Все про хромосомы
Хромосомы — это основные структурные элементы клеточного ядра, являющиеся носителями генов, в которых закодирована наследственная информация. Обладая способностью к самовоспроизведению, хромосомы обеспечивают генетическую связь поколений.
Морфология хромосом связана со степенью их спирализации. Например, если в стадии интерфазы (см. Митоз, Мейоз) хромосомы максимально развернуты, т. е. деспирализованы, то с началом деления хромосомы интенсивно спирализуются и укорачиваются. Максимальной спирализации и укорочения хромосомы достигают в стадии метафазы, когда происходит формирование относительно коротких, плотных, интенсивно окрашивающихся основными красителями структур. Эта стадия наиболее удобна для изучения морфологических характеристик хромосом.
Метафазная хромосома состоит из двух продольных субъединиц — хроматид [электронная микроскопия выявляет в строении хромосом элементарные нити (так называемые хромонемы, или хромофибриллы) толщиной 200 Å, каждая из которых состоит из двух субъединиц].
Размеры хромосом растений и животных значительно колеблются: от долей микрона до десятков микрон. Средние длины метафазных хромосом человека лежат в пределах 1,5—10 микрон.
Химической основой строения хромосом являются нуклеопротеиды — комплексы нуклеиновых кислот (см.) с основными белками — гистонами и протаминами.
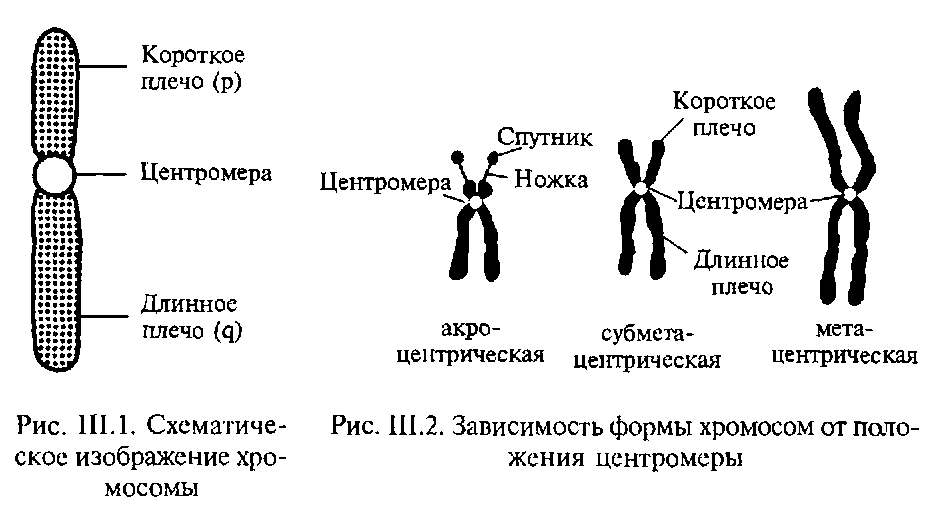
Индивидуальные хромосомы (рис. 1) различают по локализации первичной перетяжки, т. е. места расположения центромеры (во время митоза и мейоза к этому месту прикрепляются нити веретена, подтягивая ее при этом к полюсу). При утрате центромеры фрагменты хромосом утрачивают способность расходиться при делении. Первичная перетяжка делит хромосомы на 2 плеча. В зависимости от расположения первичной перетяжки хромосомы подразделяют на метацентрические (оба плеча равной или почти равной длины), субметацентрические (плечи неравной длины) и акроцентрические (центромера смещена на конец хромосомы). Помимо первичной, в хромосомах могут встречаться менее выраженные вторичные перетяжки. Небольшой концевой участок хромосом, отделенный вторичной перетяжкой, называют спутником.
Каждый вид организмов характеризуется своим специфическим (по числу, размерам и форме хромосом) так называемым хромосомным набором. Совокупность двойного, или диплоидного, набора хромосом обозначают как кариотип.
В зрелых половых клетках, яйцеклетках и сперматозоидах содержится одиночный, или гаплоидный, набор хромосом (n), составляющий половину диплоидного набора (2n), присущего хромосомам всех остальных клеток организма.
