В хлоропластах растительных клеток происходит
ГЛАВА 3. ПЛАСТИДЫ Содержание Хлоропласты Лейкопласты Хромопласты Онтогенез и взаимопревращения пластид Происхождение пластид и митохондрий Учебные задачи: 1. Выяснить особенности строения пластид. Узнать об особенно- стях внутреннего строения, составе пигментов, функционировании (фото- синтезе, образовании первичного крахмала и т.д.) хлоропластов. 2. Сравнить характеристики хромопластов и лейкопластов со свой- ствами хлоропластов (особенности расположения в теле растения, внут- ренняя структура, пигментный состав и функции). 3. Выяснить возможные взаимные превращения пластид и процесс трансформации пластид в онтогенезе растения. 4. Уяснить основные представления симбиотической теории происхо- ждения пластид и митохондрий. Глоссарий Амилопласты – лейкопласты, в которых откладывается запас- ной крахмал. Вторичный крахмал – крахмал, образующийся из продуктов гид- ролиза первичного крахмала. Граны – стопки тилакоидов. Каротин – разновидность каротиноидов оранжево-красного цве- та.
аротиноиды – жирорастворимые пигменты желтых, оранже- вых, красных цветов. Лейкопласты – бесцветные пластиды. Люмен – полость тилакоида. Первичный (ассимиляционный) крахмал – крахмал, образующийся и временно откладывающийся в хлоропластах. Пластидом – совокупность пластид клетки. Пластиды – двумембранные органеллы растений, содержащие пигменты и выполняющие функции синтеза и запасания органических веществ. Пластоглобулы – капли масла в строме пластид. Проламеллярное тело – предшественник ламеллярной системы в пропластидах. Пропластиды – недифференцированные предшественники пла- стид (встречающиеся в меристематических и эмбриональных тка- нях). 31 Протеинопласты (протеопласты) – лейкопласты, в которых откладывается запасной белок. Строма – состоящий из белковых коллоидов матрикс пластид. Тилакоиды (ламеллы) – пигментосодержащие мембранные «ме- шочки», находящиеся в строме пластид. Фикобилины – синие и красные пигменты хлоропластов некото- рых водорослей. Фотосинтез – синтез органических веществ из углекислого газа и воды за счет энергии света, в процессе которого выделяется кисло- род. Хлоропласты – пластиды растений зеленого цвета, выполняю- щие функцию фотосинтеза. Хлорофиллы – пигменты зеленого цвета, улавливающие энергию света. Хромопласты – пластиды с желтой или оранжево-красной окра- ской. Элайопласты (олеопласты) – лейкопласты, в которых отклады- вается запасные масла. Этиоляция – утрата хлоропластами структуры и пигментов в случае недостатка освещения.
тиопласты – хлоропласты с неразвитой системой тилакоидов и низким содержанием пигментов вследствие недостатка освещения. Наряду с вакуолями и клеточной оболочкой пластиды – важнейшие компоненты растительных клеток. Пластиды (греч. plástides — создающие, образующие, от plastós — вылепленный, оформленный) - органеллы цито- плазмы автотрофных растений, содержащие пигменты и осуществляющие синтез органических веществ. Совокупность всех пластид клетки называют пластидóм. Обычно они хорошо видимы под световым микроскопом. Каж- дая пластида окружена собственной оболочкой, состоящей из двух мембран. Внутри пластид различают мембранную систему и более или менее гомо- генное вещество – строму (рис. 14). Зрелые пластиды классифицируют на основании их окраски. По этому признаку различают три типа пластид: хло- ропласты, лейкопласты, хромопласты. Обычно в клетке встречаются пласти- ды только одного типа. 3.1. Хлоропласты Общая характеристика. Хлоропласты (от греч. chlërós – зеленый и plastós – вылепленный, оформленный) – важнейшие пластиды. Это центры фотосинтетической активности, в которых сосредоточен весь хлорофилл и все вспомогательные пигменты, связанные с фотосинтезом. У высших растений хлоропласты по форме напоминают двояковыпук- лую линзу. Форма эта довольно постоянна. Их диаметр равен 5-8 мкм, а толщина – около 1 мкм. В клетке имеется 15-50 хлоропластов. Число хло- 32 ропластов в клетках связано с их величиной: чем мельче хлоропласты, тем больше их в клетке.
