Водоросли размножаются спорами
У водорослей встречается вегетативное, бесполое и половое размножение.
К вегетативному размножению относят те процессы, при которых части таллома отделяются без каких-либо заметных изменений в протопластах и дочерние клетки получают часть оболочки материнской. Так, у некоторых одноклеточных водорослей оно происходит путем деления клетки пополам (например, у эвглены, диатомовых водорослей, одноклеточных конъюгат, синезеленых).
 |
|
|
У эвгленовых такой способ размножения – единственный в жизненном цикле, у других он может существовать наряду с половым (например, диатомеи и конъюгаты). Но не все одноклеточные водоросли могут размножаться вегетативно. Так, например хламидомонада и хлорелла вегетативно не размножаются! Некоторые колониальные представителей могут размножаться участками колоний (например, синура), но не все колониальные водоросли имеют вегетативное размножение. Например, вольвокс, сценедесмус вегетативно не размножаются!
|
|
У нитчатых водорослей вегетативное размножение происходит за счет распада нитей на отдельные фрагменты, иногда и одноклеточные (например, спирогира).
| |
У бурых водорослей из порядка сфацеляриевых существуют специализированные выводковые веточки, а у фукуса на подошве имеются группы клеток, способных к дальнейшему развитию в новый таллом, поэтому можно наблюдать, как от одной подошвы у фукуса могут отходить несколько талломов. Вегетативно размножаются виды саргассов, обитающих в Саргассовом море.
| |
У представителей порядка харовых для вегетативного размножения имеются на ризоидах клубеньки и группы звездчатых клеток на вертикальной части таллома.
Бесполое размножение у водорослей осуществляется с помощью подвижных (зооспоры) и неподвижных (апланоспоры) спор.
орообразование обычно сопровождается делением протопласта на части и выходом продуктов деления из оболочки материнской клетки. Из содержимого каждой клетки реже может сформироваться только одна зооспора (например, эдогониум, вошерия), но чаще содержимое клетки делится на две, четыре, восемь и более частей и образуется соответствующее количество спор бесполого размножения. Некоторые колониальные водоросли размножаются бесполым путем с помощью дочерних колоний (например, вольвокс, сценедесмус).
| |
|
У одних водорослей перед образованием спор бесполого размножения происходит мейоз (например, морская капуста, многие красные водоросли, кладофора), у других при образовании спор бесполого размножения происходит только митоз (например, хламидомонада, хлорелла). Клетки, в которых формируются споры бесполого размножения, называются спорангиями, а особь, на которой формируются спорангии — спорофитом. Бесполое размножение с помощью зооспор встречается у хламидомонады, улотрикса, кладофоры, морской капусты, ульвы.
| |
Бесполое размножение неподвижными спорами встречается у красных водорослей, хлореллы, иногда у улотрикса и др. В ряде случаев подвижные и неподвижные споры бесполого размножения носят особые названия. Например, апланоспоры, одевающиеся оболочкой и приобретающие форму материнской клетки, находясь внутри нее, называются автоспорами (например, хлорелла). Бесполое размножение у некоторых водорослей бывает единственным способом размножения в жизненном цикле (например, хлорелла). В то же время у ряда водорослей в жизненном цикле бесполое размножение с помощью спор отсутствует. Например нет бесполого размножения у диатомей, у конъюгат.
При половом размножении в результате попарного слияния гаплоидных клеток образуется диплоидная зигота. Половой процесс у водорослей может происходить с участием или без участия гамет. Типы полового процесса с участием гамет следующие: изогамия — слияние одинаковых по размеру и форме подвижных гамет (например, у некоторых видов хламидомонады, улотрикса, кладофоры); гетерогамия — слияние подвижных гамет одинаковой формы, но разного размера (например, у некоторых видов хламидомонады); оогамия — слияние крупной неподвижной женской гаметы яйцеклетки с мелким подвижным сперматозоидом (например, у некоторых видов хламидомонады, у морской капусты). Клетки, в которых формируются гаметы, называются гаметангиями, а особь, на которой они образуются — гаметофитом. Клетку, в которой формируются сперматозоиды, называют антеридием, а клетку, содержащую яйцеклетку (одну или несколько), – оогонием. У красных водорослей мужская половая клетка лишена подвижности и называется спермацием.
Эволюция формы полового процесса происходила независимо от эволюции таллома. Это положение впервые продемонстрировал русский альголог И.Н. Горожанкин, обнаружив у разных видов хламидомонады (стоящих на одном уровне морфологической дифференцировки таллома) все основные типы полового процесса: наряду с изогамией, гетерогамию и оогамию.