диплоидном наборе каждая хромосома представлена парой гомологов, один из которых материнского, а другой отцовского происхождения. В большинстве случаев хромосомы каждой пары идентичны по размерам, форме и генному составу. Исключение составляют половые хромосомы, наличие которых определяет развитие организма в мужском или женском направлении. Нормальный хромосомный набор человека состоит из 22 пар аутосом и одной пары половых хромосом. У человека и других млекопитающих женский пол определяется наличием двух Х-хромосом, а мужской — одной X-и одной Y-хромосомы (рис. 2 и 3). В женских клетках одна из Х-хромосом генетически неактивна и обнаруживается в интерфазном ядре в виде полового хроматина (см.). Изучение хромосом человека в норме и патологии составляет предмет медицинской цитогенетики. Установлено, что отклонения в числе или структуре хромосом от нормы, возникающие в половых! клетках или на ранних этапах дробления оплодотворенной яйцеклетки, вызывают нарушения нормального развития организма, обусловливая в некоторых случаях возникновение части спонтанных абортов, мертворождений, врожденных уродств и аномалий развития после рождения (хромосомные болезни). Примерами хромосомных болезней могут служить болезнь Дауна (лишняя G-хромосома), синдром Клайнфелтера (лишняя Х-хромосома у мужчин) и Шерешевского — Тернера (отсутствие в кариотипе Y- или одной из Х-хромосом). В медицинской практике хромосомный анализ проводят или прямым методом (на клетках костного мозга), или после кратковременного культивирования клеток вне организма (периферическая кровь, кожа, эмбриональные ткани).
Хромосомы (от греч. chroma — окраска и soma — тело) — нитевидные, самовоспроизводящиеся структурные элементы клеточного ядра, содержащие в линейном порядке факторы наследственности — гены. Хромосомы отчетливо видны в ядре во время деления соматических клеток (митоза) и во время деления (созревания) половых клеток — мейоза (рис. 1). В том и в другом случае хромосомы интенсивно окрашиваются основными красителями, а также видны на неокрашенных цитологических препаратах в фазовом контрасте. В интерфазном ядре хромосомы деспирализованы и не видны в световой микроскоп, так как их поперечные размеры выходят за пределы разрешающей способности светового микроскопа. В это время отдельные участки хромосом в виде тонких нитей диаметром 100—500 Å можно различить при помощи электронного микроскопа. Отдельные не деспирализовавшиеся участки хромосом в интерфазном ядре видны через световой микроскоп как интенсивно красящиеся (гетеропикнотические) участки (хромоцентры).
Хромосомы непрерывно существуют в клеточном ядре, претерпевая цикл обратимой спирализации: митоз—интерфаза—митоз. Основные закономерности строения и поведения хромосом в митозе, мейозе и при оплодотворении одинаковы у всех организмов.
Хромосомная теория наследственности. Впервые хромосомы описали И. Д. Чистяков в 1874 г.
Страсбургер (Е. Strasburger) в 1879 г. В 1901 г. Уилсон (Е. В. Wilson), а в 1902 г. Саттон (W. S. Sutton) обратили внимание на параллелизм в поведении хромосом и менделевских факторов наследственности — генов — в мейозе и при оплодотворении и пришли к выводу, что гены находятся в хромосомах. В 1915—1920 гг. Морган (Т. Н. Morgan) и его сотрудники доказали это положение, локализовали в хромосомах дрозофилы несколько сот генов и создали генетические карты хромосом. Данные о хромосомах, полученные в первой четверти 20 века, легли в основу хромосомной теории наследственности, согласно которой преемственность признаков клеток и организмов в ряду их поколений обеспечивается преемственностью их хромосомах.
Химический состав и ауторепродукция хромосом. В результате цитохимических и биохимических исследований хромосом в 30 и 50-х годах 20 века установлено, что они состоят из постоянных компонентов [ДНК (см. Нуклеиновые кислоты), основных белков (гистонов или протаминов), негистонных белков] и переменных компонентов (РНК и связанного с ней кислого белка). Основу хромосом составляют дезоксирибонуклеопротеидные нити диаметром около 200 Å (рис. 2), которые могут соединяться в пучки диаметром 500 А.
Открытие Уотсоном и Криком (J. D. Watson, F. Н. Crick) в 1953 г. строения молекулы ДНК, механизма ее авторепродукции (редупликации) и нуклеинового кода ДНК и развитие возникшей после этого молекулярной генетики привело к представлению о генах как участках молекулы ДНК. (см. Генетика). Вскрыты закономерности авторепродукции хромосом [Тейлор (J. Н. Taylor) и др., 1957], оказавшиеся аналогичными закономерностям авторепродукции молекул ДНК (полуконсервативная редупликация).
Хромосомный набор — совокупность всех хромосом в клетке. Каждый биологический вид обладает характерным и постоянным набором хромосом, закрепленным в эволюции данного вида. Различают два основных типа наборов хромосом: одиночный, или гаплоидный (в половых клетках животных), обозначаемый n, и двойной, или диплоидный (в соматических клетках, содержащий пары сходных, гомологичных хромосом от матери и отца), обозначаемый 2n.
