В результате темновой фазы фотосинтеза образуется
Фотосинтез у зеленых растений — это процесс преобразования света в химическую энергию органических соединений, синтезируемых из диоксида углерода и воды. Процесс фотосинтеза представляет собой цепь окислительно-восстановительных реакций, совокупность которых принято подразделять на две фазы — световую и темновую.
Во время световой фазы фотосинтеза энергия солнечной радиации, поглощенная пигментными системами хлоропластов, преобразуется в электрохимическую. Преобразование осуществлястся путем переноса электронов и ионов водорода с помощью специальных переносчиков через мембрану тилакоидов. Такой перенос ионов Н* и электронов получил название химиоосмоса. Этот процесс выяснен еще не до конца, но в общих чертах его можно представить следующим образом.
При попадании кванта света на молекулу хлорофилла один из его электронов переходит на более высокий энергетический уровень, т.
оказывается в возбужденном состоянии. Возбужденный электрон может вернуться в основное состояние, и в этом случае его избыточная энергия выделяется в виде флуоресценции (красное свечение) или тепла, или же он передается в качестве энергии возбуждения другим молекулам. Кроме того, электрон может отрываться от молекулы хлорофилла. В последнем случае электроны, обладающие запасом энергии, захватываются переносчиками, встроенными в мембрану, и парами переносятся на внешнюю сторону мембраны тилакоида.
Здесь электроны акцептируются коферментом НАДФ (никотинамидадениндинуклеотидфосфат), к которому присоединяется два протона из стромы и образуется НАДФ-восстановленный НАДФ ∙ H + H+:
НАДФ+ + 2e— + 2Н+ → НАДФ ∙ H + H—.
Связывание протонов приводит к формированию отрицательного поля вокруг тилакоида.
Молекулы хлорофилла, утратившие электроны, являются сильными окислителями и заполняют «электронные дырки» электронами из молекул воды, находящихся внутри тилакоидов. Молекулы воды при этом разрушаются:
2H2O – 4e— → 4H+ + O2.
Этот процесс называется фотолизом, или фотоокислением воды. Благодаря фотолизу внутри тилакоида накапливаются положительно заряженные протоны Н+ и образуется молекулярный кислород, который диффундирует в атмосферу.
Увеличение концентрации протонов внутри тилакоида осуществляется также благодаря активному закачиванию их из стромы, что сопряжено с транспортом электронов.
Таким образом, в результате поглощения хлорофиллом световой энергии и вызванного ею транспорта электронов на внутренней стороне мембраны тилакоидов создается электрохимический потенциал водорода (ΔμН+), имеющий две составляющие: концентрационную (ΔрН+), возникающую в результате неравномерного распределения Н+ по разные стороны мембраны, и электрохимическую, обусловленную противоположными зарядами разных сторон мембраны тилакоида.
По мере накопления протонов на внутренней стороне тилакоида нарастает разность потенциалов и при достижении критической величины (150 мв) протоны начинают двигаться в строму через каналы фермента АТФ-синтетазы, встроенного в мембрану тилакоида. Энергия перехода Н+ по протонному каналу используется для фосфорилирования имеющихся в матриксе молекул АДФ:
АДФ + Фн → АТФ.
Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации СO2.
Таким образом, в результате переноса электронов и протонов через мембрану тилакоида происходит превращение световой энергии в химическую энергию макроэргических связей молекулы АТФ, а также образование сильного восстановителя НАДФ ∙ Н + Н+ и выделение свободного кислорода. Кислород, образующийся при фотолизе воды, является побочным продуктом фотосинтеза. Он может использоваться дальше растительными клетками для дыхание или выделяться в биосферу.