Типы полового процесса у водорослей без образования гамет следующие: хологамия (гологамия) – слияние двух подвижных (жгутиковых) одноклеточных особей; конъюгация — слияние протопластов двух гаплоидных безжгутиковых вегетативных клеток с образованием диплоидной зиготы. Хологамия встречается у одноклеточной подвижной водоросли дуналиеллы, которая встречается в гиперсоленых водоемах. Конъюгация встречается только у группы конъюгат из харовых водорослей.
При конъюгации две нити спирогиры подходят друг к другу, между противоположными клетками двух нитей начинают формироваться копуляционные каналы. Содержимое клеток сжимается, и протопласт из одной клетки через копуляционный канал перемещается в противоположную клетку, где и происходит слияние двух протопластов с образованием диплоидной зиготы. Вторая клетка остается пустой.
Конъюгация встречается как у нитчатых, так и у одноклеточных и колониальных конъюгат.
Гаметофиты и спорофиты в жизненном цикле у водорослей могут внешне не различаться (изоморфная смена форм развития) или иметь хорошо выраженные морфологические различия (гетероморфная смена форм развития). Изоморфная смена форм развития встречается у кладофоры, улотрикса, гетероморфная – у морской капусты, порфиры. У однодомных (обоеполых) видов мужские и женские гаметы развиваются на одной особи, у двудомных (раздельнополых) на разных.
Размножение грибов
Грибы могут размножаться вегетативно, бесполым и половым путем.
Вегетативное размножение у мицелиальных грибов осуществляется фрагментами мицелия. Дрожжи вегетативно размножаются почкованием. У них на клетке образуется выпячивание, которое разрастается, а затем отделяется от материнской клетки.
Бесполое размножение у грибов осуществляется эндогенными (формируются внутри спорангиев) и экзогенными (формируются на концах спороносца свободно) спорами. Эндогенные споры могут быть подвижными (зооспоры) и неподвижными (спорангиоспоры, споры). Зооспоры встречаются у хитридиомицетов, в отличие от зооспор водорослей они имеют единственный задний жгутик.
Бесполое размножение с помощью эндогенных неподвижных характерно для мукора, у которого они развиваются в спорангиях,. Экзогенные споры или конидиями формируются на конидиеносцах — специальных гифах мицелия. Они могут быть одиночными или собраны в цепочки, как у пеницилла.
Половое размножение у грибов бывает трех типов: гаметогамия, гаметангиогамия и соматогамия. Половой процесс у грибов может быть изогамным, гетерогамным, оогамным (эти типы полового процесса встречаются у хитридиомицетов). При гаметангиогамии у грибов сливается содержимое гаметангиев, недифференцированных на гаметы (например, мукор). У шляпочных грибов половой процесс — соматогамия, когда сливается содержимое вегетативных клеток. При этом у шляпочных грибов слияние клеток происходит без слияния ядер, и плодовые тела у них состоят из клеток, содержащих 2 гаплоидных несестринских ядра Такая стадия называется дикарион и обозначается n+n. Слияние ядер осуществляется позднее, перед формированием в шляпках спор.
Источник: distant.msu.ru
Общая характеристика водорослей
Водоросли — это многоклеточные, преимущественно водные, эукариотические фотосинтезирующие организмы, которые не имеют тканей или тело которых не дифференцировано на вегетативные органы (т.е. относящиеся к подцарству низших растений).
❖ Систематические отделы водорослей (различаются по струк туре таллома, набору фотосинтезирующих пигментов и запасных питательных веществ, особенностям размножения и циклов развития, местообитанию и т. п.):
■ Золотистые;
■ Зеленые (примеры: спирогира, улотрикс);
■ Красные (примеры: порфира, филлофора);
■ Бурые (примеры: лессония, фукус);
■ Харовые (примеры: хара, нителла);
■ Диатомовые (пример: ликмофора) и др.
Количество видов водорослей — более 40 тыс.
❖ Среда обитания водорослей: пресные и соленые водоемы, влажная почва, кора деревьев, горячие источники, ледники и т.д.
❖ Экологические группы водорослей: планктонные, бентосные (подробно), наземные, почвенные и др.
Планктонные формы представлены зелеными, золотистыми и желто-зелеными водорослями, имеющими специальные приспособления для облегчения переноса водой: уменьшающие плотность организмов (газовые вакуоли, включения липидов, студенистую консистенцию) и увеличивающие их поверхность (разветвленные выросты, приплюснутую или вытянутую форму тела и др.).
