Начальная стадия фотосинтеза
 История открытия удивительного и такого жизненного важного явления, как фотосинтез уходит корнями глубоко в прошлое. Более четырех веков назад в 1600 году бельгийский ученый Ян Ван – Гельмонт поставил простейший эксперимент. Он поместил веточку ивы в мешок, где находилось 80 кг земли. Ученый зафиксировал первоначальный вес ивы, и затем на протяжении пяти лет поливал растение исключительно дождевой водой. Каково же было удивление Яна Ван – Гельмонта, когда он повторно взвесил иву. Вес растения увеличился на 65 кг, причем масса земли уменьшился всего на 50 гр! Откуда растение взяло 64 кг 950 гр питательных веществ для ученого осталось загадкой!
История открытия удивительного и такого жизненного важного явления, как фотосинтез уходит корнями глубоко в прошлое. Более четырех веков назад в 1600 году бельгийский ученый Ян Ван – Гельмонт поставил простейший эксперимент. Он поместил веточку ивы в мешок, где находилось 80 кг земли. Ученый зафиксировал первоначальный вес ивы, и затем на протяжении пяти лет поливал растение исключительно дождевой водой. Каково же было удивление Яна Ван – Гельмонта, когда он повторно взвесил иву. Вес растения увеличился на 65 кг, причем масса земли уменьшился всего на 50 гр! Откуда растение взяло 64 кг 950 гр питательных веществ для ученого осталось загадкой!
Следующий значимый эксперимент на пути открытия фотосинтеза принадлежал английскому химику Джозефу Пристли. Ученый посадил под колпак мышь, и через пять часов грызун умер.
гда же Пристли поместил с мышью веточку мяты и также накрыл грызуна колпаком, мышь осталась живой. Этот эксперимент навел ученого на мысль о том, что существует процесс, противоположный дыханию. Ян Ингенхауз в 1779 году установил тот факт, что только зеленые части растений способны выделять кислород. Через три года швейцарский ученый Жан Сенебье доказал, что углекислый газ, под воздействием солнечных лучей, разлагается в зеленых органоидах растений. Спустя всего пять лет французский ученый Жак Буссенго, проводя лабораторные исследования, обнаружил тот факт, что поглощение растениями воды также происходит и при синтезе органических веществ. Эпохальное открытие в 1864 году совершил немецкий ботаник Юлиус Сакс. Ему удалось доказать, что объем потребляемого углекислого газа и выделяемого кислорода происходит в пропорции1:1.
Фотосинтез – один из самых значимых биологических процессов
Говоря научным языком, фотосинтез (от др.-греч. φῶς — свет и σύνθεσις — соединение, связывание) — это процесс, при котором из углекислого газа и воды на свету образуются органические вещества. Заглавная роль в этом процессе принадлежит фотосинтетическим сегментам.
Если говорить образно, то лист растения можно сравнить лабораторией, окна которой выходят на солнечную сторону. Именно в ней происходит образование органических веществ. Этот процесс является основой существования всего живого на Земле.
Многие резонно зададут вопрос: чем дышат люди, живущие в городе, где не то что дерева, и травинки днем с огнем не сыщешь. Ответ очень прост. Дело в том, что на долю наземных растений приходится всего 20% выделяемого растениями кислорода. Главенствующую роль в выработке кислорода в атмосферу играют морские водоросли. На их долю приходится 80% от вырабатываемого кислорода. Говоря языком цифр, и растения, и водоросли ежегодно выделяют в атмосферу 145 млрд. тонн (!) кислорода! Недаром мировой океан называют «легкими планеты».
Общая формула фотосинтеза выглядит следующим образом:
Вода + Углекислый газ + Свет → Углеводы + Кислород
Для чего нужен фотосинтез растениям?
Как мы уяснили, фотосинтез – это необходимое условие существования человека на Земле. Однако это не единственная причина, по которой фотосинтезирующие организмы производят активную выработку кислорода в атмосферу. Дело в том, что и водоросли, и растения ежегодно образуют более 100 млрд. органических веществ (!), которые составляют основу их жизнедеятельности. Вспоминая эксперимент Яна Ван-Гельмонта мы понимаем, что фотосинтез – это основа питания растений. Научно доказано, что 95% урожая определяют органические вещества, полученные растением в процессе фотосинтеза, и 5% – те минеральные удобрения, которые садовод вносит в почву.
