Плазматическая мембрана построена из
Плазматическая мембрана, или плазмолемма, среди клеточных мембран занимает особое место. Это поверхностная периферическая структура, ограничивающая клетку снаружи, что определяет ее непосредственную связь с внеклеточной средой, поэтому она является барьером между внутриклеточным содержимым и внешней средой.
Плазматическая мембрана осуществляет функции, связанные с регулируемым избирательным трансмембранным транспортом веществ и играет роль первичного клеточного анализатора. В этом отношении её можно считать клеточным органоидом, входящим в вакуолярную систему клетки.
Окружая клетку со всех сторон, плазматическая мембрана исполняет роль механического барьера. Механическая устойчивость плазматической мембраны определяется такими дополнительными образованиями как гликокаликс и кортикальный слой цитоплазмы (рис. 127).
Гликокаликс – это внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков — гликопротеидов. В состав гликопротеидов входят такие углеводы как манноза, глюкоза, N-ацетилглюкозамин, сиаловая кислота и др.
Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что снижает в слое скорость диффузии различных веществ. В гликокаликсе находятся выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
В электронном микроскопе гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3-4 нм, покрывающего всю поверхность клетки. Гликокаликс обнаружен практически у всех животных клеток, но особенно хорошо он выражен в щеточной каемке всасывающего эпителия кишечника.
Кроме гликокаликса механическую устойчивость плазматической мембране обеспечивают кортикальный слой цитоплазмы и внутриклеточне фибриллярные структуры. Кортикальный (от слова — cortex -кора, кожица) слой цитоплазмы, лежащий в тесном контакте с наружной мембраной, имеет ряд особенностей. В нем в толщине 0,1-0,5 мкм отсутствуют рибосомы и мембранные пузырьки и в большом количестве сосредоточены микрофиламенты и микротрубочки. Основным компонентом кортикального слоя является сеть актиновых микрофибрилл. Здесь же располагается ряд вспомогательных белков, необходимых для движения участков цитоплазмы.
У простейших, особенно у инфузорий, плазматическая мембрана принимает участие в образовании пелликулы, жесткого слоя, определяющего форму клетки.
Выполнение барьерной роли плазмолеммой заключается также в ограничении свободной диффузии веществ. Она проницаема для воды, газов, малых неполярных молекул жирорастворимых веществ, но совершенно не проницаема для заряженных молекул (ионы) и для крупных незаряженных (сахара) (рис. 130).
Естественные мембраны ограничивают скорость проникновения низкомолекулярных соединений в клетку.
Трансмембранныый перенос ионов и низкомоекулярных соединений.Плазматическая мембрана, как и другие липопротеидные мембраны клетки, является полупроницаемой. Чем больше размер молекул, тем меньше скорость прохождения их через мембрану. В этом отношении она является осмотическим барьером. Максимальной проникающей способностью обладает вода и растворенные в ней газы, медленнее проникают сквозь мембрану ионы (примерно в 104 раз медленнее). Если клетку поместить в среду с концентрацией солей ниже, чем в клетке (гипотония), то вода снаружи устремляется внутрь клетки, что приводит к увеличению объема клетки и разрыву плазматической мембраны. И наоборот, при помещении клетки в растворы солей с более высокой концентрацией, чем в клетке, происходит выход воды из клетки во внешнюю среду. Клетка при этом сморщивается и уменьшается в объеме.
Такой пассивный транспорт воды из клетки и в клетку все же идет с низкой скоростью. Скорость проникновения воды через мембрану составляет около 10-4 см/с, что в 100 000 раз меньше скорости диффузии молекул воды через водный слой толщиной 7,5 нм. Оказывается, для проникновения воды и ионов в клеточной мембране существуют специальные “поры”. Число пор не велико и суммарная их площадь составляет лишь 0,06% всей клеточной поверхности.
Плазматическая мембрана с разной скоростью способна транспортировать ионы и многие мономеры, такие как сахара, аминокислоты и др. Скорость прохождения катионов (K+, Na+) более высока, в сравнении со скоростью прохождения анионов (Cl—).
В транспорт ионов через плазмалемму принимают участие мембранные транспортные белки — пермеазы. Они могут вести транспорт в одном направлении одного вещества (унипорт) или нескольких веществ одновременно (симпорт), или же вместе с импортом одного вещества выводить из клетки другое (антипорт). Например, глюкоза входит в клетки симпортно с ионом Na+.
Транспорт ионов может происходить по градиенту концентрации — пассивно без дополнительной затраты энергии. Так, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. При пассивном транспорте мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану по градиенту концентрации. Часть каналов открыта постоянно, а другая часть закрывается или открывается в ответ на сигнальные молекулы, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки — переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану (облегченная диффузия) (рис. 131).
В организме животных в цитоплазме клеток концентрация ионов резко отличается от плазмы крови, омывающей клетки. Если суммарные концентрации одновалентных катионов как внутри клеток, так и снаружи практически одинаковы (150 мМ), изотоничны,то в цитоплазме концентрация K+ почти в 50 раз выше, а Na+ ниже, чем в плазме крови.
Это связано с тем, что в клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию за счет гидролиза АТФ. Такой тип переноса носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку (рис. 132). С помощью мембранных насосов и с затратой АТФ происходит также регуляция в клетке концентрации двухвалентных катионов Mg2+ и Ca2+
Работа пермеаз и насосов создает в клетке постоянство концентраций осмотических активных веществ или гомеостаз. Примерно 80% всей АТФ клетки тратится на поддержание гомеостаза.
Совместно с активным транспортом ионов через плазматическую мембрану происходит транспорт различных сахаров, нуклеотидов и аминокислот.