водорослей форма, число и размеры хлоропластов очень разнообразны. Величина и форма хлоропластов зависят также от внешних условий. Обычно у тенелюбивых растений хлоропласты крупнее и богаче хлорофил- лом, чем у светолюбивых. В клетках высших растений хлоропласты расположены таким обра- зом, что плоская поверхность их обращена к клеточной оболочке. Особенно много их около межклетников, заполненных воздухом. Положение хлоропластов в клетке может меняться в зависимости от освещенности. Они располагаются так, что улавливают свет наилучшим об- разом, не подвергаясь действию прямых солнечных лучей. На рассеянном свету хлоропласты сосредоточиваются у поверхностной стенки клетки; на ярком свету – перемещаются на боковые стенки или поворачиваются к лучам ребром. Хлоропласты встречаются почти во всех клетках надземных органов растений, куда проникает свет. Но особенно много их в листьях и незрелых плодах. Лишь немногие типы клеток освещенных частей растений вместо хлоропластов содержат лейкопласты или хромопласты. Это гаметы, выдели- тельные клетки, проводящие элементы луба, первичные покровные ткани. В клетках корней хлоропластов, за немногими исключениями, нет. По химическому составу хлоропласты несколько отличаются от ос- тальной цитоплазмы. Так, содержание липоидов в них составляет 20-40% от сухой массы, тогда как в цитоплазме липоидов всего 2-3%. Структурной основой хлоропластов являются белки – около 50% сухой массы. Содержа- ние хлорофилла составляет 5-10%, каротиноидов – 1-2%.
к и в митохон- дриях, в хлоропластах имеется немного РНК (0,5-3,5%). Содержание ДНК ещё меньше. Строение хлоропластов (рис. 14, 15) сходно у разных растений. Каж- дый хлоропласт окружен двойной мембраной и имеет сложную внутреннюю систему мембран. Мембраны оболочки агранулярные (отсутствуют заметные белковые тельца АТФазы). Они обладают избирательной проницаемостью. Основная структурная единица хлоропласта – тилакоид (ламелла). Тилако- ид представляет собой тонкий плоский мешочек, ограниченный однослой- ной мембраной. В нем находятся хлорофилл, вспомогательные пигменты и ферменты, принимающие участие в фотохимических реакциях фотосинтеза. Полость тилакоида называется люмен. Тилакоиды собраны в группы напо- добие стопки монет. Эти стопки называются гранами. В гранах тилакоиды располагаются параллельно друг другу, соприкасаясь мембранами. Отдель- ные граны связаны между собой в единую систему с помощью тилакоидов, пронизывающих межгранные пространства. Эти межгранные тилакоиды, в отличие от тилакоидов гран, обычно не параллельны, удалены друг от друга на разные расстояния, имеют разный диаметр. Сгруппированные в граны и одиночные тилакоиды не являются от- дельными компартментами хлоропласта, а представляют собой непрерывное образование (пространственный континуум) с многочисленными мембран- 33 ными наложениями. В некоторых участках хлоропластов имеются складки внутренней мембраны пластидной оболочки. Эти складки пере- ходят в межгранные тилакоиды.
