Морфология половых клеток
Зрелые половые клетки содержат гаплоидный набор хромосом. Созревающие -диплоидный. Имеют ядро, цитоплазму, клеточные органеллы. Несмотря на это, строение мужских и женских половых клеток неодинаково. Это объясняется различными функциями. Функции сперматозоида – оплодотворение (стимуляция дальнейшего развития яйцеклетки), обеспечение генетической информацией мужского организма. Все сперматозоиды имеют жгутики, подвижны, небольшого размера (50-90мкм у человека). Состоят из головки, шейки, средней части и хвостика. Головка -5мкм, шейка – 5. головка сперматозоида почти полностью занята ядром, цитоплазмы мало, она в жидкокристаллическом состоянии (защита от вредных явлений – ионизирующего излучения). Находится по периферии ядра. На конце головки – акросома с видоизмененным комплексом Гольджи. Ферменты: гиалуронидаза, муциназа. В плазматической мембране – проакрозин, который превращается в акрозин, проходя по половым путям самки (происходит отщепление ингибитора). Функция акрозина – отщепление фолликулярных клеток, отщепление блестящей оболочки. Шейка содержит пару центриолей. Микротрубочки одной из них удлиняются, образуется основная нить хвостика. В шейке много митохондрий, расположенных по спирали. Органеллы движения – жгутики, способны к биению только при смешивании с секретом. Предстательной железы при семяизвержении. При нарушении функций предстательной железы – мужская стерильность.
Яйцеклетка .Функции: передает зародышу половину его будущего хромосомного набора; во время оплодотворения яйцеклетка приносит гораздо больше цитоплазмы; яйцеклетка снабжает зародыш пищевыми запасами до начала его собственного питания.
Размеры яйцеклеток много больше размеров сперматозоидов(130-150 мкм у человека). В зрелой яйцеклетке запасаются все материалы, которые обеспечивают начальные стадии развития зародыша. Если сперматозоид, созревая, старается избавиться от цитоплазмы, яйцеклетка, наоборот, стремится увеличить ее количество. Есть рибосомы, рРНК, т-РНК, морфогенетические факторы. Многие белки синтезируются в печени, жировом теле, а затем транспортируются в яйцеклетку. Яйцеклетка имеет плазматическую мембрану. Во время оплодотворения плазматическая мембрана контролирует поступление многих ионов (например, натрия). К ней прилегает желточная оболочка (гликопротеины – специфическое прикрепление сперматозоида своего вида к соответствующей яйцеклетке), часто прозрачна, яйцеклетка окружена слоем клеток лучистого яйца – фолликулярными питающими клетками. Для оплодотворения сперматозоид должен пройти сквозь все оболочки. Наследственный материал, приносимый яйцеклеткой и сперматозоидом по размеру одинаков
32.Оплодотворение, его фазы, биологическая сущность. Партеногенез. Типы определения пола.
Зрелая яйцеклетка может быть оплодотворена на протяжении небольшого отрезка времени(12-24 часа после овуляции у человека, у рыб и амфибий — через несколько минут после откладки). После овуляции яйцеклетка претерпевает серьезные клеточные изменения. Для оплодотворения очень важен срок жизни. Совокупность изменений яйцеклетки после овуляции – старение яйцеклетки, через 24 часа – перезрелая яйцеклетка – оплодотворение невозможно. Если происходит оплодотворение в данном случае, то невозможна имплантация. Возможны аборты. Ежемесячно созревает только 1 яйцеклетка.
Жизнеспособность сперматозоидов зависит от многих факторов. В щелочной среде они активны, но быстро погибают в кислой среде. Путь от влагалища до 2/3 яйцевода составляет 2-3 часа. В верхней трети влагалища происходит подщелачивание среды (т. к. кислотность высокая) из-за семенной жидкости. В матку проникают только сперматозоиды. Они сохраняют большую подвижность при пониженной температуре. Сперма собирается и замораживается в жидком азоте. Жидко кристаллическая цитоплазма не позволяет образовываться большим кристаллам льда (использование в племенном деле, банки спермы людей). В процессе оплодотворения выделяют 3 этапа.
1) активация яйцеклетки
2) проникновение сперматозоида в яйцеклетку
3) слияние ядер.
В ходе оплодотворения сперматозоид преодолевает все оболочки яйцеклетки. Как только происходит контакт сперматозоида с яйцеклеткой – завершается мейоз яйцеклетки, в т.ч. она выделяет фертилизин. Сперматозоид выделяет антифертилизин – происходит приклеивание. В это время акрозин разрушает оболочки, выделяются ферменты. У человека сперматозоид проникает по-разному: либо только головка, либо шейка и головка. После контакта сперматозоида и яйцеклетки у моноспермных в течение 1-3 минут происходит кортикальная реакция – образуется оболочка оплодотворения. Если несколько сперматозоидов – гибель клетки. Ядро сперматозоида набухает, разрыхляется, образуется мужской пронуклеус. Ядро яйцеклетки превращается в женский пронуклеус. Идет репликация ДНК. Пронуклеусы идут к центру, ядерные оболочки исчезают, происходит слияние пронуклеусов (кариогамия), образуется зигота(2n4c). Через сутки начинается дробление. Возможно развитие эмбриона без участия мужской гаметы (механическое, тепловое, химическое воздействие) – партеногенез. Гиногенез – развитие из яйцеклетки. При гиногенезе сперматозоид проникает и погибает (только женские организмы, т.к. только женский геном). Андрогенез – развитие из сперматозоида. При нм ядро яйцеклетки погибает (только мужской геном). Очень быстро такой геном перерождается в злокачественную опухоль хорионэпителиому.
Типы определения пола.
— прогамный. Пол будущего организма определяется в ходе гаметогенеза у родительских особей.
сингамный. Пол будущего организма определяется в момент слияния половых клеток.