Темновая фаза осуществляется в строме хлоропластов без непосредственного участия света. Это восстановление СO2 до уровня органических веществ за счет использования энергии АТФ и НАДФ ∙ Н + Н+, синтезированных во время световой фазы. Восстановление молекул СO2 начинается с их фиксации молекулами пятиуглеродного сахара рибулозодифосфата. При взаимодействии рибулозодифосфата и СO2 образуется сначала нестойкое шестиуглеродное соединение, которое затем ферментативным путем распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК). Дальнейшее превращение ФГК требует участия продуктов световой фазы фотосинтеза — АТФ и НАДФ ∙ Н + Н+. При восстановлении фосфоглицериновой кислоты образуется фосфоглицериновый альдегид (триозофосфат) — первый сахар. В конечном итоге через ряд промежуточных соединений образуются шестиуглеродные сахара, а затем другие органические вещества (амино- и органические кислоты, нуклеотиды, спирты, в том числе глицерол и др.).
2C3 + АТФ + НАДФ ∙ H + H+ → 2C3 + АДФ + НАДФ + H3PO4.
Следует учесть, что в этих реакциях одновременно участвуют многие однотипные молекулы. На определенном этапе судьба трехуглеродных молекул ФГА может оказаться различной. Одни из них соединяются друг с другом и образуют шестиуглеродные сахара (C6H12O6), которые, в свою очередь, могут полимеризоваться в крахмал, целлюлозу и другие макромолекулы или использоваться на энергетические нужды клетки. Другие молекулы ФГА идут на синтез аминокислот путем присоединения аминогрупп или на синтез карбоновых кислот, спиртов и т. д. Наконец, третьи вовлекаются в длинный ряд реакций, которые приводят к превращению трехуглеродных молекул в молекулы исходного пятиуглеродного сахара — рибулозодифосфата, которые снова могут акцептировать диоксид углерода. Поскольку часть трехуглеродных конечных продуктов превращается в новые молекулы рибулозодифосфата, процесс фиксации углерода, по существу представляет собой цикл. Его называют C3-циклом (по C3-продуктам) или циклом Кальвина, в честь ученого, открывшего этот процесс. Следует также отметить, что неорганические соединения, используемые в цикле Кальвина, поглощаются корнями растений в виде нитратов, фосфатов и сульфатов из почвы.
В темновой фазе фотосинтеза энергия макроэргических связей АТФ преобразуется в химическую энергию органических веществ, т. е. энергия как бы консервируется в химических связях между атомами органических соединений.
Если объединить реакции световой и темновой фазы, исключив все промежуточные этапы, то получается суммарное уравнение процесса фотосинтеза:
6CO2 + 6H2O → C6H12O6 + 6O2.
В изучение процесса фотосинтеза, раскрытие его механизма большой вклад внесли русский ученый К.А.Тимирязев, американцы М.Кальвин и Д.Арнон, австралийцы М.Д.Хетч и К.Р.Слэйк.
Источник: jbio.ru
Фотосинтез — уникальная система процессов создания с помощью хлорофилла и энергии света органических веществ из неорганических и выделения кислорода в атмосферу, реализуемая в огромных масштабах на суше и в воде.
Фотосинтез происходит в клетках зелёных растений с помощью пигментов, главным образом хлорофилла, находящегося в хлоропластах клетки. Его продуктами являются мономеры углеводов (моносахариды: глюкоза, фруктоза и др.).
В основе фотосинтеза лежит окислительно-восстановительный процесс, в котором электроны переносятся от донора-восстановителя (вода, водород и др.) к акцептору (CO2 , ацетат и др.) с образованием восстановленных соединений (углевода) и выделением кислорода, если окисляется вода. Фотосинтезирующие бактерии часто используют другие доноры, а не воду, кислород при этом они не выделяют.
В системе процессов фотосинтеза различают два цикла реакций, как две фазы, последовательно и непрерывно идущие друг за другом — световую и темновую (рис. 62).
Световая фаза фотосинтеза характеризуется тем, что здесь все процессы происходят только при участии энергии света, поэтому её и называют световой. Связывание солнечной (электромагнитной) энергии происходит преимущественно на мембранах тилакоидов хлоропласта. Размещающийся здесь хлорофилл и другие пигменты собраны в функциональные единицы-комплексы — пигментные системы, получившие название фотосистемы.
 |
| Рис. 62. Схема фотосинтеза |
Таким образом, светособирающие и пигментно-белковые комплексы фотосистемы I и фотосистемы II обеспечивают процесс фотосинтеза необходимой энергией в виде макроэнергетических соединений НАДФ•Н и АТФ. В этом заключается основная функция световой фазы фотосинтеза. Она реализуется только при участии света и с помощью пигментов, размещённых в тилакоидной мембране хлоропластов.