такие складки встречаются редко. У хлоропластов водорослей и в немногих типах клеток выс- ших растений типичные граны не образуют- ся. Число тилакоидов в гранах колеблет- ся от двух до нескольких десятков. Их диа- метр около 0,5 мкм. Потому они не видны в световой микроскоп. Число и расположение гран зависят от вида растения, возраста и активности хлоропластов. У светолюбивых растений граны мельче, чем у тенелюби- вых. Все пространство между гранами за- полнено бесцветной стромой. В строме на- ходятся рибосомы, светлые зоны с нитями Рис. 14. Строение хлоропласта ДНК, изредка – крахмальные зерна, белко- (а), лейкопласта (б), амилопласта вые кристаллы. Кроме того, в строме со- (в) и хромопласта (г): 1 – внешняя держатся многие ферменты, участвующие в мембрана, 2 – внутренняя темновых реакциях фотосинтеза (фиксации мембрана, 3 – строма, 4 СО2). – тилакоиды стромы, 5 – грана, При таком строении значительно уве- 6 – тилакоид граны, 7 – зерно личивается фотоактивная поверхность хло- первичного крахмала, 8 – липидная ропласта и обеспечивается максимальное капля (пластоглобула) с использование световой энергии. пигментами Содержащиеся в хлоропластах фер- менты кодируются либо ядерной, либо хлоропластной ДНК. Некоторые фер- менты состоят из двух белковых субъединиц. Причем одна из них кодируется ядерной ДНК, а другая – ДНК хлоропласта (например, рибулозобисфосфат- карбоксилаза (РУБИСКО)). В хлоропластах содержатся также рибосомы (сходные по структуре с рибосомами бактерий), РНК, аминокислоты и ферменты, необходимые для синтеза белка.
аче говоря, в них имеется собственная система синтеза бел- ка. Таким образом, хлоропласты обладают некоторой автономностью. Установлено, что большинство белков мембран тилакоидов синтези- руется в рибосомах хлоропластов. Напротив, большинство белков стромы и липиды мембран образуются вне хлоропластов. Хлоропласты способны также к синтезу и разрушению полисахаридов (крахмала), аминокислот, ли- пидов. Пигменты хлоропластов. Хлорофилл – основной пигмент, участ- вующий в фотосинтезе. Он встречается в нескольких формах: у высших 34 растений в основном хлорофилл а и хлорофилл b. Эти формы хлорофилла немного отличаются одна от другой по спектрам поглощения. Кроме того, в хлоропластах большинства высших растений присутствуют каротиноиды – желтый ксантофилл и красно-оранжевый каротин. Обычно эти пигменты маскируются хлорофиллом и незаметны на протяжении почти всего периода вегетации. Осенью, когда концентрация хлорофилла в стареющих листьях снижается, каротиноиды становятся хорошо заметными. Осенняя окраска листьев зависит в основном от них. Правда, на окраску осенних листьев влияют также антоцианы, присутствующие в вакуолях клеток. У некоторых водорослей хлоропласты особенно богаты фикобилинами – синим и крас- ным пигментами. Каротиноиды защищают хлорофилл от разрушающего действия молекулярного кислорода. Кроме того, они повышают продук- тивность фотосинтеза, поглощая и передавая хлорофиллу энергию тех длин волн, которые хлорофиллом не поглощаются (желто-зеленую часть спектра).
результате и эта энергия может использоваться для фотосинтеза. При содержании растений в темноте они приобретают бледную желто- ватую окраску. Это вызвано тем, что синтез хлорофилла происходит только на свету. Хлоропласты таких растений содержат очень мало хлорофилла и имеют слабо развитую сеть тилакоидов. Растения, выращенные при недос- татке света или в темноте, называют этиолированными. Хлоропласты таких растений называются этиопласты. Фотосинтез. В мембране тилакоидов осуществляется первичная световая стадия фо- тосинтеза. Смысл ее заключается в фиксации и запасании энергии света в энергии химиче- ских связей: АТФ и НАДФ·Н (восстановленно- го никотинамид-адениндинуклеотидфосфата), необходимых для ассимиляции CO2. У растений АТФ образуется в ос- новном за счет энергии света в хлоропластах. Поэтому значение митохондрий у растений намного меньше, чем у животных. Источником энергии для образования молекул АТФ является разность потенциалов, которая образуется на мембране в результате направленного переноса электронов. При этом происходит так называемый фотолиз воды. Под действием света переносчики электронов в мембране отбирают электроны у воды: при Рис. 15. Строение граны этом образуется молекулярный кислород (O2) хлоропласта: а – объемное и накапливаются протоны (Н+) – положитель- изображение, б – схема разреза: 1 – тилакоид граны, но заряженные ионы.