— эпигамный. Пол будущего организма определяется в процессе онтогенеза. У человека имеет место переопределение пола (при патологии) – хотя истинного нет
Партеногенез- развитие без оплодотворения. В случае естественного партеногенеза развитие идет на основе цитоплазмы и пронуклеуса яйцеклетки. Естественный партеногенез- явление редкое, обнаружен у пчел, ос, ряда чешуекрылых. Искусственный партеногенез возможен, по-видимому, у всех животных. На тутовом шелкопряде было показано, что с помощью искусственного партеногенеза можно регулировать соотношение мужского и женского пола в популяции. В активированных яйцах используется информация только женского пронуклеуса — гиногенез. При искусственном партеногенезе можно удалить женский пронуклеус, тогда развитие осуществится за счет мужских пронуклеусов — андрогенез
Источник: StudFiles.net
У высших млекопитающих существует половое размножение, которое происходит с помощью половых клеток. Бесполое (вегетативное) размножение – это размножение без участия половых клеток, в данном случае новый организм развивается из соматических клеток, сохраняющих свойства модифицированных клеток.
Основные отличительные признаки половых клеток от соматических:
1. Половым клеткам на определенном этапе развития свойственен специфический, только им присущий, вид деления – мейоз.
2. В результате мейоза половые клетки сохраняют лишь половинный набор хромосом, характерный для соматических клеток данного вида животного.
3. У половых клеток, по сравнению с соматическими, изменен ядерно-цитоплазматический индекс (соотношение ядра и цитоплазмы).
4. Интенсивный рост женских половых клеток обеспечивается последовательным развитием запаса цитоплазматических компонентов, в т. ч. нуклеиновых кислот и специфического для яйцеклеток включения – желтка.
5. Яйцеклетки окружены более сложными оболочками, чем соматические клетки.
6. У зрелых половых клеток процессы диссимиляции и ассимиляции минимальны, т. е. заторможен обмен веществ.
7. Для большинства сперматозоидов свойственно активное перемещение во внешней среде.
8. Половые клетки – это высоко дифференцированные клетки, выполняющие специфические функции.
Происхождение половых клеток.
В конце XIX в. Вейсман выдвигает теорию о непрерывности зародышевой плазмы. По этой теории существует две плазмы:
1. Соматическая, которая участвует в развитии тела организма;
2. Зародышевая, которая передается непосредственно от половых клеток одного поколения к половым клеткам каждого нового поколения.
Теория Нусс-Баума, которая считает, что существует ранняя неравноценность бластомеров. Из одних бластомеров с ограниченными потенциями развиваются соматические клетки, а другие бластомеры – полипотентные – являются источниками половых клеток. Доказательство этой теории привел Бовери в 1939 г., он обнаружил, что уже первые 4 бластомера яйца лошадиной аскариды неравноценны, и один из бластомеров является родоначальником будущих внутренних органов и половых клеток.
В 1967 – 1968 гг. Вудлен и Мориц подтверждают это положение и доказывают, что этот процесс ранней дифференцировки половых клеток находится под цитоплазматическим контролем.
На свето- и электронно-микроскопическом уровнях доказано, что в одном из бластомеров определяется небольшой участок плотных образований, которые были названы половыми детерминантами, или эктосомами. Предполагается, что эти структуры определяют, или детерминируют, развитие половых клеток в окончательные.
По другой теории, источником половых клеток является герментатический эпителий (эпителий половых желез).
По следующей теории, помимо половых желез существуют в тех или иных местах латентные половые зачатки. ри некотором условии эти резервные гоноциты оказываются способными развиться в гаметы.
Большинство современных исследователей склоняются к мысли, что первичные половые клетки образуются на ранних этапах эмбриогенеза из соматических клеток различных зародышевых листков, но определенных для каждого вида животных.
Гаметогенез.
Появление половых клеток и гаметогенез являются следствием роста, возникает эпигенетически. Их возникновение соответствует особому физиологическому состоянию, которое в норме приобретается развивающимся организмом.
Прогенез – развитие половых клеток – происходит в половых железах. Наступившее гаметогенное физиологическое состояние организма распространяется на все ткани, в особенности на те из них, в которых находятся гоноциты. Сами по себе гоноциты сексуально индифферентны, внутри них не содержится факторов, детерминирующих пол гамет.
Детерминация пола гамет исходит, наоборот, из тканей, в которых гаметогенные клетки находятся и дифференцируются. Эта лабильность половых клеток снижается у более специализированных форм, где пол детерминирован более жестко.
Различают нескольких форм гаметогенеза:
1. Диффузный гаметогенез (у губок).
2. Локализованный гаметогенез. У большинства животных развитие половых клеток происходит в гонадах: в семенниках или яичниках.
3. Солитарный гаметогенез. Развитие и питание половых клеток осуществляется без участия специализированных вспомогательных элементов. Кишечнополостные, черви.
4. Алиментарный гаметогенез. В развитии половых клеток участвуют специализированные клетки, например, трофоциты насекомых, фолликулярный эпителий гонад позвоночных.
Диффузный и локализованный гаметогенез может происходить как по солитарному, так и по алиментарному типу.
Суть гаметогенеза состоит в образовании из диплоидных стволовых предшественников половых клеток, высокодифференцированных клеток – сперматозоидов и яйцеклеток – с гаплоидным набором хромосом.
Развитие мужских половых клеток и сперматогенез.
В результате сперматогенеза возникают клетки – сперматозоиды, содержащие X- и Y-хромосому. Сперматогенез включает 4 стадии:
1. Размножение. Фаза размножения сперматогоний контролируется естественными мутагенами – гормонами аденогипофиза (фоллитропин, интерлекин-1) и гормонами, вырабатываемыми клетками Сертоли и клетками Лейдига. В мужском организме фаза размножения начинается после полового созревания, в репродуктивном возрасте.
2. Рост. Фаза роста выделяется условно в мужском организме, т. к. не связана с накоплением питательных веществ, характерных для овогенеза. Её часто объединяют с третьей фазой созревания в одну – мейотическую – фазу. В мейотической фазе сперматоцит I порядка проходит длинную профазу I деления мейоза, которая у человека продолжается около двух суток. Рост характеризуется увеличением объема сперматоцита в 4 раза.
3. Созревание. В сперматогенезе профаза непосредственно продолжается в последующие стадии I деления мейоза.
4. Формирование. Образование хвостиков.
Развитие женских половых клеток и оогенез.
В результате оогенеза возникают клетки – ооциты (яйцеклетки), содержащие Х-хромосому. Оогенез включает 3 стадии:
1. Размножение. В женском организме фаза размножения инициируется и завершается еще в эмбриогенезе. К окончанию данной фазы в каждом яичнике плода заключено 6 – 7 млн овогоний.