Темновая фаза фотосинтеза проходит в строме хлоропласта без непосредственного поглощения света, в любое время суток. В процессе световой фазы фотосинтеза накапливается достаточно высокий уровень АТФ и НАДФ•Н. Однако сами по себе эти высокоэнергетические соединения не способны синтезировать углеводы из CO2. Поэтому становится очевидным, что и темновая фаза фотосинтеза — сложный процесс, включающий большое количество реакций. Материал с сайта http://doklad-referat.ru
Все процессы темновой фазы фотосинтеза идут без непосредственного потребления света, но в них большую роль играют высокоэнергетические вещества (АТФ и НАДФ•Н), образующиеся с участием энергии света, во время световой фазы фотосинтеза. В процессе темновой фазы энергия макроэнергетических связей АТФ преобразуется в химическую энергию органических соединений молекул углеводов. Это значит, что энергия солнечного света как бы консервируется в химических связях между атомами органических веществ, что имеет огромное значение в энергетике биосферы и конкретно для жизнедеятельности всего живого населения нашей планеты.
Фотосинтез происходит в хлоропластах клетки и представляет собой синтез углеводов в хлорофиллоносных клетках, идущий с потреблением энергии солнечного света. Различают световую и темповую фазы фотосинтеза. Световая фаза при непосредственном потреблении квантов света обеспечивает процесс синтеза необходимой энергией в виде НАДН и АТФ. Темновая фаза — без участия света, но путем многочисленного ряда химических реакций (цикл Кальвина) обеспечивает образование углеводов, главным образом глюкозы. Значение фотосинтеза в биосфере огромно.
Источник: doklad-referat.ru
Определение фотосинтеза
Фотосинтез — это химический процесс, посредством которого растения, некоторые бактерии и водоросли производят глюкозу и кислород из углекислого газа и воды, используя только свет в качестве источника энергии.
Этот процесс чрезвычайно важен для жизни на Земле, поскольку благодаря ему выделяется кислород, от которого зависит вся жизнь.
Зачем растениям нужна глюкоза (пища)?
Подобно людям и другим живым существам, растения также нуждаются в питании для поддержания жизнедеятельности. Значение глюкозы для растений заключается в следующем:
- Глюкоза, полученная в результате фотосинтеза, используется во время дыхания для высвобождения энергии, необходимой растению для других жизненно важных процессов.
- Растительные клетки также превращают часть глюкозы в крахмал, который используют по мере необходимости. По этой причине мертвые растения используются в качестве биомассы, ведь в них хранится химическая энергия.
- Глюкоза также необходима, чтобы производить другие химические вещества, такие как белки, жиры и растительные сахара, необходимые для обеспечения роста и других важных процессов.
Фазы фотосинтеза
Процесс фотосинтеза разделен на две фазы: световую и темновую.
 Световая фаза фотосинтеза
Световая фаза фотосинтеза
Как следует из названия, световые фазы нуждаются в солнечном свете. В светозависимых реакциях энергия солнечного света поглощается хлорофиллом и преобразуется в запасенную химическую энергию в виде молекулы электронного носителя НАДФН (никотинамидадениндинуклеотидфосфат) и молекулы энергии АТФ (аденозинтрифосфат). Световые фазы протекают в тилакоидных мембранах в пределах хлоропласта.
Темновая фаза фотосинтеза или цикл Кальвина
В темновой фазе или цикле Кальвина возбужденные электроны из световой фазы обеспечивают энергию для образования углеводов из молекул углекислого газа. Не зависящие от света фазы иногда называют циклом Кальвина из-за цикличности процесса.