Наборы хромосом отдельных биологических видов значительно различаются по числу хромосом: от 2 (лошадиная аскарида) до сотен и тысяч (некоторые споровые растения и простейшие). Диплоидные числа хромосом некоторых организмов таковы: человека — 46, гориллы — 48, кошки — 60, крысы — 42, дрозофилы — 8.
Размеры хромосом у разных видов также различны. Длина хромосом (в метафазе митоза) варьирует от 0,2 мк у одних видов до 50 мк у других, а диаметр от 0,2 до 3 мк.
Морфология хромосом хорошо выражена в метафазе митоза. Именно метафазные хромосомы используют для идентификации хромосом. В таких хромосомах хорошо видны обе хроматиды, на которые продольно расщеплена каждая хромосома и центромер (кинетохор, первичная перетяжка), соединяющий хроматиды (рис. 3). Центромер виден как суженный участок, не содержащий хроматина (см.); к нему крепятся нити ахроматинового веретена, благодаря чему центромер определяет движение хромосом к полюсам в митозе и мейозе (рис. 4).
Потеря центромера, например при разрыве хромосомы ионизирующими излучениями или другими мутагенами, приводит к потере способности куска хромосомы, лишенного центромера (ацентрический фрагмент), участвовать в митозе и мейозе и к потере его из ядра. Это может привести к тяжелому повреждению клетки.
Центромер делит тело хромосомы на два плеча. Расположение центромера строго постоянно для каждой хромосомы и определяет три типа хромосом: 1) акроцентрические, или палочкообразные, хромосомы с одним длинным и вторым очень коротким плечом, напоминающим головку; 2) субметацентрические хромосомы с длинными плечами неравной длины; 3) метацентрические хромосомы с плечами одинаковой или почти одинаковой длины (рис. 3, 4, 5 и 7).
Характерными чертами морфологии определенных хромосом являются вторичные перетяжки (не обладающие функцией центромера), а также спутники — маленькие участки хромосом, соединенные с остальным ее телом тонкой нитью (рис. 5). Спутничные нити обладают способностью формировать ядрышки. Характерная структура в хромосоме (хромомеры) — утолщения или более плотно спирализованные участки хромосомной нити (хромонемы). Рисунок хромомер специфичен для каждой пары хромосом.
Число хромосом, их размеры и форма на стадии метафазы характерны для каждого вида организмов. Совокупность этих признаков набора хромосом называется кариотипом. Кариотип можно представить в виде схемы, называемой идиограммой (см. ниже хромосомы человека).
Половые хромосомы. Гены, детерминирующие пол, локализованы в специальной паре хромосом — половых хромосомах (млекопитающие, человек); в других случаях иол определяется соотношением числа половых хромосом и всех остальных, называемых аутосомами (дрозофила). У человека, как и у других млекопитающих, женский пол определяется двумя одинаковыми хромосомами, обозначаемыми как Х-хромосомы, мужской пол определяется парой гетероморфных хромосом: Х и Y. В результате редукционного деления (мейоза) при созревании ооцитов (см. Овогенез) у женщин все яйца содержат по одной Х-хромосоме. У мужчин в результате редукционного деления (созревания) сперматоцитов половина спермиев содержит Х-хромосому, а другая половина Y-хромосому. Пол ребенка определяется случайным оплодотворением яйцеклетки спермием, несущим Х- или Y-хромосому. В результате возникает зародыш женского (XX) или мужского (XY) пола. В интерфазном ядре у женщин одна из Х-хромосом видна как глыбка компактного полового хроматина.
Функционирование хромосом и метаболизм ядра. Хромосомная ДНК является матрицей для синтеза специфических молекул информационной РНК. Этот синтез происходит тогда, когда данный участок хромосомы деспирализован. Примерами локальной активации хромосом служат: образование деспирализованных петель хромосом в ооцитах птиц, амфибий, рыб (так называемые Х-ламповые щетки) и вздутий (пуффов) определенных локусов хромосом в многонитчатых (политенных) хромосомах слюнных желез и других секреторных органов двукрылых насекомых (рис. 6). Примером инактивации целой хромосомы, т. е. выключения ее из метаболизма данной клетки, является образование одной из Х-хромосом компактного тела полового хроматина.
Рис. 6. Политенные хромосомы двукрылого насекомого Acriscotopus lucidus: А и Б — участок, ограниченный пунктирными линиями, в состоянии интенсивного функционирования (пуфф); В — тот же участок в нефункционирующем состоянии. Цифрами обозначены отдельные локусы хромосом (хромомеры).