2. Рост. В овогенезе фаза роста более продолжительна и включает в себя малый и большой рост. Малый рост происходит в эмбриогенезе, а большой рост – после полового созревания, в репродуктивном периоде. В фазе роста женские половые клетки увеличивают свои размеры в результате синтеза и накопления желтка (вителлина) – вителлогенез.
3. Созревание. В овогенезе половые клетки останавливаются в стадии диакинеза под влиянием мейозингибирующей субстанции, и могут прибывать в ней несколько лет – стационарная стадия профазы I деления мейоза. Женские половые клетки выходят из стационарного состояния и продолжают своё развитие в разные периоды репродуктивного возраста. Многие из них погибают. Фактором, стимулирующим продолжение мейоза является мейозстимулирующая субстанция, вырабатываемая фолликулами яичников.
В период размножения исходные половые клетки в ходе клеточного цикла синтезируют ДНК и митотически делятся, их размножение протекает на основе общих закономерностей, характерных для соматических клеток.
Фаза созревания – мейоз. Перед вступлением в мейоз диплоидные клетки с генетическим набором 2c2n в синтетическом периоде клеточного цикла увеличивают количество ДНК и, соответственно, количество субъединиц хромосом. Собственно мейоз включает в себя два последовательных деления созревания:
1. Редукционное.
2. Эквационное.
Профаза I деления мейоза в сперматогенезе и оогенезе включает 5 стадий:
1. Лептотена. Реплицированные хромосомы деспирализуются и принимают вид длинных и спутанных нитей.
2. Зиготена. Гомологичные по отцовской и материнской линиям хромосом сближаются, соединяются попарно – конъюгируют, образуются биваленты. При конъюгации контактируют друг с другом строго гомологичные хромосомы.
3. Пахитена. Хромосомы обмениваются геномами и группами генов – кроссинговер.
4. Диплотена. Гомологичные хромосомы, образующие бивалент, отходят друг от друга, но между ними сохраняется связь в области центромеры благодаря хиазмам.
5. Диакинез. Заключительная стадия профазы I деления мейоза, характеризуется спирализацией и укорочением хромосом.
Метафаза I деления мейоза. Тетрады хроматид выстраиваются по экватору. Распределение отцовских и материнских хромосом абсолютно случайно. Исчезает ядерная мембрана. Возникает веретено деления.
В процессе анафазы и телофазы в каждую дочернюю клетку расходится по одной гомологичной двойной хромосоме от каждого бивалента. Это происходит потому, что при I делении мейоза центромера каждого бивалента не расцепляется, и каждый бивалент имеет собственные кинетохоры, которые контактируют с веретеном деления.
Таким образом, каждая дочерняя клетка получает гаплоидный набор хромосом, в связи с чем деление названо редукционным.
II деление мейоза – это мейотическое деление удвоенного набора хромосом.
Источник: megaobuchalka.ru
Световая микроскопия. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
Типы:
· равноплечие, или метацентрические (с центромерой посередине),
· неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов),
· палочковидные, или акроцентрические (с центромерой, расположенной практическина конце хромосомы),
· точковые —очень небольшие, форму которых трудно определить
Совокупность всех структурных и количественных особенностей полного набора хромосом, характерного для клеток конкретного вида живых организмов, называется кариотипом.
Кариотип будущего организма формируется в процессе слияния двух половых клеток (сперматозоида и яйцеклетки). При этом объединяются их хромосомные наборы. Ядро зрелой половой клетки содержит половинный набор хромосом (для человека – 23). Подобный одинарный набор хромосом, аналогичный таковому в половых клетках, называется гаплоидным и обозначается – n. При оплодотворении яйцеклетки сперматозоидом в новом организме воссоздаётся специфический для данного вида кариотип, включающий у человека 46 хромосом. Полный состав хромосом обычной соматической клетки является диплоидным (2n). В диплоидном наборе каждая хромосома имеет аналогичную по размеру и расположению центромеры другую парную хромосому. Такие хромосомы называются гомологичными. Гомологичные хромосомы не только похожи друг на друга, но и содержат гены, отвечающие за одни и те же признаки.
Кариотип женщины в норме содержит две Х-хромосомы, и его можно записать – 46, ХХ. Кариотип мужчины включает Х- и Y-хромосомы (46, ХY). Все остальные 22 пары хромосом получили название аутосомы.
Группы аутосом:
· В группу А входят 3 пары самых длинных хромосом (1, 2, 3-я);
· группа В объединяет 2 пары крупных субметацентрических хромосом (4 и 5-я).
· группа С, включающая 7 пар средних субметацентрических аутосом (с 6-й по 12-ю). По морфологическим особенностям хромосому Х трудно отличить от этой группы.
· Средние акроцентрические хромосомы 13, 14 и 15-й пар входят в группу D.
· Три пары мелких субметацентрических хромосом составляют группу Е (16, 17 и 18-я).
· Самые маленькие метацентрические хромосомы (19 и 20) составляют группу F.
21 и 22-я пары коротких акроцентрических хромосом включены в группу G. Y-хромосома морфологически очень похожа на аутосомы этой группы.
23.Хромосомная теория Т. Моргана.
Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены.
- Материальные носители наследственности – гены находяться в хромосомах, распологаются в них линейно на определенном расстоянии друг от друга.
- Гены, расположенные в одной хромосоме, относятся к одной группе сцепления. Число групп сцепления соответствует гаплоидному числу хромосом.
- Признаки, гены которых находятьс в одной хромосоме, наследуются сцеплено.
- В потомстве гетерозиготных родителей новые сочетания генов, расположенных в дной паре хромосом, могут возникать в результате кроссинговера в процессе мейоза.
- Частота кроссинговера, определяемая по проценту кроссоверных особей, зависит от расстояния между генами.
- На основании линейного расположения генов в хромосоме и частоты кроссинговера как покозателя расстояния между генами можно построить карты хромосом.
Работы Т. Моргана и его сотрудников не только подтвердили значение
хромосом как основных носителей наследственного материала представленного отдельными генами, но и установили линейность расположения их по длине хромосомы.