Хотя темновые фазы не используют свет в качестве реагента (и, как результат, могут происходить днем или ночью), им необходимо, чтобы продукты светозависимых реакций функционировали. Независимые от света молекулы зависят от молекул энергоносителей — АТФ и НАДФН — для создания новых молекул углеводов. После передачи энергии молекулы энергоносители возвращаются к световым фазам для получения более энергичных электронов. Кроме того, несколько ферментов темновой фазы активируются с помощью света.
Схема фаз фотосинтеза
Заметка: Это означает, что темновые фазы не будут продолжаться, если растения будут лишены света слишком долго, так как они используют продукты световых фаз.
Строение листьев растений
Мы не можем полностью изучить фотосинтез, не зная больше о строении листа. Лист адаптирован для того, чтобы играть жизненно важную роль в процессе фотосинтеза.
Внешнее строение листьев
- Площадь
Одной из самых главных особенностей растений является большая площадь поверхности листьев. Большинство зеленых растений имеют широкие, плоские и открытые листья, которые способны захватывать столько солнечной энергии (солнечного света), сколько необходимо для фотосинтеза.
- Центральная жилка и черешок
Центральная жилка и черешок соединяются вместе и являются основанием листа. Черешок располагает лист таким образом, чтобы он получал как можно больше света.
- Листовая пластинка
Простые листья имеют одну листовую пластину, а сложные — несколько. Листовая пластинка — одна из самых главных составляющих листа, которая непосредственно участвует в процессе фотосинтеза.
- Жилы
Сеть жилок в листьях переносит воду от стеблей к листьям. Выделяемая глюкоза также направляется в другие части растения из листьев через жилки. Кроме того, эти части листа поддерживают и удерживают листовую пластину плоской для большего захвата солнечного света. Расположение жилок (жилкование) зависит от вида растения.
- Основание листа
Основанием листа выступает самая нижняя его часть, которая сочленена со стеблем. Зачастую, у основания листа располагается парное количество прилистников.
- Край листа
В зависимости от вида растения, край листа может иметь различную форму, включая: цельнокрайнюю, зубчатую, пильчатую, выемчатую, городчатую и т.п.
- Верхушка листа
Как и край листа, верхушка бывает различной формы, включая: острую, округлую, туповатую, вытянутую, оттянутою и т.д.
Внутреннее строение листьев
Ниже представлена близкая схема внутреннего строения тканей листьев:
- Кутикула
Кутикула выступает главным, защитным слоем на поверхности растения. Как правило, она толще на верхней части листа. Кутикула покрыта веществом, похожим на воск, благодаря которому защищает растение от воды.
- Эпидермис
Эпидермис — слой клеток, который является покровной тканью листа. Его главная функция — защита внутренних тканей листа от обезвоживания, механических повреждений и инфекций. Он также регулирует процесс газообмена и транспирации.
- Мезофилл
Мезофилл — это основная ткань растения. Здесь происходит процесс фотосинтеза. У большинства растений мезофилл разделен на два слоя: верхний — палисадный и нижний — губчатый.
- Защитные клетки
Защитные клетки — специализированные клетки в эпидермисе листьев, которые используются для контроля газообмена. Они выполняют защитную функцию для устьица. Устьичные поры становятся большими, когда вода есть в свободном доступе, в противном случае, защитные клетки становятся вялыми.
- Устьице
Фотосинтез зависит от проникновения углекислого газа (CO2) из воздуха через устьица в ткани мезофилла. Кислород (O2), полученный как побочный продукт фотосинтеза, выходит из растения через устьица. Когда устьица открытые, вода теряется в результате испарения и должна быть восполнена через поток транспирации, водой, поглощенной корнями. Растения вынуждены уравновешивать количество поглощенного СО2 из воздуха и потерю воды через устьичные поры.
Условия, необходимые для фотосинтеза
Ниже приведены условия, которые необходимы растениям для осуществления процесса фотосинтеза:
- Углекислый газ. Бесцветный природный газ без запаха, обнаруженный в воздухе и имеет научное обозначение CO2. Он образуется при горении углерода и органических соединений, а также возникает в процессе дыхания.