Современные дачники основное внимание уделяют почвенному питанию растений, забывая о его воздушном питании. Неизвестно, какой урожай могли бы получить садоводы, если бы они внимательно относились к процессу фотосинтеза.
Однако ни растения, ни водоросли не могли бы так активно производить кислород и углеводы, не будь у них удивительного зеленого пигмента – хлорофилла.
Тайна зеленого пигмента
Главное отличие клеток растения от клеток иных живых организмов – это наличие хлорофилла. К слову сказать, именно он является виновником того, что листья растений окрашены именно в зеленый цвет. Это сложное органическое соединение обладает одним удивительным свойством: оно способно поглощать солнечный свет! Благодаря хлорофиллу становится возможны и процесс фотосинтеза.
Две стадии фотосинтеза
Говоря простым языком, фотосинтез представляет собой процесс, при котором поглощенные растением вода и углекислый газ на свету при помощи хлорофилла образуют сахар и кислород. Таким образом, неорганические вещества удивительным образом превращаются в органические. Полученный в результате преобразования сахар является источником энергии растений.
Фотосинтез имеет две стадии: световую и темновую.
Световая фаза фотосинтеза
Осуществляется на мембранах тилакойдов.
Тилакойд – это структуры, ограниченные мембраной. Они располагаются в строме хлоропласта.
Порядок событий световой стадии фотосинтеза:
- На молекулу хлорофилла попадает свет, который затем поглощается зеленым пигментом и приводит его в возбужденное состояние. Входящий в состав молекулы электрон переходит на более высокий уровень, участвует в процессе синтеза.
- Происходит расщепление воды, в ходе которого протоны под воздействием электронов превращаются в атомы водорода. Впоследствии они расходуются на синтез углеводов.
- На завершающем этапе световой стадии происходит синтез АТФ (Аденозинтрифосфат). Это органическое вещество, которое играет роль универсального аккумулятора энергии в биологических системах.
Темновая фаза фотосинтеза
Местом протекания темновой фазы являются строму хлоропластов. Именно в ходе темновой фазы происходит выделение кислорода и синтез глюкозы. Многие подумают, что такое название эта фаза получила потому что процесс, происходящие в рамках этого этапа осуществляются исключительно в ночное время. На самом деле, это не совсем верно. Синтез глюкозы происходит круглосуточно. Дело в том, что именно на данном этапе световая энергия больше не расходуется, а значит, она попросту не нужна.
Значение фотосинтеза для растений
Мы уже определили тот факт, что фотоинтез нужен растениям ничем не меньше, чем нам. О масштабах фотосинтеза очень просто говорить языком цифр. Ученые рассчитали, что только растения суши запасают столько солнечной энергии, сколько могли бы израсходовать 100 мегаполисов в течение 100 лет!
Дыхание растений – это процесс, противоположный фотосинтезу. Смысл дыхания растений заключается в освобождении энергии в процессе фотосинтеза и направление ее на нужды растений. Говоря простым языком, урожай – это разница между фотосинтезом и дыханием. Чем больше фотосинтез и ниже дыхание, тем больше урожай, и наоборот!
Фотосинтез – это удивительный процесс, который делает возможной жизнь на Земле!
Источник: xn—-8sbiecm6bhdx8i.xn--p1ai
Фазы фотосинтеза
Он происходит в две фазы:
Световая фаза (фотофосфорилирование) – представляет собой набор светозависимых фотохимических (т. е. светозахватывающих) реакций, в которых электроны транспортируются через обе фотосистемы (PSI и PSII) для получения АТФ (богатая энергией молекула) и NADPHH (восстанавливающий потенциал).
Таким образом, светлая фаза фотосинтеза позволяет непосредственно превращать световую энергию в химическую энергию. Именно через этот процесс наша планета теперь имеет атмосферу, богатую кислородом. В результате высшие растения сумели доминировать на поверхности Земли, обеспечивая пищу многим другим организмам, которые питаются или находят убежище через неё. Первоначальная атмосфера содержала такие газы, как аммоний, азот и углекислый газ, но очень мало кислорода. Растения нашли способ превратить этот CO настолько обильно в пищу, используя солнечный свет.
Темновая фаза – соответствует полностью ферментативному и не зависящему от света циклу Кальвина, в котором аденозинтрифосфат (АТФ) и НАДФН+Н+ (никотин амид адениндинуклеотидфосфат) используются для конверсии углекислого газа и воды в углеводы. Эта вторая фаза позволяет усвоить углекислый газ.