Бентосные формы обитают на дне водоемов или обволакивают находящиеся в воде предметы; к субстрату прикрепляются ризоидами, базальными дисками и присосками. В морях и океанах они представлены преимущественно бурыми и красными водорослями, а в пресных водоемах — всеми отделами водорослей, кроме Бурых. Бентосные водоросли содержат крупные хло-ропласты с высоким содержанием хлорофилла.
Наземные, или воздушные, водоросли (это обычно Зеленые или Желто-зеленые водоросли) образуют налеты и пленки различного цвета на коре деревьев, влажных камнях и скалах, заборах, крышах домов, на поверхности снега и льда и т.п. При недостатке влаги наземные водоросли пропитываются органическими и неорганическими веществами.
Почвенные водоросли (в основном Желто-зеленые, Золотистые и Диатомовые) живут в толще почвенного слоя на глубине до 1-2 м.
Особенности строения водорослей
Тело водорослей не разделено на вегетативные органы и представлено прочным в упругим талломом (слоевищем). Структура таллома — нитчатая (примеры: улотрикс, спирогира), пластинчатая {пример: ламинария), разветвленная или кустистая (пример: хара). Размеры — от 0,1 мм до нескольких десятков метров (у некоторых бурых и красных водорослей). Таллом разветвленных и кустистых водорослей — рассеченный и имеет линейно-членистое строение; в нем можно различить главную ось, «листья» и ризоиды.
У некоторых водорослей имеются специальные воздушные пузыри, которые удерживают слоевище у поверхности воды, где есть возможность максимального улавливания света для фотосинтеза.
Таллом многих водорослей выделяет слизь, которая заполняет их внутренние полости и частично выводится наружу, помогая лучше удерживать воду в препятствуя обезвоживанию.
Клетки таллома водорослей не дифференцированы и имеют проницаемою клеточную оболочку, внутренний слой которой состоит из целлюлозы, а наружный — из пектиновых веществ и (у многих видов) ряда добавочных компонентов: извести, лигнина, кутина (задерживающего ультрафиолетовые лучи и предохраняющего клетки от излишней потери воды в период отлива) и др. Оболочка выполняет защитную и опорную функции, обеспечивая при этом возможность роста. При дефиците влаги оболочки значительно утолщаются.
Цитоплазма клетки у большинства водорослей образует тонкий слой между большой центральной вакуолью и клеточной стенкой. В цитоплазме имеются органеллы: хроматофоры, эндоплазматический ретикулум, митохондрии, аппарат Гольджи, рибосомы, одно или несколько ядер.
Хроматофоры — это органеллы водорослей, содержащие фотосинтезирующие пигменты, рибосомы, ДНК, липидные гранулы и пиреноиды. В отличие от хлоропластов высших растений хроматофоры более разнообразны по форме (могут быть чашевидными, лентовидными, пластинчатыми, звездчатыми, дисковидными и др.), размерам, числу, строению, местоположению и набору фотосинтезирующих пигментов.
У мелководных (зеленых) водорослей фотосинтезирующими пигментами являются в основном хлорофиллы а и b, поглощающие красный и желтый свет. У бурых водорослей, обитающих на средних глубинах, куда проникает зеленый и синий свет, фотосинтезирующими пигментами являются хлорофиллы а и с, а также каротин и фукоксантин, имеющие бурый цвет. У красных водорослей, обитающих на глубинах до 270 м, фотосинтезирующими пигментами являются хлорофилл d (характерный только для этой группы растений) и имеющие красноватую окраску фикобилины — фикоэритрин, фикоцианин и аллофикоцианин, хорошо поглощающие синие и фиолетовые лучи.
Пиреноиды — особые включения, входящие в состав матрикса хроматофор и являющиеся зоной синтеза и накопления запасных питательных веществ.
Запасные вещества водорослей: крахмал, гликоген, масла, полисахариды и др.
Размножение водорослей
Водоросли размножаются бесполым и половым путем.
❖ Органы размножения водорослей (одноклеточные):
■ спорангии (органы бесполого размножения);
■ гаметангии (органы полового размножения).
❖ Способы бесполого размножения водорослей: вегетативное (фрагментами таллома) или одноклеточными зооспорами.
❖ Формы полового процесса у водорослей:
■ изогамия — слияние одинаковых по строению и величине подвижных гамет,
■ гетерогамия — слияние подвижных гамет разных размеров (более крупную считают женской),
■ оогамия — слияние крупной неподвижной яйцеклетки со сперматозоидом,
■ конъюгация — слияние содержимого двух неспециализированных клеток.