Доказательством связи материального субстрата наследственности и изменчивости с хромосомами было, с одной стороны, строгое соответствие открытых Г. Менделем закономерностей наследования признаков поведению хромосом в ходе митоза, при мейозе и оплодотворении. С другой стороны, в лаборатории Т. Моргана был обнаружен особый тип наследовани признаков, который хорошо объяснялся связью соответствующих генов с Х хромосомой. Речь идет о сцепленном с полом наследовании окраски глаз у дрозофилы.
Представление о хромосомах как носителях комплексов генов было высказано на основе наблюдения сцепленного наследования ряда родительских признаков друг с другом при передаче их в ряду поколений. Такое сцепление неальтернативных признаков было объяснено размещением соответствующих генов в одной хромосоме, которая представляет собой достаточно устойчивую структуру, сохраняющую состав генов в ряду поколений клеток и организмов.
Согласно хромосомной теории наследственности, совокупность генов,
входящих в состав одной хромосомы, образует группу сцепления. Каждая хромосома уникальна по набору заключенных в ней генов. Число групп сцепления в наследственном материале организмов данного вида определяется, таким образом, количеством хромосом в гаплоидном наборе их половых клеток. При оплодотворении образуется диплоидный набор, в котором каждая группа сцепления представлена двумя вариантами —отцовской и материнской хромосомами, несущими оригинальные наборы аллелей соответствующего комплекса генов.
Представление о линейности расположения генов в каждой хромосоме возникло на основе наблюдения нередко возникающей рекомбинации (взаимообмена) между материнским и отцовским комплексами генов, заключенными в гомологичных хромосомах. Было установлено, что частота
рекомбинации характеризуется определенным постоянством для каждой пары генов в данной группе сцепления и различна для разных пар. Это наблюдение дало возможность высказать предположение о связи частоты рекомбинации с последовательностью расположения генов в хромосоме и процессом кроссинговера, происходящим между гомологами в профазе I мейоза (см. разд. 3.6.2.3).
Представление о линейном распределении генов хорошо объясняло зависимость частоты рекомбинации от расстояния между ними в хромосоме.
Открытие сцепленного наследования неальтернативных признаков легло в основу разработки методики построения генетических карт хромосом с использованием гибридологического метода генетического анализа.
Таким образом, в начале XX в. была неопровержимо доказана роль хромосом как основных носителей наследственного материала в эука-риотической клетке. Подтверждение этому было получено при изучении химического состава хромосом.
24. Деление соматических клеток. Хар-ка фаз митоза.
Деление соматической клетки и ее ядра (митоз) сопровождается сложными многофазными трансформациями хромосом: 1) в процессе митоза происходит удвоение каждой хромосомы на основе комплементарной репликации молекулы ДНК с образованием двух сестринских нитевидных копий (хроматид), соединенных в области центромеры; 2) в последующем сестринские хроматиды разъединяются и эквивалентно распределяются по ядрам дочерних клеток.
В результате в делящихся соматических клетках поддерживается идентичность хромосомного набора и генетического материала.
Отдельно следует сказать о нейронах — высокодифференцированных постмитотических клетках, не претерпевающих клеточных делений на протяжении жизни. Компенсаторные возможности нейронов в ответ на действие повреждающих факторов ограничиваются внутриклеточной регенерацией и репарацией ДНК в неделящемся ядре, чем в значительной степени обусловлена специфика нейропатологических процессов наследственной и ненаследственной природы.
Митоз — сложное деление ядра клетки, биологическое значение которого заключается в точном идентичном распределении дочерних хромосом с содержащейся в них генетической информацией между ядрами дочерних клеток, в результате этого деления ядра дочерних клеток имеют набор хромосом, по количеству и качеству идентичный таковому в материнской клетки.
Хромосомы — основной субстрат наследственности, они — та единственная структура, для которой доказана самостоятельная способность к редупликации. Все другие органоиды клетки, способные к редупликации, осуществляют ее под контролем ядра. В связи с этим важно сохранить постоянство числа хромосом и равномерно распределить их между дочерними клетками, что и достигается всем механизмом митоза. Такой способ деления в клетках растений был открыт в 1874 г. русским ботаником И. Д. Чистяковым а в клетках животных — в 1878 г. русским гистологом П. И. Перемежко (1833—1894).
В процессе митоза (рис. 2.15) последовательно протекает пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза. Эти фазы, непосредственно следующие друг за другом, связаны незаметными переходами. Каждая предыдущая обусловливает переход к последующей.
В клетке, вступающей в деление, хромосомы приобретают вид клубка из множества тонких, слабо спирализованных нитей. В это время каждая хромосома состоит из двух сестринских хроматид. Образование хроматид происходит по матричному принципу в S-период митотического цикла как следствие репликации ДНК.
В самом начале профазы, а иногда и до ее наступления центриоль делится на две, и они расходятся к полюсам ядра. Одновременно хромосомы претерпевают процесс скручивания (спирализации), вследствие чего значительно укорачиваются и утолщаются. Хроматиды несколько отходят друг от друга, оставаясь связанными лишь центромерами. Между хроматидами появляется щель. К концу профазы в животных клетках вокруг центриолей образуется лучистая фигура. В большинстве растительных клеток центриолей нет.
К концу профазы ядрышки исчезают, ядерная оболочка под действием ферментов из лизосом растворяется, хромосомы оказываются погруженными в цитоплазму. Одновременно появляется ахроматиновая фигура, которая состоит из нитей, тянущихся от полюсов клетки (если есть центриоли, то от них). Ахроматиновые нити прикрепляются к центромерам хромосом. Образуется характерная фигура, напоминающая веретено. Электронно-микроскопические исследования показали, что нити веретена — это трубочки,канальцы.
В прометафазе в центре клетки находится цитоплазма, имеющая незначительную вязкость. Погруженные в нее хромосомы направляются к экватору клетки.
В метафазе хромосомы находятся в упорядоченном состоянии в области экватора. Хорошо видны все хромосомы, благодаря чему изучение кариотипов (подсчет числа, изучение форм хромосом) проводится именно в этой стадии. В это время каждая хромосома состоит из двух хроматид, концы которых разошлись. Поэтому на метафазных пластинках (и идиограммах из метафазных хромосом) хромосомы имеют А-образную форму. Изучение хромосом проводится именно в этой стадии.