электроны тем вре- 2 – межгранный тилакоид, 3 менем уходят на другую сторону мембраны и – участок спаренных мембран захватываются НАДФ, и в результате образу- ется НАДФ– – отрицательно заряженная мо- 35 лекула. Благодаря мембране, играющей роль барьера, осуществляется про- странственное разобщение положительно заряженных ионов и отрицательно заряженных молекул, т.е. на мембране накапливается электро-химический потенциал. Наружная поверхность тилакоида гранулярная – покрыта белко- выми частицами диаметром 14-15 нм, которые представляют собой «фактор сопряжения», фермент АТФазу, имеющий канал. В итоге мембрана разряжа- ется: протоны проходят через канал АТФазы, и энергия этого потока запаса- ется в форме синтезируемой АТФ. Разноименно заряженные ион и молекула взаимодействуют и образуется вышеупомянутый НАДФ·Н. В строме же сосредоточены ферменты фиксации CO2, т.е. происходит темновая стадия фотосинтеза (рис. 16). Как понятно из названия, для ее про- хождения не требуется света. В ее процессе с помощью фермента РУБИСКО CO2 присоединяется к органическим веществам и при участии запасенных в световой стадии АТФ и НАДФ·Н восстанавливается в цикле Кальвина. На одной из стадий этого цикла отчуждается синтезированный моносахарид- гексоза. То есть в итоге, за счет запасенной энергии света углекислый газ восстанавливается до углевода. Хлоропласты содержат свою, специфическую ДНК, сходную с ДНК бактерий. Она отличается от ДНК ядра и передается по наследству через пропластиды, которые находятся в цито- плазме яйцеклетки.
отцовского растения (т.е. через клет- ку пыльцы) хлоропластная ДНК не наследуется. Синтезируемые хлоропластами соединения выполняют не только функцию конституционных веществ, но могут откладываться в них про за- пас. Крахмал, возникающий в хлоропластах на свету, называют первичным (ассимиляционным). Биологическое значение образования первичного крахмала: • в ходе полимеризации глюкозы и образования крахмала уст- раняется один из конечных продуктов фотосинтеза и тем са- мым предупреждается торможение этого процесса; • образование крахмала предотвращает повышение осмоти- ческого давления внутри хлоропласта и возможность его раз- рыва от избытка воды. 3.2. Лейкопласты Общая характеристика. Лейкопласты (от греч. leucós – белый и plastós – вылепленный, оформленный) мелкие бесцветные пластиды. В све- товой микроскоп обнаружить их можно, если внутри них накапливаются крупные включения. Лейкопласты встречаются в клетках органов, скрытых от света: в корнях, корневищах, клубнях, луковицах, семенах, сердцевине стеблей. Ред- ко они находятся в клетках ярко освещенных частей растения. Например, в 36 клетках эпидермиса. Нередко лейкопласты собираются вокруг ядра, окружая его со всех сторон. Форма лейкопластов очень непостоянна и может быстро изменяться даже в одной клетке. Строение лейкопластов. Оболочка лейкопласта состоит из двух эле- ментарных мембран. Внутренняя из них, врастая в строму, образует немно- гочисленные тилакоиды.