Половой процесс завершается образованием диплоидной зиготы, из которой формируется новая особь или образуются подвижные жгутиковые зооспоры, служащие для расселения водоросли.
❖ Особенности размножения водорослей:
■ у некоторых видов водорослей каждая особь способна формировать (в зависимости от времени года или условий среды) и споры, и гаметы;
■ у отдельных видов водорослей функции бесполого и полового размножения выполняют разные особи — спорофиты (они образуют споры) и гаметофиты (они формируют гаметы);
■ в цикле развития многих видов водорослей (красных, бурых, некоторых зеленых) наблюдается строгое чередование поколений — спорофита и гаметофита;
■ гаметы водорослей, как правило, обладают таксисами, определяющими направление их движения в зависимости от интенсивности света, температуры и т.п.;
■ безжгутиковые споры совершают амебоидное движение;
■ у морских водорослей выход спор или гамет совпадает с приливом; период покоя в развитии зиготы отсутствует (т.е. зигота начинает развиваться сразу же после оплодотворения, чтобы не быть унесенной в море).
Значение водорослей
❖ Значение водорослей:
■ они за счет фотосинтеза продуцируют органические вещества;
■ насыщают воду кислородом и поглощают из нее двуокись углерода;
■ являются пищей для водных животных;
■ являются родоначальниками растений, заселивших сушу;
■ участвовали в образовании горных известняковых и меловых пород, некоторых видов каменного угля и горючих сланцев;
■ зеленые водоросли очищают водоемы, загрязненные органическими отбросами;
■ используются человеком как органические удобрения и кормовые добавки в рацион животных;
■ используются в биохимической, пищевой и парфюмерной промышленности для получения белков, витаминов, спиртов, органических кислот, ацетона, йода, брома, агар-агара (необходим для изготовления мармелада, пастилы, суфле и т.п.), лаков, красителей, клея;
■ многие виды используются в пищу человеком (ламинария, некоторые зеленые и красные водоросли);
■ некоторые виды применяются при лечении рахита, зоба, желудочно-кишечных и других заболеваний;
■ ил из отмерших водорослей (сапропель) используется в грязелечении;
■ могут вызывать «цветение» воды.
Зеленые водоросли
❖ Спирогира
■ Местообитание: пресные стоячие и медленно текущие водоемы, где она образует тину ярко-зеленого цвета; распространена в Беларуси.
■ Форма тела: тонкая нитевидная; клетки расположены в один ряд.
■ Особенности строения клетки — цилиндрической формы с хорошо выраженной клеточной стенкой; покрыты пектиновой оболочкой и слизистым чехлом. Хроматофор лентовидный, спирально закрученный. Вакуоль занимает большую часть клетки. Ядро расположено в центре и соединяется тяжами с пристенной цитоплазмой; содержит гаплоидный набор хромосом.
■ Размножение: бесполое осуществляется путем разрыва нити на короткие участки; спорообразование отсутствует. Половой процесс — конъюгация. При этом две нити водорослей обычно располагаются параллельно друг другу и срастаются при помощи копуляционных выростов или мостиков. Затем оболочки клеток в местах соприкосновения нитей растворяются, образуя сквозной канал, через который содержимое одной из клеток перемещается в клетку другой нити и сливается с ее протопластом, образуя зиготу с плотной оболочкой. Зигота делится мей-озом; образуются 4 ядра, три из них погибают; из оставшейся клетки после периода покоя развивается взрослая особь.
❖ Улотрикс
■ Местообитание: пресные, реже морские и солоноватые водоемы, почва;
■ Жизненный цикл — сложный, с чередованием полового и бесполого поколений: многоклеточный гаметофит (половое поколение, формирующее гаметы) сменяется одноклеточным спорофитом (бесполым поколением, способным образовывать споры).
■ Форма тела гаметофита: нитевидная, неразветвленная; в водоемах формирует ярко-зеленые кустики высотой около 10 см.
■ Особенности строения гаметофита: клетки -цилиндрической формы, покрыты толстой целлюлозной оболочкой, расположены в один ряд. Нити таллома прикрепляются к субстрату бесцветной конической базальной клеткой, выполняющей функции ризоида. Все клетки, кроме базальной, способны делиться, обусловливая непрерывное нарастание таллома. Хроматофор имеет вид пластинки, образующей незамкнутый поясок или кольцо (цилиндр) вдоль клеточной стенки.