Рис. 7. Хромосомный набор в культуре лейкоцитов периферической крови мужчины (2n=46).
Вскрытие механизмов функционирования политенных хромосом типа ламповых щеток и других типов спирализации и деспирализации хромосом имеет решающее значение для понимания обратимой дифференциальной активации генов.
Хромосомы человека. В 1922 г. Пейнтер (Т. S. Painter) установил диплоидное число хромосом человека (в сперматогониях), равное 48. В 1956 г. Тио и Леван (Н. J. Tjio, A. Levan) использовали комплекс новых методов исследования хромосом человека: культуру клеток; исследование хромосом без гистологических срезов на тотальных препаратах клеток; колхицин, приводящий к остановке митозов на стадии метафазы и накоплению таких метафаз; фитогемагглютинин, стимулирующий вступление клеток в митоз; обработку метафазных клеток гипотоническим солевым раствором.
е это позволило уточнить диплоидное число хромосом у человека (оно оказалось равным 46) и дать описание кариотипа человека. В 1960 г. в Денвере (США) международная комиссия разработала номенклатуру хромосом человека. Согласно предложениям комиссии, термин «кариотип» следует применять к систематизированному набору хромосом единичной клетки (рис. 7 и 8). Термин «идиотрамма» сохраняется для представления о наборе хромосом в виде диаграммы, построенной на основании измерений и описания морфологии хромосом нескольких клеток.
Хромосомы человека пронумерованы (отчасти серийно) от 1 до 22 в соответствии с особенностями морфологии, допускающими их идентификацию. Половые хромосомы не имеют номеров и обозначаются как Х и Y (рис. 8).
Обнаружена связь ряда заболеваний и врожденных дефектов в развитии человека с изменениями в числе и структуре его хромосом. (см. Наследственность).
См. также Цитогенетические исследования.
Все эти достижения создали прочную базу для развития цитогенетики человека.
Источник: www.medical-enc.ru
Что такое хромосомы
Хромосомы представляют собой нуклеопротеидные структуры эукариотической клетки, в которых хранится большая часть наследственной информации. Благодаря своей способности к самовоспроизведению, именно хромосомы обеспечивают генетическую связь поколений. Хромосомы образуются из длинной молекулы ДНК, в которой содержится линейная группа множества генов, и вся генетическая информация будь-то о человеке, животном, растении или любом другом живом существе.
Морфология хромосом связана с уровнем их спирализации. Так, если во время стадии интерфазы хромосомы максимально развернуты, то с началом деления хромосомы активно спирализуются и укорачиваются. Своего максимального укорочения и спирализации они достигают во время стадии метафазы, когда происходит формирование новых структур. Эта фаза наиболее удобна для изучения свойств хромосом, их морфологических характеристик.
История открытия хромосом
Еще в середине позапрошлого XIX века многие биологи изучая в микроскопе строение клеток растений и животных, обратили внимание на тонкие нити и мельчайшие кольцевидные структуры в ядре некоторых клеток. И вот немецкий ученый Вальтер Флеминг применив анилиновые красители для обработки ядерных структур клетки, что называется «официально» открывает хромосомы. Точнее обнаруженное вещество было им названо «хроматид» за его способность к окрашиванию, а термин «хромосомы» в обиход чуть позже (в 1888 году) ввел еще один немецкий ученый – Генрих Вайлдер. Слово «хромосома» происходит от греческих слов «chroma» — окраска и «somo» — тело.

Хромосомная теория наследственности
Разумеется, история изучения хромосом не закончилась на их открытии, так в 1901-1902 годах американские ученые Уилсон и Сатон, причем независимо друг от друга, обратили внимание на сходство в поведении хромосом и менделеевских факторов наследственности — генов. В результате ученые пришли к заключению, что гены находятся в хромосомах и именно посредством их из поколения в поколения, от родителей к детям передается генетическая информация.
В 1915-1920 годам участие хромосом в передаче генов было доказано на практике в целой серии опытов, сделанных американским ученым Морганом и сотрудниками его лаборатории. Им удалось локализировать в хромосомах мухи-дрозофилы несколько сот наследственных генов и создать генетические карты хромосом. На основе этих данных была создана хромосомная теория наследственности.
Строение хромосом
Строение хромосом разнится в зависимости от вида, так метафазная хромосома (образующаяся в стадии метафазе при митозном делении клетки) состоит из двух продольных нитей – хроматид, которые соединяются в точке, именуемой центромерой. Центромера – это участок хромосомы, который отвечает за расхождение сестринских хроматид в дочерние клетки. Она же делит хромосому на две части, названные коротким и долгим плечом, она же отвечает за деление хромосомы, так как именно в ней содержится специальное вещество – кинетохор, к которому крепятся структуры веретена деления.