обще лейкопласты отличаются слабым развити- ем внутренней мембранной системы. В них отсутствуют граны, а имеются только одиночные тилакоиды. Причем эти тилакоиды располагаются без определенной ориентации или параллельно оболочке пластиды (рис. 14). В лейкопластах имеются ДНК, рибосомы и ферменты, осуществляющие син- тез и гидролиз запасных веществ. Функция лейкопластов – синтез запасных питательных веществ. В первую очередь крахмала, иногда белков, редко – жиров. Лейкопласты, на- капливающие крахмал, называют амилопластами (рис. 14), масла – элайо- пластами (олеопластами), белки – протеинопластами (протеопластами). Крахмал образуется из сахаров, поступающих из фотосинтезирующих кле- ток. Образующийся в лейкопластах крахмал называется вторичным. Он имеет вид зерен различного размера и формы (см. ниже). Запасной белок в лейкопластах может откладываться в виде кристал- лоподобных структур или аморфных включений; липиды – в виде пластог- лобул. Однако белки и липиды встречаются в лейкопластах редко. В одном и том же лейкопласте могут накапливаться разные вещества. 3.3. Хромопласты Общая характеристика. Хромопласты (от хромо- и греч. plastós – вылепленный, оформленный) - пластиды желтого, оранжевого и красного цвета. Хромопласты встречаются в клетках осенних листьев, зрелых пло- дов, лепестков многих растений (лютик - Ranunculus, одуванчик - Taraxacum, нарцисс - Narcissus, тюльпан - Tulipa и др.), редко – в клетках корнеплодов (морковь – Daucus carota, сахарная свекла – Beta vulgaris). Строение хромопластов. Внутренняя мембранная система в них обычно отсутствует (рис. 14). Лишь иногда она представлена оди- ночными тилакоидами. По размерам хромо- пласты меньше хлоропластов. Их форма может быть самой разной (зубчатой, серповидной, игловидной, пластинчатой, в виде треугольни- Рис. 16. Схема основных ков, ромбов), но не линзовидной. функций хлоропласта: Форма хромопластов определяется их 1 - световые реакции локализованы в тилакоидах, 2 происхождением, состоянием в них пигмен- – темновые – в строме тов, систематическим положением образую- 37 щего их растения. Пигменты. Окраска хромопластов обусловлена пигментами группы каротиноидов. Пигменты нерастворимы в воде, но растворяются в жирах. Хромопласты лишены хлорофилла и не способны к фотосинтезу. В зависимости от формы накопления каротиноидов различают хромо- пласты трех типов: глобулярного – каротиноиды растворены в субмикроскопических ли- поидных глобулах (пример – лепестки лютика, алоэ); фибриллярного – каротиноиды собраны в пучки, состоящие из субмик- роскопических нитей и связанные с фибриллами белка (пример – плоды то- матов, мандарина, красного перца); кристаллического – пигменты откладываются в виде мелких, но ви- димых в световой микроскоп кристаллоидов (пример – плоды шиповника, арбузов, лепестки нарциссов, корнеплоды моркови). Наиболее распространенный тип пластид – глобулярный. В них ли- пидные глобулы с растворенными пигментами занимают основной объем пластиды. Встречаются переходные формы от хлоропластов к глобулярному типу хромопластов – хлорохромопласты. У них сохраняется небольшое число мелких гран, межгранные тилакоиды и одновременно имеется большое чис- ло крупных глобул. Хромопласты – конечный этап в развитии пластид. 3.4. Онтогенез и взаимопревращения пластид В ходе эволюции растений исходным типом пластид были хлоропла- сты. При расчленении тела растений на органы из хлоропластов возникли лейко- и хромопласты. Хлоропласты в клетках развиваются из пропластид (т.е. зачаточных пластид). Это мелкие бесцветные или бледно-зеленые недифференцирован- ные пластиды. Они находятся в делящихся клетках корней и побегов. Изучение строения пропластид показало, что они представляют собой лейкопласты мелкого размера. Пропластиды гораздо мельче хлоропластов, не обладают свойственной хлоропластам тилакоидной или ламеллярной (слоистой) структурой. Вместо этого они содержат проламеллярное тело – упорядоченный, «паракристаллический» центр каналов. Каналы после стимуляции светом изменяют свою ориентацию и превращаются в парал- лельные слои. Причем у цветковых растений зрелые хлоропласты могут раз- виться из пропластид только при освещении, а у некоторых голосеменных это превращение может совершаться и в полной темноте. Пропластиды способны реплицироваться путем деления надвое. Именно так увеличивается их число в клетке. Зрелые пластиды также иногда проявляют способность к репликации. В этом отношении они напоминают бактерии. 