■ Строение спорофита: спорофит образован одной безжгутико-вой клеткой, которая представляет собой зиготу, окруженную плотной оболочкой и снабженную слизистой ножкой, которой спорофит прикрепляется к субстрату.
■ Размножение:
— бесполое — путем распада нити на короткие участки или путем образования жгутиковых зооспор (в одной клетке образуются 4 зооспоры);
половой процесс — изогамный; после оплодотворения зигота плавает, затем оседает на дно, теряет жгутики и образует спорофит. После периода покоя ядро спорофита делится, образуются зооспоры, которые затем прорастают.
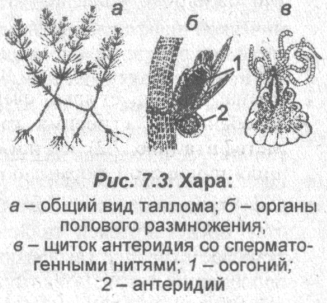
Харовые водоросли
Харовые водоросли (или харофиты) — высокоорганизованная группа пресноводных водорослей длиной от нескольких десятков сантиметров до нескольких метров, по внешнему виду похожих на высшие растения — хвощи.
Местообитание: чистые пресные водоемы с жесткой водой, насыщенной растворимыми солями кальция, на глубине 1-5 и более метров; образуют заросли на илистом или песчаном дне.
Особенности строения: тело мутовчато разветвленное, его главная ось («стебель») расчленена узлами на длинные (до 1 м) междоузлия. Каждое междоузлие образовано одной длинной многоядерной клеткой, часто покрытой коркой. От каждого узла отходит по четыре и более боковых побегов («листьев»). Органы прикрепления таллома к субстрату — нитевидные ризоиды. Хроматофоры содержат хлорофиллы а и b и каротиноиды. Запасный продукт — крахмал.
Размножение:
■ бесполое — клубеньками или участками таллома; споры не образуются;
■ половой процесс — оогамный.
Органы полового размножения — многоклеточные; мужские — антеридии (в которых развивается по несколько сотен двужгутиковых сперматозоидов) и женские — оогонии (в каждом из них образуется одна яйцеклетка). Сперматозоиды проникают к яйцеклетке через щель в оогонии. После оплодотворения развивается ооспора (зигота) с толстой оболочкой, которая прорастает после периода покоя.
Красные и бурые водоросли
Красные водоросли
■ Обитают в морях и океанах (иногда в пресных водоемах) на глубине более 200 метров.
■ Размеры таллома — от долей миллиметра до нескольких десятков сантиметров (большинство видов).
■ Форма таллома — нитчатая, кустистая, пластинчатая, корко- и коралловидная и др.
■ Цвет — от ярко-красного до голубовато-зеленого и желтого.
■ Структура таллома — нитчатая или пластинчатая.
■ Органы прикрепления — ризоиды, присоски, подошвы.
■ Хроматофоры пластинчатые, содержат хлорофиллы а и d и пигменты фикоэритрин (красный) и фикоцианин (синий).
■ Размножаются бесполым (спорами) и половым путем. В цикле развития большинства красных водорослей происходит чередование поколений — гаметофита и спорофита.
Бурые водоросли.
■ Это — наиболее высокоразвитые (по морфологической и анатомической дифференцировке таллома) среди всех групп низших растений; они, возможно, являются предками высших растений.
■ Обитают в морях и океанах (обычно на глубинах 40-100 м).
■ Размеры таллома — от нескольких сантиметров до 100 м и более.
■ Форма таллома — ветвисто-кустистая, пластинчатая или ленточная, расчлененная на стебле- и листовидные «органы».
■ Цвет -желтовато-бурый.
■ Структура таллома — пластинчатая или ленточная. У большинства представителей талломы имеют ложно- или истиннотканевое строение (выделяют ассимиляционные, запасающие, проводящие ткани). Некоторые представители имеют воздушные капсулы для поддержания вертикального положения в воде.
■ Органы прикрепления — ризоиды или базальный диск — дисковидное разрастание в основании таллома.
■ Таллом разрастается в результате вставочного роста или за счет деления верхушечных клеток.
■ Хроматофоры пластинчатые, содержат хлорофиллы о и с и пигменты каротин и фукоксантин (имеют бурый цвет).
■ Размножаются бесполым (спорами) и половым путем. Период покоя в развитии зиготы в новую особь отсутствует. В цикле развития большинства бурых водорослей происходит чередование поколений — гаметофита и спорофита.