В анафазекаждая хромосома продольно расщепляется по всей ее длине, в том числе и в области центромеры, точнее сказать, происходит расхождение хроматид, которые после этого становятся сестринскими, или дочерними, хромосомами. Они имеют палочкообразную форму, изогнутую в области первичной перетяжки. Нити веретена сокращаются, направляются к полюсам, а за ними начинают расходиться к полюсам и дочерние хромосомы. Расхождение их осуществляется быстро и всех одновременно, как «по команде». Это хорошо показывают кинокадры делящихся клеток. Бурные процессы происходят и в цитоплазме, которая на кинопленке напоминает кипящую жидкость.
В телофазе дочерние хромосомы достигают полюсов. После этого хромосомы деспирализуются, теряют ясные очертания, вокруг них формируются ядерные оболочки. Ядро приобретает строение, сходное с интерфазным материнской клетки. Восстанавливается ядрышко.
Далее происходит цитокинез т. е. разделение цитоплазмы.
25. Половые клетки человека, их строение. Типы строения яйцеклеток.
Для участия в половом размножении в родительских организмах вырабатываются гаметы —клетки, специализированные к обеспечению генеративной функции.
Слияние материнской и отцовской гамет приводит к
возникновению зиготы — клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития.
У некоторых организмов зигота образуется в результате объединения гамет, неотличимых по строению. В таких случаях говорят об изогамии.
У большинства видов по структурным и функциональным признакам половые клетки делятся на материнские (яйцеклетки) и отцовские (сперматозоиды). Как правило, яйцеклетки и сперматозоиды вырабатываются разными организмами — женскими (самки) и мужскими (самцы). В подразделении гамет на яйцеклетки и сперматозоиды, а особей на самок и самцов заключается явление полового диморфизма (рис. 5.1; 5.2). Наличие его в природе отражает различия в задачах, решаемых в процессе полового размножения мужской или женской гаметой, самцом или самкой.
Мужские половые клетки человека — сперматозоиды, или спермии, длиной около 70 мкм, имеют головку, шейку и хвост.
Сперматозоид покрыт цитолеммой, которая в переднем отделе содержит рецептор —обеспечивающий узнавание рецепторов яйцеклетки.
Головка сперматозоида включает небольшое плотное ядро с гаплоидным набором хромосом. Передняя половина ядра покрыта плоским мешочком, составляющим чехлик сперматозоида. В нем располагается акросома (от греч. асго — верхушка, soma — тело),
состоящая из видоизмененного комплекса Гольджи. Акросома содержит набор ферментов. В ядре сперматозоида человека, занимающего
основную массу головки, содержится 23 хромосомы, одна из которых является половой (X или Y), остальные — аутосомами. Хвостовой отдел сперматозоида состоит из промежуточной, главной и терминальной частей.
При исследовании сперматозоонов под электронным микроскопом обнаружено, что протоплазма головки его имеет не коллоидное, а жидкокристаллическое состояние. Этим достигается устойчивость сперматозоонов к неблагоприятным влияниям внешней среды. Например, они в меньшей степени повреждаются ионизирующей радиацией по сравнению с незрелыми половыми клетками.
Все сперматозооны несут одноименный (отрицательный) электрический заряд, что препятствует их склеиванию.
Человек выделяет около 200 млн. сперматозоидов
Яйцеклетки, или овоциты(от лат. ovum — яйцо), созревают в неизмеримо меньшем количестве, чем сперматозоиды. У женщины в течение полового цикла 24—28 дней) созревает, как правило, одна яйцеклетка. Таким образом, за детородный период образуются около 400 зрелых яйцеклеток.
Выход овоцита из яичника называется овуляцией. Вышедший из яичника овоцит окружен венцом фолликулярных клеток, число которых достигает 3—4 тыс. Он подхватывается бахромками маточной трубы (яйцевода) и продвигается по ней. Здесь заканчивается созревание половой клетки. Яйцеклетка имеет шаровидную форму, больший, чем у спермия, объем цитоплазмы, не обладает способностью самостоятельно передвигаться.
Строение. Яйцеклетка человека имеет диаметр около 130 мкм. К цитолемме прилежат блестящая, или прозрачная, зона и далее слой фолликулярных клеток. Ядро женской половой клетки имеет гаплоидный набор хромосом с X-половой хромосомой, хорошо выраженное ядрышко, в кариолемме много поровых комплексов. В период роста ооцита в ядре происходят интенсивные процессы синтеза иРНК, рРНК.
В цитоплазме развиты аппарат синтеза белка (эндоплазматическая сеть, рибосомы) и аппарат Гольджи. Количество митохондрий умеренно, они расположены около желточного ядра, где идет интенсивный синтез желтка, клеточный центр отсутствует. Аппарат Гольджи на ранних стадиях развития располагается около ядра, а в процессе созревания яйцеклетки смещается на периферию цитоплазмы.
Яйцеклетки покрыты которые выполняют защитную функцию, обеспечивают необходимый тип обмена веществ, у плацентарных млекопитающих служат для внедрения зародыша в стенку матки, а также выполняют и другие функции.
Цитолемма яйцеклетки имеет микроворсинки, располагающиеся между отростками фолликулярных клеток. Фолликулярные клетки выполняют трофическую и защитную функции.
Яйцеклетки значительно крупнее, чем соматические клетки. Внутриклеточная структура цитоплазмы в них специфична для каждого вида животных, чем обеспечиваются видовые (а нередко, и индивидуальные) особенности развития. В яйцеклетках содержится ряд веществ, необходимых для развития зародыша. К их числу относится питательный материал (желток).
Классификация яйцеклеток основывается на признаках наличия, количества и распределения желтка (lecithos), представляющего собой белково-липидное включение в цитоплазме, используемое для питания зародыша.
Различают безжелтковые (алецитальные), маложелтковые (олиголецитальные), среднежелтковые (мезолецитальные), многожелтковые (полилецитальные) яйцеклетки.
У человека наличие малого количества желтка в яйцеклетке обусловлено развитием зародыша в организме матери.