38 Для пластид характерны относительно легкие переходы от одного типа к другому (рис. 17). Наиболее обычные процессы – превращения лейкопла- стов в хлоропласты и хлоропластов в хромопласты. При этом в лейкопластах формируется характерная для зеленых пластид внутренняя мембранная сис- тема (система тилакоидов). Например, такие превращения происходят при развитии зародыша из зиготы, при развитии листьев в почке. Характерный пример превращения зеленых пластид в хромопласты – при созревании плодов, осеннем окрашивании листьев. При этом пластиды уменьшаются в размерах, происходит постепенное разрушение внутренних мембран и накопление веществ в глобулах. Хлорофилл полностью разруша- ется и перестает маскировать каротиноиды. Преобладающим компонентом таких пластид становятся пластоглобулы. Этот процесс до определенной стадии обратим. Но в природных условиях превращения хромопластов в хлоропласты не происходит. Хромопласты можно рассматривать как заклю- чительный этап развития пластид – этап старения. Лейкопласты тоже могут превращаться в хромопласты. Например, в выделительных клетках при их старении. Превращение хлоропластов в лейкопласты может происходить, если растение поместить в темноту или поранить. В этом случае мембранная сис- Рис. 17. Цикл развития пластид. Последовательные стадии развития пластид: Незеленые пропластидные стадии 1-3: эопласт («эо» – ранний), амилопласт и амебоидная пластида. Стадия 4 – догранальная пластида (может быть зеленой или незеленой). В отсутствие света догранальная стадия может быть представлена этиопластом – стадия 4’. Хромопласты (стадия 1‘) могут возникнуть из пластид нескольких типов 39 тема пластид в значительной степени разрушается, хлорофилл исчезает, но накопления глобул не происходит. Причем этот процесс обратим. Так, если растение вновь поместить на свет, из лейкопластов опять развиваются хло- ропласты. Все взаимопревращения пластид можно отразить в виде схемы: 3.5. Происхождение пластид и митохондрий В настоящее время широкое распространение получила симбиотиче- ская теория происхождения пластид и митохондрий. Согласно этой теории, в процессе эволюции предшественниками пластид и митохондрий были сине- зеленые водоросли и бактерии. Они оказались встроенными в клетку и по- степенно превратились в хлоропласты и митохондрии (рис. 18). Сначала они были окружены плазмалеммой, непрерывной с плазмалеммой зачаточной клетки. Т.е. находились по сути вне протопласта, в его углублениях. Затем окружающие их участки плазмалеммы отделились от остальной плазмати- ческой мембраны и превратились в наружную мембрану оболочки органелл. В результате симбионты превратились в полуавтономные органеллы клетки (рис. 18). Факты, свидетельствующие в пользу симбиотической теории: 1) наличие у пластид и митохондрий двумембранной оболоч- ки; 2) их относительная генетическая автономность (наличие собственной ДНК и системы синтеза белка, сходных по свой- ствам с бактериальными); 3) сходство ультраструктуры клеток сине-зеленых водорослей и хлоропластов высших растений; 4) сходство ультраструктуры клеток бактерий и митохонд- рий эукариотических клеток. 5) сходство процессов деления прокариотических клеток, пла- стид и митохондрий 6) наличие у современных водорослей внутриклеточных обли- гатных и полуоблигатных бактериальных симбионтов. В подтверждение симбиотической гипотезы приводится и такой факт. Если выращивать Euglena на протяжении нескольких поколений при высо- 40
Источник: window.edu.ru
Строение и функции хлоропластов
Хлоропласты — зеленые пластиды, содержащие зеленый пигмент — хлорофилл.
Основная функция хлоропласт — фотосинтез.
В хлоропластах есть свои рибосомы, ДНК, РНК, включения жира, зерна крахмала. Снаружи хлоропласта покрыты двумя белково-липидными мембранами, а в их полужидкую строму (основное вещество) погружены мелкие тельца — граны и мембранные каналы.