То есть в этой фазе фотосинтеза, примерно через пятнадцать секунд после поглощения CO происходит реакция синтеза и появляются первые продукты фотосинтеза — сахара: триосы, пентозы, гексозы, гептозы. Из определённых гексоз образуются сахароза и крахмал. Помимо углеводов, могут также развиваться липидами и белками путём связывания с молекулой азота.
Этот цикл существует в водорослях, умеренных растениях и всех деревьях; эти растения называются «растениями С3», наиболее важными промежуточными телами биохимического цикла, имеющими молекулу три атома углерода (С3).
В этой фазе хлорофилл после поглощения фотона имеет энергию 41 ккал на моль, некоторые из которых преобразуются в теплоту или флуоресценцию. Использование изотопных маркеров (18O) показало, что кислород, высвобождаемый во время этого процесса, происходит из разложенной воды, а не из поглощённого диоксида углерода.
Как происходит фотосинтез
Фотосинтез происходит главным образом в листьях растений и редко (когда-либо) в стеблях и т. д. Части типичного листа включают: верхний и нижний эпидермис;
- мезофилл;
- сосудистый пучок (вены);
- устьица.
Если клетки верхнего и нижнего эпидермиса не являются хлоропластами, фотосинтез не происходит. Фактически они служат прежде всего в качестве защиты для остальной части листа.
Устьица — это дыры, существующие главным образом в нижнем эпидермисе, и позволяют проводить обмен воздуха (CO и O2). Сосудистые пучки (или вены) в листе составляют часть транспортной системы растения, при необходимости перемещая воду и питательные вещества вокруг растения. Клетки мезофилла имеют хлоропласты, вот это и есть место фотосинтеза.
Механизм фотосинтеза очень сложный. Однако эти процессы в биологии имеют особое значение. При энергичном воздействии света хлоропласты (части растительной клетки, содержащие хлорофилл), вступая в реакцию фотосинтеза, объединяют углекислый газ (СО) с пресной водой с образованием сахаров C6H12O6.
Они в процессе реакции превращаются в крахмал C6H12O5, для квадратного дециметра поверхности листа, в среднем 0,2 г крахмала в день. Вся операция сопровождается сильным высвобождением кислорода.
Фактически процесс фотосинтеза состоит в основном из фотолиза молекулы воды.
Формула этого процесса:
6 Н 2 О + 6 СО 2 + свет = 6 O 2 + С 6 Н 12 О 6
Вода + углекислый газ + свет = кислород + глюкоза
- Н 2 О = вода
- СО 2 = диоксид углерода
- O 2 = Кислород
- С 6 Н 12 О 6 = глюкоза
В переводе этот процесс означает: растению для вступления в реакцию нужны шесть молекул воды + шесть молекул углекислого газа и света. Это приводит к образованию шести молекул кислорода и глюкозы в химическом процессе. Глюкоза — это глюкоза, которую растение использует в качестве исходного материала для синтеза жиров и белков. Шесть молекул кислорода являются всего лишь «необходимым злом» для растения, которое он доставляет в окружающую среду через закрывающие клетки.
Основные продукты фотосинтеза
Как уже было сказано, углеводы являются наиболее важным прямым органическим продуктом фотосинтеза в большинстве зелёных растений. В растениях образуется мало свободной глюкозы; вместо этого глюкозные единицы связаны с образованием крахмала или соединены с фруктозой, другим сахаром, с образованием сахарозы.
При фотосинтезе синтезируются не только углеводы, как это когда-то считалось, но также:
- аминокислоты;
- белки;
- липиды (или жиры);
- пигменты и другие органические компоненты зелёных тканей.
Минералы поставляют элементы (например, азот, N; фосфор, Р; серы, S), необходимых для образования этих соединений.
Химические связи разрушаются между кислородом (O) и углеродом (С), водородом (Н), азотом и серы, а новые соединения образуются в продуктах, которые включают газообразный кислород (O 2) и органические соединения. Для разрушения связей между кислородом и другими элементами (например, в воде, нитрате и сульфате) требуется больше энергии, чем высвобождается, когда в продуктах образуются новые связи. Это различие в энергии связи объясняет большую часть световой энергии, хранящейся в виде химической энергии в органических продуктах, образующихся при фотосинтезе. Дополнительная энергия хранится при создании сложных молекул из простых.
