступлением в следующий период клеточного цикла -синтетический.
2. Синтетический период S (от английского Synthesis — синтез), составляет 30-50% цикла
Понятие синтеза в названии этого периода относится к синтезу (репликации) ДНК, а не к каким-либо другим процессам синтеза. Достигнув определенного размера в результате прохождения периода первого роста, клетка вступает в синтетический период, или фазу, S, в котором происходит синтез ДНК. За счёт репликации ДНК клетка удваивает свой генетический материал (хромосомы), т.к. в ядре образуется точная копия каждой хромосомы. Каждая хроммосома становится двойной и весь хромосомный набор становится двойным, или диплоидным. В результате клетка теперь готова поделить наследственный материал поровну между двумя дочерними клетками, не потеряв при этом ни одного гена.
3. Период второго роста G2 (от английского Growth — рост), составляет 10-20% цикла
Синонимы: премитотический (проходит перед митозом) период, постсинтетический (наступает после синтетического) период.
Период G2 является подготовительным к очередному делению клетки. Во время второго периода роста G2 клетка производит белки, требующиеся для митоза, в частности, тубулин для веретена деления; создаёт запас энергии в виде АТФ; проверяет, закончена ли репликация ДНК, и готовится к делению.
4. Период митотического деления M (от английского Mitosis — митоз), составляет 5-10% цикла
Митоз M (синоним: митотический цикл), заключается в том, что клетка правильно делится на две дочерние клетки. Благодаря механизмам комплементарного синтеза при репликации ДНК в синтетическом периоде и механизму распределения хроматид в митозе каждая дочерняя клетка получает идентичный набор хромосом, являющийся точной копией хромосомного набора материнской клетки. Короче говоря, за счёт танцевв хромосом, они расределяются пополам и поровну между двумя дочерними клетками, образовавшимися в результате деления. Подробнее о митозе…
После деления клетка оказывается в новой фазе G1, и клеточный цикл завершается.
Рис. 2. Клеточный цикл. Источник изображения: http://pisum.bionet.nsc.ru/kosterin/lectures/lecture9/lecture9.htm
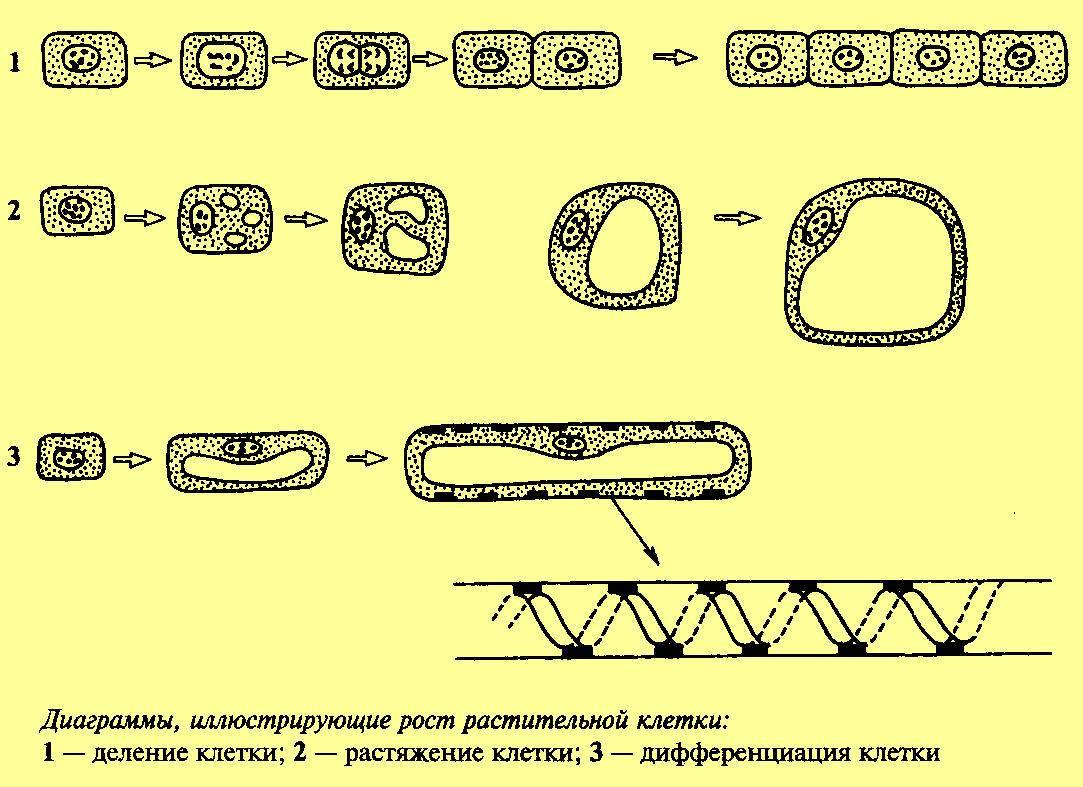
Рис. 3. Клеточный цикл растительных клеток. Источник изображения: http://fizrast.ru/razvitie/rost/osobennosti.html
Регуляция клеточного цикла
На молекулярном уровне переход от одной фазы цикла к другой регулируют два белка — циклин и циклинзависимая киназа (CDK).