Тут на картинке показано наглядное строение хромосомы: 1. хроматиды, 2. центромера, 3. короткое плечо хроматид, 4. длинное плечо хроматид. На концах хроматид располагаются теломеры, специальные элементы, которые защищают хромосому от повреждений и препятствуют слипанию фрагментов.
Формы и виды хромосом
Размеры хромосом растений и животных значительно различаются: от долей микрона до десятков микрон. Средние длины метафазных хромосом человека лежат в диапазоне от 1,5 до 10 микрон. В зависимости от вида хромосомы отличаются и ее способности к окрашиванию. В зависимости от расположения центромеры различают такие формы хромосом:
- Метацентрические хромосомы, для которых характерно срединное расположение центромеры.
- Субметацентрические, для них характерно неравномерное расположение хроматид, когда одно плечо более длинное, а второе более короткое.
- Акроцентрические или палочковидные. У них центромера расположена практически в самом конце хромосомы.
Функции хромосом
Основные функции хромосом, как для животных, так и для растений и вообще всех живых существ – передача наследственной, генетической информации от родителей к детям.
Набор хромосом
Значение хромосом столь велико, что их количество в клетках, а также особенности каждой хромосомы определяют характерный признак того или иного биологического вида. Так, например, у мухи-дрозофилы в наличии 8 хромосом, у обезьян – 48, а хромосомный набор человека составляет 46 хромосом.
В природе существует два основных типа набора хромосом: одиночный или гаплоидный (содержится в половых клетках) и двойной или диплоидный. Диплоидный набор хромосом имеет парную структуру, то есть вся совокупность хромосом состоит из хромосомных пар.
Хромосомный набор человека
Как мы уже написали выше, клетки человеческого организма содержат 46 хромосом, которые объединены в 23 пары. Все вместе они и составляют хромосомный набор человека. Первые 22 пары человеческих хромосом (их называют аутосомами) являются общими как для мужчин, так и для женщин, и лишь 23 пара — половых хромосом — разнится у разных полов, она же определяет половую принадлежность человека. Совокупность всех пар хромосом также называется кариотипом.
Такой вид имеет хромосомный набор человека, 22 пары двойных диплоидных хромосом содержат всю нашу наследственную информацию, и последняя пара различается, у мужчин она состоит из пары условных X и Y половых хромосом, в то время как у женщин в наличии две хромосомы Х.
Аналогичную структуру хромосомного набора имеют и все животные, только количество неполовых хромосом у каждого из них свое.
Генетические болезни, связанные с хромосомами
Нарушение в работе хромосом, или даже само их неправильно количество является причиной многих генетических заболеваний. Например, синдрома Дауна появляется из-за наличия лишней хромосомы в хромосомном наборе человека. А такие генетические болезни как дальтонизм, гемофилия вызваны сбоями в работе имеющихся хромосом.
Источник: www.poznavayka.org
Весь механизм управления развитием и активностью организма заключен в дезоксирибонуклеиновой кислоте (ДНК), из которой состоят хромосомы клеточных ядер и их основные функциональные единицы — гены. ДНК состоит из двух длинных параллельных макромолекулярных цепочек, свернутых в виде спирали; молекулы ДНК состоят из трех основных элементов: фосфатных молекул, молекул сахара — дезоксирибозы и четырех азотистых оснований: аденина, гуанина, тимина и цитозина. Каждая цепочка ДНК состоит из последовательных звеньев, которые называются нуклеотидами: волокна азотистых оснований соединены водородными связями друг с другом таким образом, что двойная спираль ДНК по форме похожа на винтовую лестницу.
Деление всех клеток организма, за исключением клеток зародыша, происходит путем удваивания хромосомного материала, поскольку каждая из дочерних клеток должна получить точную копию ДНК от материнской клетки. В ходе этого процесса, который называется репликацией, две цепи ДНК разъединяются и благодаря воздействию особого фермента формируются две новые комплементарные цепи. Каждая исходная цепь служит прообразом для новой цепи, в которой азотистые основания соединяются (соединяться между собой могут только аденин и тимин или цитозин и гуанин), — в результате каждая из двух цепей получается дополненной. Таким образом, воссоздаются две идентичные макромолекулы ДНК, поскольку каждая цепь состоит из исходной и новой цепи.