- Вода. Прозрачное жидкое химическое вещество без запаха и вкуса (в нормальных условиях).
- Свет. Хотя искусственный свет также подходит для растений, естественный солнечный свет, как правило, создает лучшие условия для фотосинтеза, потому что в нем присутствует природное ультрафиолетовое излучение, которое оказывает положительное влияние на растения.
- Хлорофилл. Это зеленый пигмент, найденный в листьях растений.
- Питательные вещества и минералы. Химические вещества и органические соединения, которые корни растений поглощают из почвы.
Что образуется в результате фотосинтеза?
- Глюкоза;
- Кислород.
Заметка: Растения получают CO2 из воздуха через их листья, и воду из почвы через корни. Световая энергия исходит от Солнца. Полученный кислород выделяется в воздух из листьев. Получаемую глюкозу можно превратить в другие вещества, такие как крахмал, который используется как запас энергии.
Если факторы, способствующие фотосинтезу, отсутствуют или присутствуют в недостаточном количестве, это может негативно повлиять на растение. Например, меньшее количество света создает благоприятные условия для насекомых, которые едят листья растения, а недостаток воды замедляет.
Где происходит фотосинтез?
Фотосинтез происходит внутри растительных клеток, в мелких пластидах, называемых хлоропластами. Хлоропласты (в основном встречающиеся в слое мезофилла) содержат зеленое вещество, называемое хлорофиллом. Ниже приведены другие части клетки, которые работают с хлоропластом, чтобы осуществить фотосинтез.
Строение растительной клетки
Функции частей растительной клетки
- Клеточная стенка: обеспечивает структурную и механическую поддержку, защищает клетки от патогенов, фиксирует и определяет форму клетки, контролирует скорость и направление роста, а также придает форму растениям.
- Цитоплазма: обеспечивает платформу для большинства химических процессов, контролируемых ферментами.
- Мембрана: действует как барьер, контролируя движение веществ в клетку и из нее.
- Хлоропласты: как было описано выше, они содержат хлорофилл, зеленое вещество, которое поглощает световую энергию в процессе фотосинтеза.
- Вакуоль: полость внутри клеточной цитоплазмы, которая накапливает воду.
- Клеточное ядро: содержит генетическую марку (ДНК), которая контролирует деятельность клетки.
Хлорофилл поглощает световую энергию, необходимую для фотосинтеза. Важно отметить, что поглощаются не все цветовые длины волны света. Растения в основном поглощают красную и синюю волны — они не поглощают свет в зеленом диапазоне.
Углекислый газ в процессе фотосинтеза
Растения получают углекислый газ из воздуха через их листья. Углекислый газ просачивается через маленькое отверстие в нижней части листа — устьицу.
Нижняя часть листа имеет свободно расположенные клетки, чтобы углекислый газ достиг других клеток в листьях. Это также позволяет кислороду, образующемуся при фотосинтезе, легко покидать лист.
Углекислый газ присутствует в воздухе, которым мы дышим, в очень низких концентрациях и служит необходимым фактором темновой фазы фотосинтеза.
Свет в процессе фотосинтеза
Лист обычно имеет большую площадь поверхности, поэтому он может поглощать много света. Его верхняя поверхность защищена от потери воды, болезней и воздействия погоды восковым слоем (кутикулой). Верх листа находится там, где падает свет. Этот слой мезофилла называется палисадным. Он приспособлен для поглощения большого количества света, ведь в нем находится много хлоропластов.
В световых фазах, процесс фотосинтеза увеличивается с большим количеством света. Больше молекул хлорофилла ионизируется, и больше генерируется АТФ и НАДФН, если световые фотоны сосредоточены на зеленом листе. Хотя свет чрезвычайно важен в световых фазах, необходимо отметить, что чрезмерное его количество может повредить хлорофилл, и уменьшить процесс фотосинтеза.
Световые фазы не слишком сильно зависят от температуры, воды или углекислого газа, хотя все они нужны для завершения процесса фотосинтеза.