Метки: растения
Источник: esculappro.ru
Споры бактерий
Некоторые бактерии образовывают споры, называемые эндоспорами, как средство борьбы с экстремальными условиями окружающей среде, которые угрожают их выживанию. Эти условия включают высокие температуры, засушливость, наличие токсичных ферментов или химических веществ и отсутствие пищи.
Спорообразующие бактерии развивают толстую клеточную стенку, которая является водонепроницаемой и защищает бактериальную ДНК от высыхания и повреждения. Эндоспоры могут выживать в течение длительных периодов времени, пока условия не изменятся и не станут пригодными для прорастания. Примеры бактерий, способных образовывать эндоспоры, включают клостридии (Clostridium) и бациллы (Bacillus).
Споры водорослей
Водоросли производят споры для бесполого размножения. Эти споры могут быть не подвижными (апланоспоры), или подвижными (зооспоры) и перемещаться из одного места в другое с помощью жгутиков. Некоторые водоросли способны размножаться половым или бесполым способом. Когда условия благоприятны, зрелые водоросли делятся и производят споры, которые развиваются в новых особей.
Споры гаплоидные и производятся митозом. Во времена, когда условия неблагоприятны для развития, водоросли подвергаются половому размножению и образовывают гаметы. Эти половые клетки сливаются, чтобы стать диплоидным зигоспорами. Зигоспора останется бездействующей до тех пор, пока условия не станут благоприятными еще раз. В это время зигоспора подвергается мейозу для создания гаплоидных спор.
Некоторые водоросли имеют жизненный цикл, который чередуется между различными периодами бесполого и полового размножения. Этот тип жизненного цикла называется чередованием поколений и состоит из гаплоидной и диплоидной фаз. В гаплоидной фазе структура, называемая гаметофитом, производит мужские и женские гаметы (половые клетки). При слиянии этих гамет образуется зигота. В диплоидной фазе зигота развивается в диплоидную структуру, называемую спорофитом. Спорофит продуцирует гаплоидные споры через мейоз.
Споры грибов
Большинство спор грибов выполняют две основных функции: перенесение неблагоприятных условий в состоянии покоя и размножение через рассеивание. Споры грибов могут быть одноклеточными или многоклеточными. Они бывают разных цветов, форм и размеров в зависимости от вида. Споры грибов бывают бесполыми или половыми:
- Бесполые споры, такие как спорангиоспоры, производятся и удерживаются внутри органов, называемых спорангиями. Другие бесполые споры, такие как конидии, образуются в результате митоза на нитчатых образованиях, называемых гифами.
- Половые споры включают аскоспоры, базидиоспоры и зигоспоры. Большинство грибов полагаются на ветер, чтобы рассеять споры в областях, где они могут успешно прорастать. Споры способны активно выбрасываться из репродуктивных структур (баллистоспор) или могут быть высвобождены без активного выброса (статисмоспор). Споры переносятся ветром в другие места. Чередование поколений распространено среди грибов. Иногда условия окружающей среды таковы, что необходимо, чтобы грибковые споры были бездействующими. Прорастание после периодов покоя у некоторых грибов может быть вызвано разными факторами, включая температуру, уровень влажности и количество других спор в определенной области. Покой позволяет грибам выжить в стрессовых условиях.
Споры растений
Как водоросли и грибы, растения также демонстрируют чередование поколений. Растения без семян, такие как папоротники и мхи, развиваются из спор. Споры производятся внутри спорангий и выпускаются в окружающую среду. Первичной фазой жизненного цикла несосудистых растений, таких как мхи, является генерация гаметофитов (половая фаза).
Фаза гаметофита состоит из зеленой мшистой растительности, а фаза спорофита (бесполая фаза) состоит из удлиненных стеблей со спорами, заключенными внутри спорангий, расположенных на кончике стеблей. У сосудистых растений, которые не производят семена, такие как папоротники, поколения спорофит и гаметофит являются независимыми. Лист папоротника представляет собой зрелый диплоидный спорофит, тогда как спорангии на нижней стороне листьев образовывают споры, которые развиваются в гаплоидный гаметофит.
У цветковых растений (покрытосеменных) и нецветковых семенных растений генерация гаметофитов полностью зависит от доминантного спорофита. У покрытосеменных, цветок вырабатывает как мужские микроспоры, так и женские мегаспоры. Микроспоры содержатся в пыльце, а мегаспоры производятся внутри цветочной завязи. При опылении микроспоры и мегаспоры объединяются, образуя семена.
Источник: natworld.info