Хотя многие хромосомы отличаются по размеру, все они имеют одинаковую форму маленькой палочки, которую сжимает центромера, разделяющая хромосому на два плеча, часто неодинаковой длины. Однако типичное изображение хромосом соответствует стадии процесса клеточного деления, когда ДНК уже дублировалась, — момент, в который хромосома похожа на букву X с двумя длинными и двумя короткими плечами.
Количество хромосом человека со всеми генами определяющими развитие и функционирование нашего организма, соответствует количеству хромосом материнской яйцеклетки и отцовского сперматозойда, которые соединяются в момент оплодотворения.
Это возможно из-за того, что, в отличие от остальных клеток человеческого организма, в которых содержится по 46 хромосом, гаметы содержат всего по 23 хромосомы: слияние яйцеклетки и сперматозоида образует зиготу с 46 хромосомами (23 пары родительских хромосом), клетки зиготы постоянно делятся, и в результате их деления образуется эмбрион человека с набором хромосом, идентичным родительским. Таким образом, каждый человек получает половину материнского и половину отцовского хромосомного набора.
Источник: tardokanatomy.ru
Все, что изложено выше относительно химического состава и структуры хромосом эукариот, типично и для хромосом человека. Некоторой детализации требует информация, позволяющая идентифицировать с большей степенью точности любую хромосому человека.
1956 год — шведы Тио и Леван, англичане Форд и Хамертон установили, что ядро диплоидной клетки человека содержит 46 хромосом – это хромосомный набор или кариотип человека; в 1960 – Мурхед и сотр. (США) разработали метод приготовления препаратов хромосом из кратковременной культуры лимфоцитов; в 1968-70 гг. разработаны методы дифференциального окрашивания хромосом, что позволило однозначно идентифицировать все хромосомы человека — все эти манипуляции производились и производятся только на метафазных хромосомах, ибо они различимы лучше всего, т.к. они максимально укорочены и утолщены, лежат свободно одна от другой, располагаются все в одной плоскости клетки (экваториальной); кроме того, исследуются только те метафазные хромосомы, хроматиды которых отделились друг от друга в области плечей, а в центромерной части еще соединены.
Совокупность всех метафазных хромосом, расположенных относительно произвольно в экваториальной плоскости клетки, именуется метафазной пластинкой или просто хромосомным набором. После приготовления препаратов хромосом, которые можно приготовить из всех тканей и клеточных суспензий, содержащих делящиеся клетки (в зависимости от целей важно, конечно, количество метафаз), хромосомы окрашиваются, ибо только после этого их можно различить в световой микроскоп, получить микрофотографию, идентифицировать и, расположив их в определенном порядке, т.е. составив кариограмму, получить целостное представление о кариотипе конкретного человека. Кариограмма — это те же хромосомы метафазной пластинки, но расположенные упорядоченно. Принцип упорядоченности общий для всего вида и определяется идеограммой. Идиограмма — это графическое изображение гаплоидного набора хромосом (можно и диплоидного) и расположение их по группам в зависимости от формы и величины. Группы располагаются в порядке уменьшения величины входящих в них хромосом.
В современных цитогенетических лабораториях процесс составления кариограммы компьютеризирован.
Наиболее простой способ окрашивания хромосом красителем Гимза или 2%-ым ацетоорсеином, или 2%-ым ацетокармином. При этом хромосомы окрашиваются целиком, равномерно и интенсивно. Окрашенные таким образом хромосомы, согласно Денверской классификации (I960), располагались в идиограмме в зависимости от их длины и нумеровались по парам от 1 до 23. Тогда же Патау предложил разбить 23 пары хромосом на 7 групп от А до G с учетом расположения центромеры. Важным признаком, уточняющим форму хромосомы, стал центромерный индекс: отношение длины короткого плеча к длине всей хромосомы, выраженное в %. Комплекс этих параметров позволял с немалой степенью точности распределить хромосомы по группам, но идентифицировать их, особенно в группах В, С, D, F и G, было невозможно.
Однако уже при стандартном (рутинном) равномерном окрашивании хромосом замечали, но оставили без внимания, некоторую неоднородность в плотности окрашивания по длине хромосом. И только позже (1968 г.), когда Касперсон с сотрудниками обнаружили, что после обработки акрихин-ипритом флуоресценция по длине хромосом распределена не равномерно, а в виде сегментов, они показали, что каждую хромосому можно надежно идентифицировать с помощью такого метода дифференциального окрашивания, ибо расположение сегментов для каждой хромосомы строго специфично. Вскоре стало ясно, что очень сходный рисунок сегментации хромосом можно получить и с помощью красителя Гимза, дополнив окрашивание некоторыми приемами. Впоследствии при разных способах обработки хромосом были обнаружены разные типы сегментов.