Факторы, влияющие на скорость фотосинтеза
Скорость фотосинтеза определяется в зависимости от скорости производства кислорода либо на единицу массы (или площади) зелёных растительных тканей, либо на единицу веса всего хлорофилла.
Количество света, подача углекислого газа, температура, водоснабжение и наличие полезных ископаемых являются наиболее важными факторами окружающей среды, которые влияют на скорость реакции фотосинтеза на наземных установках. Его скорость определяется также видами растений и его физиологическим состоянием, например, его здоровьем, зрелостью и цветением.
Место фотосинтеза
Фотосинтез происходит исключительно в хлоропластах (греческий хлор = зелёный, пластообразный) растения. Хлоропласты преимущественно обнаруживаются в палисадах, но также и в губчатой ткани. На нижней стороне листа находятся блокирующие ячейки, которые координируют обмен газами. CO 2 течёт в межклеточные клетки снаружи.
Вода, необходимая для фотосинтеза, транспортирует растение изнутри через ксилему в клетки. Зелёный хлорофилл обеспечивает поглощение солнечного света. После того как углекислый газ и вода превращаются в кислород и глюкозу, закрывающие клетки открывают и выделяют кислород в окружающую среду. Глюкоза остаётся в клетке и превращается растением среди других в крахмал. Сила сравниваются с полисахаридом глюкозы и лишь слегка растворимой, так что даже в высоких потерях воды в прочности растительных остатков.
Важность фотосинтеза в биологии
Из света, полученного листом, отражается 20%, 10% передаются и 70% фактически поглощаются, из которых 20% рассеивается в тепле, 48% теряется при флуоресценции. Около 2% остаётся для фотосинтеза.
Благодаря этому процессу растения играют незаменимую роль на поверхности Земли; на самом деле зелёные растения с некоторыми группами бактерий являются единственными живыми существами, способными выработать органические вещества из минеральных элементов. По оценкам, каждый год 20 миллиардов тонн углерода фиксируются наземными растениями из углекислого газа в атмосфере и 15 миллиардов водорослями.
Зелёные растения являются основными первичными производителями, первое звено в пищевой цепи; не хлорофилловые растения и травоядные и плотоядные животные (включая людей) полностью зависят от реакции фотосинтеза.
Упрощённое определение фотосинтеза заключается в том, чтобы преобразовать световую энергию от солнца в химическую энергию. Этот фотонный биосинтез углевода производится из углекислого газа СО2 с помощью световой энергии.
То есть фотосинтез является результатом химической активности (синтеза) растений хлорофилла, которые продуцируют основные биохимические органические вещества из воды и минеральных солей благодаря способности хлоропластов захватывать часть энергии солнца.
Источник: obrazovanie.guru
-
Фотосинтез. Общая характеристика основных этапов фотосинтеза: фотофизического, фотохимического, фотоэнзиматического и энзиматического. Разнообразие биосинтетических процессов темновой стадии фотосинтеза: цикл Кальвина, циклы Карпилова-Хэтча-Слэка, цикл гликолевой кислоты
Фотосинтез — совокупность физико-химических процессов, в ходе которых углекислый газ восстанавливается до углеводов, а энергия света трансформируется в энергию химических связей органических молекул. Суммарное уравнение фото-за: 6CO2+ 6H2O=(hню, т.е. на свету в хлоропласте) C6H12O6 + 6O2.Фотосинтез делят на 2 фазы: световую и темновую. Световая – сов-ть процессов ФС, требующие для их протекания свет, темновая не требует свет. Световая фаза протекает на тилакоидах, темновая – в строме хлоропласта.
Этапы ФС:
-
Фотофизический этап, самый быстрый, идет в пигментах. В ходе него Е света превращается в Е электронного возбуждения молекулы пигмента. Основные пигменты сосредоточены в системе ССК – свето-собирающего комплекса. Электрон получает доп. Энергию и переходит в возбужденное состояние.