Полярность яйцеклеток.При малом количестве желтка в яйцеклетке он обычно распределен в цитоплазме равномерно и ядро располагается примерно в центре. Такие яйцеклетки называют изолецитальными (от греч. изос —равный). У большинства позвоночных желтка много, и он распределен в цитоплазме яйцеклетки неравномерно. Это анизолециталъные клетки. Основная масса желтка скапливается у одного из полюсов клетки — вегетативного полюса. Такие яйцеклетки называют телолецйтальными (от греч. телос — конец). Противоположный полюс, к которому оттесняется свободная от желтка активная цитоплазма, называют анималъным. Если желток все же погружен в цитоплазму и не обособлен от нее в виде отдельной фракции, как у осетровых и земноводных, яйцеклетки называют умеренно телолецитальными. Если желток полностью отделен от цитоплазмы, как у амниот, то это резко телолецитальные яйцеклетки.
26.Репродукция живого. Классификация способов размножения.
Размножение, или репродукция,— одно из основных свойств, характеризующих жизнь. Под размножением понимается способность организмов производить себе подобных. Явление размножения тесно связано с одной из черт, характеризующих жизнь,— дискретностью. Как известно, целостный организм состоит из дискретных единиц — клеток. Жизнь почти всех клеток короче жизни особи, поэтому существование каждой особи поддерживается размножением клеток. Каждый вид организмов также дискретен, т. е. состоит из отдельных особей. Каждая из них смертна. Существование вида поддерживается размножением (репродукцией) особей. Следовательно, размножение — необходимое условие существования вида и преемственности последовательных генераций внутри вида. В основе классификации форм размножения лежит тип деления клеток: митотический (бесполое) и мейотический (половое). Формы размножения можно представить в виде следующей схемы

Бесполое размножение. У одноклеточных эукариот это — деление, в основе которого лежит митоз, у прокариот — разделение нуклеоида, а у многоклеточных организмов — вегетативное (лат. vegetatio — расти) размножение, т. е. частями тела или группой соматических клеток.
Бесполое размножение одноклеточных организмов. У одноклеточных растений и животных различают следующие формы бесполого размножения: деление, эндогония, множественное деление (шизогония) и почкование.
Деление характерно для одноклеточных (амебы, жгутиковые, инфузории). Сначала происходит митоти-ческое деление ядра, а затем в цитоплазме возникает все углубляющаяся перетяжка. При этом дочерние клетки получают равное количество информации. Органоиды обычно распределяются равномерно. В ряде случаев обнаружено, что делению предшествует их удвоение. После деления дочерние особи растут и, достигнув величины материнского организма, переходят к новому делению.
Эндогония — внутреннее почкование. При образовании двух дочерних особей — эндодиогонии — материнская дает лишь двух потомков (так происходит размножение токсоплаз-мы), но может быть множественное внутреннее почкование, что приведет к шизогонии.
Шизогония, или множественное деление,— форма размножения, развившаяся из предыдущей. Она тоже встречается у одноклеточных организмов, например у возбудителя малярии — малярийного плазмодия. При шизогонии происходит многократное деление ядра без цитокинеза, а затем и вся цитоплазма разделяется на частички, обособляющиеся вокруг ядер. Из одной клетки образуется много дочерних. Эта форма размножения обычно чередуется с половой.
Почкование заключается в том, что на материнской клетке первоначально образуется небольшой бугорок, содержащий дочернее ядро, или нуклеоид. Почка растет, достигает размеров материнской особи и затем отделяется от нее. Эта форма размножения наблюдается у бактерий, дрожжевых грибов, а из одноклеточных животных — у сосущих инфузорий.
Спорообразование встречается у животных, относящихся к типу простейших, классу споровиков. Спора — одна из стадий жизненного цикла, служащая для размножения, она состоит из клетки, покрытой оболочкой, защищающей от неблагоприятных условий внешней среды. Некоторые бактерии после полового процесса способны образовывать споры. Споры бактерий служат не для размножения, а для переживания неблагоприятных условий и по своему биологическому значению отличаются от спор простейших и многоклеточных растений.
Вегетативное размножение многоклеточных ж-ных При вегетативном размножении у многоклеточных животных новый организм образуется из группы клеток, отделяющейся от материнского организма. Вегетативное размножение встречается лишь у наиболее примитивных из многоклеточных животных: губок, некоторых кишечнополостных, плоских и кольчатых червей.
У губок и гидры за счет размножения группы клеток на теле образуются выпячивания (почки). В почку входят клетки экто- и энтодермы. У гидры почка постепенно увеличивается, на ней формируются щупальца, и, наконец, она отделяется от материнской особи. Ресничные и кольчатые черви делятся перетяжками на несколько частей; в каждой из них восстанавливаются недостающие органы. Так может образоваться цепочка особей. У некоторых кишечнополостных встречается размножение стробиляцией, заключающейся в том, что полиплоидный организм довольно интенсивно растет и по достижении известных размеров начинает поперечными перетяжками делиться на дочерние особи. В это время полип напоминает стопку тарелок. Образовавшиеся особи — медузы отрываются и начинают самостоятельную жизнь. У многих видов (например, кишечнополостных) вегетативная форма размножения чередуется с половой.
Особой формой вегетативного размножения следует признать полиэмбрионию, при которой эмбрион делится на несколько частей, каждая из которых развивается в самостоятельный организм. Полиэмбриония распространена у ос (наездники), ведущих паразитический образ жизни в личиночном состоянии, из млекопитающих — у броненосца. К этой категории явлений относится образование однозиготных близнецов у человека и других млекопитающих.
Половое размножение
Половой процесс. Половое размножение отличается наличием полового процесса, который обеспечивает обмен наследственной информацией и создает условия для возникновения наследственной изменчивости. В нем, как правило, участвуют две особи — женская и мужская, которые образуют гаплоидные женские и мужские половые клетки — гаметы. В результате оплодотворения, т. е. слияния женской и мужской гамет, образуется диплоидная зигота с новой комбинацией наследственных признаков, которая и становится родоначальницей нового организма.
Половое размножение по сравнению с бесполым обеспечивает появление наследственно более разнообразного потомства. Формами полового процесса являются конъюгация и копуляция.