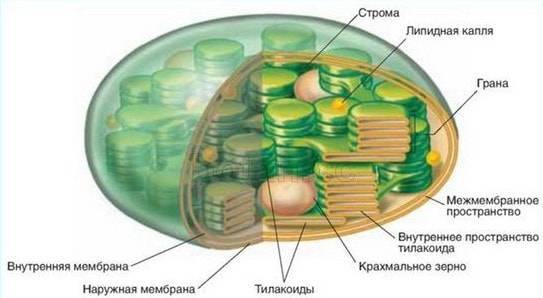
Граны (размером около 1мкм) — пакеты круглых плоских мешочков (тилакоидов), сложенных подобно столбику монет. Располагаются они перпендикулярно поверхности хлоропласта. Тилакоиды соседних гран соединены между собой мембранными каналами, образуя единую систему. Число гран в хлоропластах различно. Например, в клетках шпината каждый хлоропласт содержит 40-60 гран.
Хлоропласты внутри клетки могут двигаться пассивно, увлекаемые током цитоплазмы, либо активно перемещаться с места на место.
- Если свет очень интенсивен, они поворачиваются ребром к ярким лучам солнца и выстраиваются вдоль стенок, параллельных свету.
- При слабом освещении, хлоропласты перемещаются на стенки клетки, обращенные к свету, и поворачиваются к нему своей большой поверхностью.
- При средней освещенности они занимают среднее положение.
Этим достигаются наиболее благоприятные для процесса фотосинтеза условия освещения.
Хлорофилл
В гранах пластид растительной клетки содержится хлорофилл, упакованный с белковыми и фосфолипидными молекулами так, чтобы обеспечить способность улавливать световую энергию.
Молекула хлорофилла очень сходна с молекулой гемоглобина и отличается главным образом тем, что расположенный в центре молекулы гемоглобина атом железа заменен в хлорофилле на атом магния.
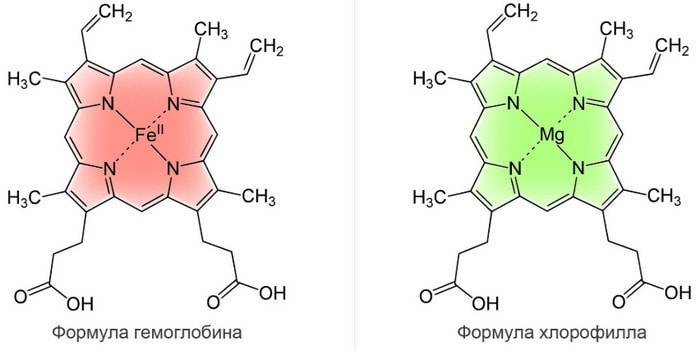
В природе встречается четыре типа хлорофилла: a, b, c, d.
Хлорофиллы a и b содержат высшие растения и зеленые водоросли, диатомовые водоросли содержат a и c, красные — a и d.
Лучше других изучены хлорофиллы a и b (их впервые разделил русский ученый М.С.Цвет в начале XXв.). Кроме них существуют четыре вида бактериохлорофиллов — зеленых пигментов пурпурных и зеленых бактерий: a, b, c, d.
Большинство фотосинтезирующих бактерий содержат бактериохлорофилл a, некоторые — бактериохлорофилл b, зеленые бактерии — c и d.
Хлорофилл обладает способностью очень эффективно поглощать солнечную энергию и передавать ее другим молекулам, что является его главной функцией. Благодаря этой способности хлорофилл — единственная структура на Земле, которая обеспечивает процесс фотосинтеза.
Главная функция хлорофилла в растениях — поглощение энергии света и передача ее другим клеткам.
Пластидам, так же, как и митохондриям, свойственна до некоторой степени автономность внутри клетки. Они размножаются путем деления.
Наряду с фотосинтезом, в пластидах происходит процесс биосинтеза белка. Благодаря содержанию ДНК пластиды играют определенную роль в передаче признаков по наследству (цитоплазматическая наследственность).
Строение и функции хромопластов
Хромопласты относятся к одному из трех видов пластид высших растений. Это небольших размеров, внутриклеточные органеллы.
Хромопласты имеют различный окрас: желтый, красный, коричневый. Они придают характерный цвет созревшим плодам, цветкам, осенней листве. Это необходимо для привлечения насекомых-опылителей и животных, которые питаются плодами и разносят семена на дальние расстояния.