-
Фотохимический этап протекает в реакционных центрах фотосистемы (ФС). В ходе этого этапа Е электронного возбуждения трансформируется в электрическую (кулоновскую) Е. Каждая ФС (их всего две) содержит 3 компонента: первичный донор (D), первичный акцептор (А) и пигмент-ловушку (Р). ФС1:D– пластоцианин( синий белок с Сu),A– Хлорофилл «а» 695, не требующий света, Р – димер Хлорофилла «а» Р700. ФС2:D–Mn-кластер, А –Pheo(феофетин) и Р – Р680.В результате фотохимических реакций в хлоропластах создается необходимый уровень АТР и NADPH, которые необходимы для функционирования темновой стадии фотосинтеза, где СО2восстанавливается до углевода.
-
Фотоэнзимотический этап — это совокупность биохимических реакций, в результате которых происходит усвоение растениями углекислого газа атмосферы (С02) и образование углеводов, протекает в строме. При нециклическом потоке электронов НАДФ восстан-ся до НАДФН и происходит Образование АТФ. Суммарное уравнение нециклич-го фосфор-ия: АДФ + Фн + НАДФ+ 2Н2О= АТФ+НАДФН+Н, накапливаются световые прод-ты АТФ и НАДФН. АДФ+Фн=АТФ+Н2О.
Нециклический и циклический транспорт электронив (z-схема):В ФС11 димер П680, поглотив энергию 2 квантов переходит в возбужденное сосотояние и отдает 2 электрона Фф. От Фф электроны передаются на пластохиноны QA, QB , затем на пул липидорастворимых молекул PQ, которые переносят электроны и протоны через липидную фазу мембраны на железосерный белок Риске FeSR и цитохром f (гемопротеин) цитохромного комплекса b6-f, восстанавливая Сu-содержащий белок пластоцианин (Пц). Вакантные места («дырки») в П680 заполняются двумя электронами из содержащего марганец переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду (фотолиз) и восстанавливается за счет электронов воды. Для осуществления этих реакций в белковом комплексе S необходимы Mn и Cl—. При возбуждении П700 в реакционном центре ФС1 2 электрона захватываются мономерной формой хлорофилла а (А1) и передаются последовательно на А2, Ав и Фд, с которого электроны с помощью NADP-оксидоредуктазы с FAD в качестве кофактора идут на восстановление NADP+.
На вакантные места в П700 переходят электроны с Пц и нециклическая цепь переноса электронов таким образом замыкается.
Наряду с нециклическим в мембранах хлоропластов функционирует циклический транспорт электронов, включающий в себя только ФС1 и комплекс цитохромов b6-f. В этом случае NADP+ не восстанавливается и освобождающаяся энергия используется для фосфорилирования АDP.
-
Энзиматический этап идет в строме, переводит трансформирование световых продуктов в энергию хим. связей темновых продуктов.
Цикл Кальвина – основной биохимический процесс фотосинтеза. 3 этапа цикла:
Первая фаза — карбоксилирование. Эта реакция катализируется специфическим для процесса фотосинтеза ферментом рибулозобисфосфат-карбоксилазой/ оксигеназой (сокращенно Rubisco). Особенностью фермента является то, что катализируемая им реакция является самой медленной стадией в цикле фиксации углекислоты При взаимодействии РБФ с С02 образуется сначала промежуточное нестойкое шестиуглеродное соединение, которое затем распадается на две молекулы ФГК. Образовавшаяся ФГК — это органическая кислота, и ее энергетический уровень ниже уровня Сахаров. Поэтому это соединение не может непосредственно превращаться в углеводы. Необходимо превращение его в трехуглеродный сахар — фосфоглицериновый альдегид (ФГА).
Вторая фаза — восстановление.Дальнейшие превращения ФГК требуют участия продуктов световой фазы фотосинтеза: АТФ и НАДФН + Н+. Реакция идет в два этапа. Прежде всего, происходит реакция фосфорилирования 3-ФГК. Донором фосфатной группы является АТФ. АТФ требуется здесь в качестве дополнительного источника энергии. Образуется 1,3-дифосфоглицериновая кислота. Образовавшийся ФГА является по уровню восстановленным углеводом. Это соединение вступает в две последние фазы. Пять молекул ФГА используется на регенерацию акцептора РБФ для того, чтобы фиксация С02 могла снова осуществляться. Оставшаяся шестая молекула вступает в фазу «синтеза продуктов», где превращается в более сложные соединения (углеводы, аминокислоты и др.).
Третья фаза — регенерация.В процессе регенерации акцептора используется пять молекул ФГА, в результате чего образуются три молекулы рибулезо-5-фос-фата. Этот процесс идет через образование 4-, 5-, 6-, 7-углеродных соединений. Образовавшийся триозофосфат (ФГА) вступает в четвертую стадию темновых реакций — стадию образования продуктов фотосинтеза.