На Парижской конференции по стандартизации и номенклатуре хромосом человека (1971) все полученные к тому времени данные по дифференциальному окрашиванию хромосом были сопоставлены и оказалось, что все методы в принципе выявляют одни и те же структуры, но каждый специфичен в отношении определенных сегментов. И обозначать различные типы сегментов решили по методам, с помощью которых они выявляются.
Q — сегменты — флуоресцирующие после окраски акрихин-ипритом;
G — сегменты (Гимза) — выявляются при окрашивании красителем Гимза в сочетании с дополнительными процедурами; Q и G сегменты идентичны, но в большинстве лабораторий предпочитают этот метод, т.к. он не требует использования флуоресцентного микроскопа и эти препараты дольше хранятся; однако, только с помощью Q-метода можно идентифицировать Y-хромосому человека даже в интерфазном ядре;
R — сегменты — окрашиваются после контролируемой тепловой денатурации, располагаются между Q и G — сегментами;
С — сегменты — конститутивный гетерохроматин, располагается в прицентромерных районах обоих плечей хромосомы;
Т — сегменты — расположены в теломерных (концевых) районах хромосом.
Химическая природа дифференциального окрашивания еще только исследуется. Обсуждаются две основные гипотезы: первая исходит из того, что различные участки хромосом человека отличаются по количественному содержанию пар оснований аденин — тимин и гуанин — цитозин. Отсюда разная степень усвоения ими красителей. В частности, блоки с большим содержанием пар А-Т связываются преимущественно с акрихин-ипритом, следовательно, Q-сегменты соответствуют участкам, богатым А-Т — парами; R-сегменты соответствуют участкам, богатым Г-Ц — парами, которые более устойчивы к тепловой денатурации — это, однако, не объясняет всех особенностей сегментации хромосом. Вторая гипотеза, белковая, исходит из данных о том, что предварительная протеолитическая обработка перед окрашиванием красителем Гимза индуцирует появление G-сегментов, а так как разные по составу участки ДНК связаны с разными белками, можно полагать, что рисунок сегментации зависит от особенностей комплекса ДНК — белок.
И все же, что собой представляют полосы — сегменты митотических хромосом, остается загадкой. Даже небольшие тонкие полосы содержат не менее 30 гигантских петель, суммарный нуклеотидный состав которых более 1 млн., нуклеотидов. Возможно, существование таких структурных блоков связано с функционированием эукариотического генома вообще, хотя сами по себе сегменты ничего конкретного о функционировании индивидуальных генов не говорят, ибо в самой тонкой полосе, которую еще можно различить, содержится от 10 до 100 генов. Но то, что картина распределения сегментов в хромосомах почти не изменилась за долгие периоды эволюции (почти каждая хромосома человека имеет своего аналога в кариотипе шимпанзе, гориллы, орангутана), свидетельствует о большом значении пространственной организации ДНК для экспрессии соответствующих генов.
Итак, информация, полученная в результате анализа дифференциально окрашенных хромосом, позволяет представить идиограмму хромосом человека следующим образом:
Группа А, 1-3 хромосомы — большие метацентрические и субметацентрические хромосомы; 1-ая — самая большая метацентрическая, центромерный индекс (ЦИ) 48 — 49%, в длинном плече вблизи центромеры часто обнаруживается вторичная перетяжка; вторая самая большая субметацентрическая ЦИ 38-40%; 3-я -почти на 20% короче 1 -ой, ЦИ 45-46%, метацентрическая.
Группа В, 4 и 5 хромосомы — большие субметацентрические. ЦИ 24-30%, без дифференциального окрашивания друг от друга не отличаются.
Группа С, 6-12 хромосомы и Х-хромосома — средние Субметацентрические хромосомы 6, 7, 8,11 и 12 — относительно субметацентрические, ЦИ — 27-35; 11 и 12 обнаруживают очень сходный рисунок сегментации, однако 11-я хромосома более метацентрическая; в 9-й в длинном плече часто обнаруживают вторичную перетяжку, которая не окрашивается ни акрихином, ни красителем Гимза; Х-хромосома значительно варьирует по длине, в целом сходна с самыми длинными из С-группы, ЦИ — 40,12+2,12, отличить от других при стандартном окрашивании очень трудно.
Группа D, 13-15 хромосомы — акроцентрические, ЦИ около 15 -наименьший в кариотипе человека, все они могут иметь вторичную перетяжку на коротком плече или не иметь, а следовательно, иметь спутники или не иметь, спутники могут быть очень большими, а иногда двойными; короткие плечи этих хромосом содержат ядрышковый организатор.