Конъюгация — своеобразная форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, образуемому двумя особями. При конъюгации обычно не происходит увеличения количества особей, но происходит обмен генетическим материалом между клетками, что обеспечивает перекомбинацию наследственных свойств. Конъюгация типична для ресничных простейших (например, инфузорий), некоторых водорослей (спирогиры).
Копуляция (гаметогамия) — форма полового процесса, при которой две различающиеся по полу клетки — гаметы — сливаются и образуют зиготу. При этом ядра гамет образуют одно ядро зиготы.
Различают следующие основные формы гаметогамии: изогамия, анизогамия и оогамия.
При изогамии образуются подвижные, морфологически одинаковые гаметы, однако физиологически они различаются на «мужскую» и «женскую». Изогамия встречается у многих водорослей.
При анизогамии (гетерогамии) формируются подвижные, различающиеся морфологически и физиологически гаметы. Такой тип полового процесса характерен для многих водорослей.
В случае оогамии гаметы сильно отличаются друг от друга. Женская гамета — крупная неподвижная яйцеклетка, содержащая большой запас питательных веществ. Мужские гаметы — сперматозоиды — мелкие, чаще всего подвижные клетки, которые перемещаются с помощью одного или нескольких жгутиков. У семенных растений мужские гаметы — спермии — не имеют жгутиков и доставляются к яйцеклетке с помощью пыльцевой трубки. Оогамия характерна для животных, высших растений и многих грибов.
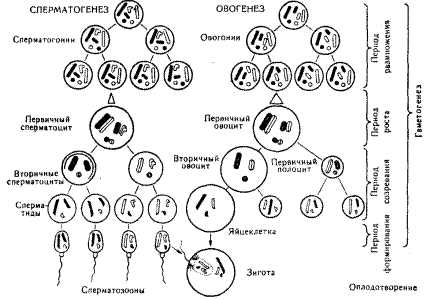
27. Овогенез и сперматогенез.
Сперматогенез.Семенник состоит из многочисленных канальцев. На поперечном разрезе через каналец видно, что в нем имеется несколько слоев клеток. Они пред ставляют собой последовательные стадии развития сперматозоонов.
Наружный слой (зона размножения) составляют сперматогонии — клетки округлой формы; у них относительно большое ядро и значительное количество цитоплазмы. В период эмбрионального развития и после рождения до полового созревания сперматогонии делятся путем митоза, благодаря чему увеличиваются число этих клеток и сам семенник. Период интенсивного деления называется периодом размножения
После наступления половой зрелости часть сперматогониев также продолжает делиться митотически и образовывать такие же клетки, но некоторые из них перемещаются в следующую зону роста, расположенную ближе к просвету канальца. Здесь происходит значительное возрастание размеров клеток за счет увеличения количества цитоплазмы. В этой стадии они называются первичными сперматоцитами.
Третий период развития мужских гамет называется периодом созревания. В этот период происходят два быстро наступающих одно вслед за другим деления. Из каждого первичного сперматоцита сначала образуются два вторичных сперматоцита, а затем четыре сперматиды, имеющие овальную форму и значительно меньшие размеры. Деление клеток во время периода созревания сопровождается перестройкой хромосомного аппарата (происходит мейоз; см. ниже). Сперматиды перемещаются в зону, ближайшую к просвету канальцев, где из них формируются сперматозооны.
У большинства диких животных сперматогенез происходит лишь в определенные периоды года. В промежутках между ними в канальцах семенни ков содержатся лишь сперматогонии. Но у человека и большинства домашних животных сперматогенез происхо дит в течение всего года.
Овогенез. Фазы овогенеза сопоставимы с таковыми при сперматогенезе. В этом процессе также имеется период размножения, когда интенсивно делятся овогонии — мелкие клетки с относительно крупным ядром и небольшим количеством цитоплазмы. У млеко питающих и человека этот период заканчивается еще до рождения. Сформировавшиеся к этому времени первичные овоциты сохраняются далее без изменений многие годы. С наступлением половой зрелости периодически отдельные овоциты вступают в период роста клетки, увеличиваются, в них накапливаются желток, жир, пигменты.
В цитоплазме клетки, ее органоидах и мембранах происходят сложные морфологические и биохимические преобразования. Каждый овоцит окружается мелкими фоликулярными клетками, обеспечивающими его питание.
Далее наступает период созревания. в процессе которого происходят два последовательных деления, связанных с преобразованием хромосомного аппарата (мейоз). Кроме того, эти деления сопровождаются неравномерным разделением цитоплазмы между дочерними клетками. При делении первичного овоцита образуется одна круп ная клетка — вторичный овоцит, содержащая почти вся цитоплазму, и маленькая клетка, получившая название первичного полоцита. При втором делении созревания цитоплазма снова распределяется неравномерно. Образуется один крупный вторичный овоцит и вторичный полоцит. В это время первичный полоцит также может разделиться на две клет ки. Таким образом, из одного первичного овоцита образуются один вторичный овоцит и три полоцита (редукционные тельца). Далее из вторичного овоцита формируется яйцеклетка, а полоциты рассасываются или сохраняются на поверхности яйца, но не принимают участия в дальнейшем развитии. Неравномерное распределе ние цитоплазмы обеспечивает яйце клетке получение значительного коли чества цитоплазмы и питательных веществ, которые потребуются в будущем для развития зародыша.
У млекопитающих и человека периоды размножения и роста яйцеклеток проходят в фолликулах (рис. 3.5). Зрелый фолликул заполнен жидкостью, внутри него находится яйце клетка. Во время овуляции стенка фолликула лопается, яйцеклетка попадает в брюшную полость, а затем, как правило, в маточные трубы. Период созревания яйцеклеток протекает в трубах, здесь же происходит оплодотворение.
У многих животных овогенез и созревание яйцеклеток совершаются лишь в определенные сезоны года. У женщин обычно ежемесячно созре вает одна яйцеклетка, а за весь период половой зрелости — около 400. Для человека имеет существенное значение тот факт, что первичные овоциты фор мируются еще до рождения и затем сохраняются всю жизнь и лишь по степенно некоторые из них начинают переходить к созреванию и дают яйце клетки. Это значит, что различные не благоприятные факторы, которым под вергается в течение жизни женский организм, могут сказаться на их даль нейшем развитии; ядовитые вещества (в том числе никотин и алкоголь), попадающие в организм, могут никнуть в овоциты и в дальнейшем зызвать нарушения нормального развития будущего потомства.