С4-путь фотосинтеза (Цикл Хетча и Слэка).
К группе растений с С4-путем фотосинтеза относятся сахарный тростник, кукуруза, сорго и др.Листья этих растений содержат два разных типа хлоропластов: хлоропласты обычного вида – в клетках мезофилла и большое количество крупных хлоропластов в клетка, окружающих проводящие пучки (обкладка).
СО2 попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с фосфоенолпируватом (ФЕП), образуя щавелевоуксусную кислоту (оксалоацетат). Затем уже в хлоропластах оксалоацетат восстанавливается до яблочной кислоты (малата) за счет NADPH, образующегося в ходе световой стадии фотосинтеза. Затем малат переносится в хлоропласты клетки обкладки сосудистого пучка, где он декарбоксилируется малаьдегидрогеназой декарбоксилирующей до пирувата и СО2. СО2вступает в цикл Кальвина, а пируват возвращается в хлоропласты клеток мезофилла. Такая компартментализация процесс фиксации и использования СО2 позволяет растениям с С4-путем осуществлять фотосинтез даже при закрытых устьицах (в засушливы или засоленных местах обитания), так как хлоропласты клеток обкладки используют малат, образовавшийся ранее, как донор СО2.С4-растения могут также использовать СО2, возникающий при фотодыхании.
САМ-метаболизм (по типу толстянковых)
У суккулентов наблюдается суточный цикл метаболизма С4-кислот с образованием яблочной кислоты ночью. Этот тип фотосинтеза называют САМ-метаболизм.Устьица этих растений днем обычно закрыты, что предотвращает потерю воды, и открываются ночью. СО2 поступает в листья, где взаимодействует с ФЕП, образуя ЩУК (оксалоацетат), который восстанавливается до яблочной кислоты. Она накапливается в вакуолях клеток листа, что приводит к закислению клеточного сока в ночное время.
Днем в условиях высокой температуры, когда устьица закыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии малатдегидрогеназы декарбоксилирующей (малик-энзима) с образованием СО2 и пирувата. СО2 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
Фотодыхание –это активируемый светом процесс высвобождения СО2 и поглощения О2, т.е. процесс обратный фотосинтезу. Этот процесс усиливается при низком содержании СО2 и высоких концентрациях О2 или при повышении температуры. В этих условиях РДФ-карбоксилаза в хлоропластах может функционировать как оксигеназа, катализируя окислительное расщепление Рибулозо-1,5-дифосфата на 3-ФГК и 2-фосфогликолевую кислоту, которая дефосфорилируется в гликолевую кислоту.
Гликолат из хлоропластов поступает в пероксисому и там окисляется до глиоксилата. Возникающая перекись водорода устраняется каталазой. Глиоксилат аминируется, превращаясь в глицин и транспортируется в митохондрию.
В митохондрии из двух молекул глицина образуется серин и освобождается СО2. Серин может снова поступать в пероксисому, где трансаминируется с пируватом. Образовавшийся гидроксипируват восстанавливается до глицерата, который может вновь поступать в хлоропласты и включаться в цикл Кальвина. У С4-растений высвобождающийся при фотодыхании СО2 перехватывается в клетках мезофилла, где из него образуются оксалоацетат и малат.
Значение цикла гликолевой кислоты: фотосинтез неуглеводных продуктов; защита хлоропластов от высокой концентрации кислорода, что особенно актуально при избытке света. При блокировании фотодыхания С3-растения «выгорают» на свету.
Источник: StudFiles.net
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
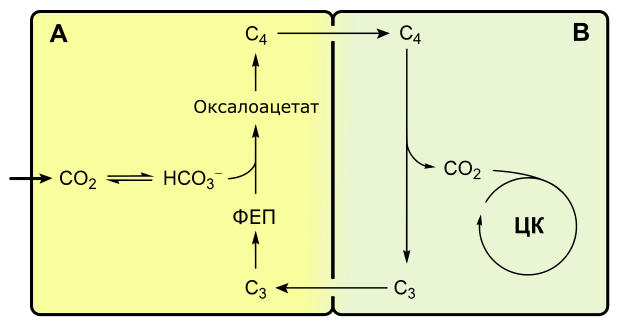
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su