Для регуляции клеточного цикла используется процесс обратимого фосфорилирования/дефосфорилирования регуляторных белков, т.е. присоединение к ним фосфатов с последующим отщеплением. Ключевым веществом, регулирующим вступление клетки в митоз (т.е. её переход от фазы G2 к фазе M), является специфическая серин/треонин-протеинкиназа, которая носит название фактор созревания — ФС, или MPF, от английского maturation promoting factor. В активной форме этот белковый фермент катализирует фосфорилирование многих белков, принимающих участие в митозе. Это, например, входящий в состав хроматина гистон H1, ламин (компонент цитоскелета, находящийся в ядерной мембране), факторы транскрипции, белки митотического веретена, а также ряд ферментов. Фосфорилирование этих белков фактором созревания MPF активирует их и запускает процесс митоза. После завершения митоза регуляторная субъединица ФС, циклин, маркируется убиквитином и подвергается распаду (протеолизу). Теперь наступает очередь протеинфосфатаз, которые дефосфорилируют белки, принимавшие участие в митозе, чем переводят их в неактивное состояние. В итоге клетка возвращается в состояние интерфазы.
ФС (MPF) — это гетеродимерный фермент, включающий в себя регуляторную субъединицу, а именно циклин, и каталитическую субъединицу, а именно циклинзависимую киназу ЦЗК (CDK от англ. cyclin dependent kinase), она же p34cdc2; 34 кДа. Активной формой этого фермента является лишь димер ЦЗК+циклин. Кроме того, активность ЦЗК регулируется путем обратимого фосфорилирования самого фермента. Циклины получили такое название потому, что их концентрация циклически изменяется в соответствии с периодами клеточного цикла, в частности, она снижается перед началом деления клетки.
В клетках позвоночных присутствует ряд различных циклинов и циклинзависимых киназ. Разнообразные сочетания двух субъединиц фермента регулируют запуск митоза, начало процесса транскрипции в G1-фазе, переход критической точки после завершения транскрипции, начало процесса репликации ДНК в S-периоде интерфазы (стартовый переход) и другие ключевые переходы клеточного цикла (на схеме не приведены).
В ооцитах лягушки вступление в митоз (G2/M-переход) регулируется путем изменения концентрации циклина. Циклин непрерывно синтезируется в интерфазе до достижения максимальной концентрации в фазе М, когда запускается весь каскад фосфорилирования белков, катализируемый ФС. К окончанию митоза циклин быстро разрушается протеиназами, также активируемыми ФС. В других клеточных системах активность ФС регулируется за счет различной степени фосфорилирования самого фермента.
Источник: kineziolog.su