28. Митоз, его биологическое значение.
Важнейшим компонентом клеточного цикла является митотический (пролиферативный) цикл. Он представляет собой комплекс взаимосвязанных и согласованных явлений во время деления клетки, а также до и после него. Митотический цикл — это совокупность процессов, происходящих в клетке от одного деления до следующего и заканчивающихся образованием двух клеток следующей генерации. Кроме этого, в понятие жизненного цикла входят также период выполнения клеткой своих функций и периоды покоя. В это время дальнейшая клеточная судьба неопределенна: клетка может начать делиться (вступает в митоз) либо начать готовиться к выполнению специфических функций.
Источник: helpiks.org
Сперматозоиды животных
Длина сперматозоидов животных различна, но в основном это мелкие клетки. Их длина от размеров животного не зависит. Сперматозоид морской свинки имеет длину 100 мкм, быка – 65 мкм, крокодила – 20 мкм, человека – 53 мкм.
В половые протоки добавочными железами выделяется жидкость. Вместе жидкость и сперматозоиды образуют сперму. В состав спермы могут входить лейкоциты и эпителиальные клетки. Количество выбрасываемых сперматозоидов животными очень велико, что выработалось в процессе эволюции как приспособление к более успешному оплодотворению яйцеклетки. Например, у человека, в 1 см3 спермы – содержится 60 млн. сперматозоидов. Активность их обуславливается энергией, которая продуцируется в митохондриях средней части хвостика. Наибольшая подвижность сперматозоидов наблюдается в нейтральной среде при температуре 35оС.
Продолжительность жизни сперматозоидов незначительна и различна у разных животных, например, у рыб сперматозоиды живут несколько минут, в половых путях свиньи они сохраняют активность в течение 22–30 часов, у овцы – 36 часов. Во влагалище женщины сперматозоиды активны в течение 2,5 часа, но в матке сохраняют жизнеспособность до 48 часов. Однако у многих насекомых, например, у пчел, самки имеют семяприемники, в которых сперматозоиды могут сохраняться в течение нескольких лет.
Вне организма, регулируя условия, сперматозоиды можно сохранять сколько угодно долго. Это имеет особенно большое значение в животноводческой практике, в мероприятиях по искусственному осеменению, когда сперму можно сохранять в течение длительных сроков. Сперматозоиды обладают реакцией реотаксиса, т.е. передвижения против тока жидкости. У животных с внешним оплодотворением поступательное движение осуществляется по спирали, а при внутреннем – прямолинейно.
Типы яйцеклеток
Яйцо или яйцеклетка – это специально дифференцированная клетка, приспособленная к оплодотворению и дальнейшему развитию. В отличие от сперматозоидов яйцеклетки не способны к активному движению и имеют однообразную форму: у большинства животных они округлые, могут быть овальные или вытянутые. Ядро, как правило, повторяет форму яйцеклетки. Для нее характерно большое количество цитоплазмы, в которой, помимо обычных органоидов, содержится большое количество желтка – запасного питательного материала для развития зародыша. Яйцеклетки с большим количеством желтка, как правило, больших размеров (рыбы, рептилии, птицы), яйцеклетки с малым количеством желтка (ланцетник) или не содержащие вообще (млекопитающие) не больших размеров, но всегда крупнее сперматозоидов. Строение яиц определяется содержанием и местоположением желтка. По этим признакам можно выделить следующие типы яйцеклеток. Алецитальные яйцеклетки вообще не содержат желтка. Такие яйцеклетки характерны для плацентарных млекопитающих. Гомолецитальные яйцеклетки содержат небольшое количество желтка, более или менее равномерно распределенного по всей цитоплазме (ланцетник). Следующий тип – телолецитальные. Они характеризуются содержанием среднего или большого количества желтка, расположенного полярно. Этот тип подразделяется на два подтипа: «средне» телолецитальный и «крайне» телолецитальный. «Средне» телолецитальные яйцеклетки содержат среднее количество желтка, распложенного в вегетативной части (земноводные). «Крайне» телолецитальный тип содержит большое количество желтка также сконцентрированного в вегетативной части (костистые рыбы, рептилии, птицы). Центролецитальный тип яйцеклетки также характеризуется наличием большого количества желтка, который расположен в центре яйцеклетки (насекомые).
Наличие большого количества желтка обуславливает полярность яиц (исключение – центролецитальные клетки). Полярность яиц хорошо выражена у земноводных, рептилий, птиц. Верхняя часть яйца, бедная желтком, называется анимальным полюсом, а нижняя, содержащая большое количество желтка, – вегетативным. Мысленная линия соединяющая анимальный и вегетативный полюсы и проходящая через центр яйцеклетки, называется осью яйца.
Характерной особенностью для строения яйцеклеток является наличие у них оболочек. Оболочки сохраняют форму и строение яйца, предохраняют его содержимое от высыхания, защищают от механических и химических воздействий внешней среды.
Оболочки яйцеклетки
Оболочки яйцеклеток подразделяют на три группы: первичные, вторичные и третичные.
Первичная оболочка яйцеклетки образуется самим яйцом и представляет собой ее поверхностный уплотненный слой, ее называют желточной оболочкой и образуется она до оплодотворения в процессе оогенеза.
Вторичные оболочки вырабатываются клетками, питающими яйцо. Примером могут служить фолликулярные клетки. Часто эти оболочки могут быть плотными и тогда у них имеются микропили – отверстия для проникновения сперматозоида.
Третичные оболочки служат для защиты яйца, они образуются во время прохождения яйцеклетки по яйцеводу. Примером третичных оболочек могут служить белковая, подскорлуповые и скорлуповая у птиц.
Яйцеклетки очень чувствительны к колебаниям температуры, ультрафиолетовым лучам, лучам Рентгена и радия.
При сравнительно небольшом повышении температуры, которое животные переносят безболезненно, яйцеклетки погибают. Повышение дозировки лучей Рентгена, радия, ультрафиолетовых лучей смертельно для яйцеклеток. Установлено, что если развитие и оплодотворение половых клеток ещё молодое, то оно более чувствительно к облучению.
Источник: kto.guru