Группа Е, 16-18 хромосомы — относительно короткие метацентрические и субметацентрические; 16 — ЦИ — около 40, длина вариабельна, в длинном плече в 10% случаев выявляется вторичная перетяжка; 17-я, ЦИ -31; 18-ая хромосома на 5 -10%короче17,ЦИ-26.
Группа F, 19, 20 хромосомы — мелкие метацентрические, ЦИ — 36-46, при стандартной окраске выглядят одинаково, при дифференциальной — резко отличаются.
Группа G, 21, 22, Y-хромосомы — мелкие акроцентрические, ЦИ — 13-33; 21 и 22-ая могут иметь спутники, короткие плечи имеют ядрышковый организатор; Y-хромосома обычно (но не всегда) больше, хроматиды ее длинного плеча, как правило, лежат параллельно одна другой, а у 21 и 22 — ой хромосомы они чаще образуют широкий угол; спутники в Y-хромосоме отсутствуют, ЦИ от 0 до 26. В интерфазных ядрах дистальный участок длинного плеча при окрашивании акрихин-ипритом сильно флуоресцирует и выявляется как яркое пятно, которое называется Y-хроматин.
В соответствии с Парижской номенклатурой в хромосомах идиограммы показан рисунок сегментации (G — Q , R — сегменты) — позитивные светлые G (они же Q) сегменты, негативные темные — R, районы с варьирующей окраской заштриховываются. Латинскими буквами р и q обозначаются соответственно короткое и длинное плечо, в каждом плече выделяются районы, обозначенные арабскими цифрами, районы нумеруются от центромеры к теломерным участкам хромосомы. А уже внутри района выделяются сегменты (англ. bands), обозначенные арабскими цифрами по такому же принципу, т.е. сегмент имеет свой символ, например, 1 q 32 — второй сегмент третьего района в длинном плече 1 -ой хромосомы (при чтении справа налево).
Если в заключение сформулировать интегральную модель хромосомы, то она состоит из единственной двойной спирали ДНК, объединенной с гистонами в нуклеосомы. Некоторые районы этой двойной спирали представлены повторяющимися последовательностями, которые могут быть рассеяны по всему геному. Участки с повторяющимися последовательностями обнаруживают признаки конститутивного гетерохроматина. Участки с уникальными последовательностями пар нуклеотидов проявляют свойства эухроматина, это транскрибирующиеся участки — т.е. собственно гены, они соответствуют светлым G- и темным R-сегментам дифференциально окрашенных хромосом.
Благодаря успехам в молекулярной генетике человека разработан принципиально новый метод изучения хромосом – метод флюоресцентной гибридизации in situ (FISH) (in situ – в месте нахождения).
Суть этого молекулярно-цитогенетического метода заключается в следующем:
1 – для изучаемой хромосомы или ее конкретного участка готовят комплементарный однонитевой участок ДНК, к которому присоединяют биотин или дигоксигенин, — такой помеченный участок ДНК называется зондом;
2 – на микроскопическом препарате хромосом (in situ) при обработке щелочью хромосомная ДНК денатурирует, т.е. разрываются водородные связи между двумя комплементарными нитями ДНК;
3 – полученным ранее зондом обрабатывают препарат – зонд присоединяется к хромосоме в комплементарном участке ДНК, происходит ренатурация – между зондом и соответствующим участком молекулы ДНК формируются водородные связи;
4 – затем препарат обрабатывают веществом, которое избирательно может присоединяться к биотину или дигоксигенину, после чего к зонду можно присоединить флюоресцентный краситель (или красный – родамин, или зеленый – флюоресцеина изотиоцианат);
5 – теперь с помощью люминесцентного микроскопа можно увидеть окрашенные хромосомы на фоне неокрашенных.
Можно использовать не только двух – но и трехцветные варианты метода.
Метод FISH применяется очень широко – от определения локализации гена, до расшифровки сложных перестроек между несколькими хромосомами. Он требует меньше времени, чем кариотипирование дифференциально окрашенных хромосом.
Метод FISH можно применять для диагностики анеуплоидий в интерфазных ядрах – интерфазная цитогенетика. Например, в течение нескольких часов можно получить информацию о количестве 21 — х хромосом в клетках амниотической жидкости (пренатальная – дородовая диагностика синдрома Дауна у плода) – специфический ДНК – зонд для 21 — ой хромосомы покажет в ядрах этих клеток или 2 — е светящиеся точки, что соответствует двум 21-м хромосомам, или три – что выявит трисомию по 21-ой хромосоме.
Источник: StudFiles.net