Структура хромопласта похожа на другие пластиды. Их двух оболочек внутренняя развита слабо, иногда вовсе отсутствует. В ограниченном пространстве расположена белковая строма, ДНК и пигментные вещества (каротиноиды).
Каротиноиды – это жирорастворимые пигменты, которые накапливаются в виде кристаллов.
Форма хромопластов очень разнообразна: овальная, многоугольная, игольчатая, серповидная.
Роль хромопластов в жизни растительной клетки до конца не выяснена. Исследователи предполагают, что пигментные вещества играют важную роль в окислительно-восстановительных процессах, необходимы для размножения и физиологичного развития клетки.
Строение и функции лейкопластов
Лейкопласты — это органоиды клетки, в которых накапливаются питательные вещества. Органеллы имеют две оболочки: гладкую наружную и внутреннюю с несколькими выступами.
Лейкопласты на свету превращаются в хлоропласты (к примеру зеленые клубни картофеля), в обычном состоянии они бесцветны.
Форма лейкопластов шаровидная, правильная. Они находятся в запасающей ткани растений, которая заполняет мягкие части: сердцевину стебля, корня, луковиц, листьев.
Функции лейкопластов зависят от их вида (в зависимости от накапливаемого питательного вещества).
Разновидности лейкопластов:
- Амилопласты накапливают крахмал, встречаются во всех растениях, так как углеводы основной продукт питания растительной клетки. Некоторые лейкопласты полностью наполнены крахмалом, их называют крахмальными зернами.
- Элайопласты продуцируют и запасают жиры.
- Протеинопласты содержат белковые вещества.
Лейкопласты также служат ферментной субстанцией. Под действием ферментов быстрее протекают химические реакции. А в неблагоприятный жизненный период, когда процессы фотосинтеза не осуществляются, они расщепляют полисахариды до простых углеводов, которые необходимы растениям для выживания.
В лейкопластах не может происходить фотосинтез, потому что они не содержат гран и пигментов.
Луковицы растений, в которых содержится много лейкопластов, могут переносить длительные периоды засухи, низкую температуру, жару. Это связано с большими запасами воды и питательных веществ в органеллах.
Предшественниками всех пластид является пропластиды, небольшие органоиды. Допускают, что лейко — и хлоропласты способны трансформироваться в другие виды. В конечном итоге после выполнения своих функций хлоропласты и лейкопласты становятся хромопластами — это последняя стадия развития пластид.
Важно знать! Одновременно в клетке растения может находиться только один вид пластид.
Источник: animals-world.ru
Хлоропласт: структура
Хлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений. Охранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
- Мембрана — содержит внутренние и внешние липидные двухслойные оболочки, которые выступают в качестве защитных покрытий и сохраняют замкнутые структуры хлоропластов. Внутренняя мембрана отделяет строму от межмембранного пространства и регулирует прохождение молекул в/из хлоропласта.
- Межмембранное пространство — пространство между внешней и внутренней мембранами.
- Тилакоидная система — внутренняя система мембран, состоящая из сплющенных мешкообразных мембранных структур, называемых тилакоидами, которые служат местами преобразования энергии света в химическую энергию.
- Тилакоид с просветом (люменом) — отсек в каждом тилакоиде.
- Грана — плотные слоистые стопки тилакоидных мешков (10-20), которые служат местами преобразования энергии света в химическую энергию.
- Строма — плотная жидкость внутри хлоропласта, содержащая внутри оболочки, но вне тилакоидной мембраны. Здесь происходит конверсия углекислого газа в углеводы (сахара).
- Хлорофилл — зеленый фотосинтетический пигмент в хлоропласт-гране, поглощающий световую энергию.
Хлоропласт: фотосинтез
» data-layzr=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg» alt=»» width=»500″ height=»365″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg 300w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-500×365.jpg 500w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет.jpg 650w» sizes=»(max-width: 500px) 100vw, 500px» />
При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Понравилась статья? Поделись с друзьями:
Источник: natworld.info