Вода в процессе фотосинтеза
Растения получают воду, необходимую для фотосинтеза через свои корни. Они имеют корневые волоски, которые разрастаются в почве. Корни характеризуются большой площадью поверхности и тонкими стенками, что позволяет воде легко проходить сквозь них.
На изображении представлены растения и их клетки с достаточным количеством воды (слева) и ее нехваткой (справа).
Заметка: Корневые клетки не содержат хлоропластов, поскольку они, как правило, находятся в темноте и не могут фотосинтезировать.
Если растение не впитывает достаточное количество воды, оно увядает. Без воды, растение будет не способно фотосинтезировать достаточно быстро, и может даже погибнуть.
Какое значение имеет вода для растений?
- Обеспечивает растворенными минералами, которые поддерживают здоровье растений;
- Является средой для транспортировки минеральных ресурсов;
- Поддерживает устойчивость и прямостояние;
- Охлаждает и насыщает влагой;
- Дает возможность проводить различные химические реакции в растительных клетках.
Значение фотосинтеза в природе
Биохимический процесс фотосинтеза использует энергию солнечного света для преобразования воды и углекислого газа в кислород и глюкозу. Глюкоза используется в качестве строительных блоков в растениях для роста тканей. Таким образом, фотосинтез — это способ, благодаря которому формируются корни, стебли, листья, цветы и плоды. Без процесса фотосинтеза растения не смогут расти или размножаться.
- Продуценты
Из-за фотосинтетической способности, растения известны как продуценты и служат основой почти каждой пищевой цепи на Земле. (Водоросли являются эквивалентом растений в водных экосистемах). Вся пища, которую мы едим, происходит от организмов, являющихся фотосинтетиками. Мы питаемся этими растениями напрямую или едим животных, таких как коровы или свиньи, которые потребляют растительную пищу.
- Основа пищевой цепи
Внутри водных систем, растения и водоросли также составляют основу пищевой цепи. Водоросли служат пищей для беспозвоночных, которые, в свою очередь, выступают источником питания для более крупных организмов. Без фотосинтеза в водной среде жизнь была бы невозможна.
- Удаление углекислого газа
Фотосинтез превращает углекислый газ в кислород. Во время фотосинтеза углекислый газ из атмосферы поступает в растение, а затем выделяется в виде кислорода. В сегодняшнем мире, где уровни двуокиси углерода растут ужасающими темпами, любой процесс, который устраняет углекислый газ из атмосферы, является экологически важным.
- Круговорот питательных веществ
Растения и другие фотосинтезирующие организмы играют жизненно важную роль в круговороте питательных веществ. Азот в воздухе фиксируется в растительных тканях и становится доступным для создания белков. Микроэлементы, находящиеся в почве, также могут быть включены в растительную ткань и стать доступными для травоядных животных, дальше по пищевой цепи.
- Фотосинтетическая зависимость
Фотосинтез зависит от интенсивности и качества света. На экваторе, где солнечный свет обилен весь год и вода не является ограничивающим фактором, растения имеют высокие темпы роста, и могут стать довольно большими. И наоборот, фотосинтез в более глубоких частях океана встречается реже, поскольку свет не проникает в эти слои, и в результате эта экосистема оказывается более бесплодной.
Источник: natworld.info
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
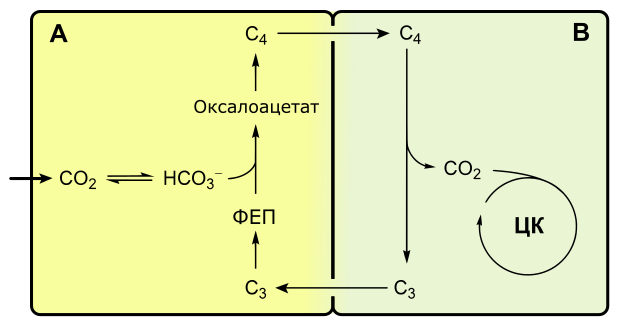
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su