Активный транспорт сахаров и аминокислот в бактериальных клетках связан с градиентом ионов водорода.
Участие специальных мембранных белков в пассивном или активном транспорте низкомолекулярных соединений свидетельствует о выcокой специфичности данных процессов, они меняют при этом свою конформацию и функционируют. Таким образом мембраны выступают как анализаторы, как рецепторы.
Везикулярный перенос: эндоцитоз и экзоцитоз. Любые клеточные мембраны не способны к трансмембранному переносу макромолекул, биополимеров, за исключением мембран, имеющих особые белковые комплексные переносчики — порины (мембраны митохондрий, пластид, пероксисом). В клетку макромолекулы попадают заключенными внутри вакуолей или везикул. Везикулярный перенос разделяют на два вида: экзоцитоз — вынос из клетки макромолекулярных продуктов, и эндоцитоз — поглощение клеткой макромолекул (рис. 133).
При эндоцитозе участок плазмалеммы обволакивает внеклеточный материал и заключает его в мембранную вакуоль, возникшую за счет впячивания плазматической мембраны. В такую первичную вакуоль, или эндосому, могут попадать биополимеры, макромолекулярные комплексы, части клеток или даже целые клетки, в которых они распадаются до мономеров и затем путем трансмембранного переноса попадают в гиалоплазму. Биологическое значение эндоцитоза заключается в получении питательных веществ за счет внутриклеточного переваривания, которое осуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой, содержащей набор гидролитических ферментов (см. ниже).
Эндоцитоз формально разделяют на пиноцитоз и фагоцитоз (рис. 134). Фагоцитоз — захват и поглощение клеткой крупных частиц — был впервые описан И,И, Мечниковым. Фагоцитоз встречается как среди одноклеточных (например, амебы, некоторые хищные инфузории), так и у специализированных клеток многоклеточных животных. Сейчас известно, что фагоцитоз и пиноцитоз протекают очень сходно и различия заключаются лишь в массе поглощенных веществ.
В настоящее время эндоцитоз подразделяют на неспецифический или конститутивный, постоянный и специфический, опосредованный рецепторами (рецепторный). Неспецифический эндоцитоз (пиноцитоз и фагоцитоз) протекает автоматически и приводит к захвату и поглощению совершенно чуждых или безразличных для клетки веществ, например, частичек сажи или красителей.
Неспецифический эндоцитоз сопровождается первоначальной сорбцией захватывающего материала гликокаликсом плазмолеммы. Жидкофазный пиноцитоз приводит к поглощению вместе с жидкой средой растворимых молекул, которые не связываются с плазмолеммой.
На следующем этапе происходят впячивания плазматической мембраны, инвагинации, появляются на поверхности клетки выросты, складки, которые как бы захлестываются, складываются, отделяя небольшие объемы жидкой среды (рис. 135, 136). Первый тип возникновения пиноцитозного пузырька, пиносомы, характерен для клеток кишечного эпителия, эндотелия, для амеб, второй — для фагоцитов и фибробластов. Эти процессы зависят от поступления энергии.
Вслед перестройкой поверхности следуют слипание и слияние контактирующих мембран, которые приводят к образованию пеноцитозного пузырька (пиносома). Она отрывается от клеточной поверхности и уходит вглубь цитоплазмы.
Неспецифический и рецепторный эндоцитоз, приводящие к отщеплению мембранных пузырьков, происходят в окаймленных ямках, специализированных участках плазматической мембраны. В окаймленных ямках со стороны цитоплазмы плазматическая мембрана покрыта тонким (около 20 нм) волокнистым слоем, который на ультратонких срезах как бы окаймляет, покрывает небольшие впячивания, ямки (рис.
7). Эти ямки есть почти у всех клеток животных, они занимают около 2% клеточной поверхности. Окаймляющий слой состоит в основном из белка клатрина, ассоциированного с рядом дополнительных белков. Три молекулы клатрина вместе с тремя молекулами низкомолекулярного белка образуют структуру трискелиона, напоминающего трехлучевую свастику (рис. 138). Клатриновый трискелионы на внутренней поверхности ямок плазматической мембраны образуют рыхлую сеть, состоящую из пяти- и шестиугольников, в целом напоминающую корзинку. Клатриновый слой одевает весь периметр отделяющихся первичных эндоцитозных вакуолей, окаймленных пузырьков.
Клатрин относится к одному из видов т.н. “одевающих” белков (COP — coated proteins). Эти белки связываются с интегральными белками-рецепторами со стороны цитоплазмы и образуют одевающий слой по периметру возникающей пиносомы, первичного эндосомного пузырька — “окаймленного” пузырька. в отделении первичной эндосомы участвуют также белки — динамины, которые полимеризуются вокруг шейки отделяющегося пузырька (рис. 139).
После того как окаймленный пузырек отделится от плазмолеммы и начнет переноситься вглубь цитоплазмы клатриновый слой распадается. После потери клатринового слоя эндосомы начинают сливаться друг с другом.
Интенсивность жидкофазного неспецифического пиноцитоза может быть очень высокой. Так клетка эпителия тонкого кишечника образует до 1000 пиносом в секунду, а макрофаги образуют около 125 пиносом в минуту. Размер пиносом невелик, их нижний предел составляет 60-130 нм, но обилие их приводит к тому, что при эндоцитозе плазмолемма быстро замещается, как бы расходуется на образование множества мелких вакуолей. Так у макрофагов вся плазматическая мембрана заменяется за 30 минут, у фибробластов — за два часа.
Дальнейшая судьба эндосом может быть различной, часть из них может возвращаться к поверхности клетки и сливаться с ней, но большая часть вступает в процесс внутриклеточного пищеварения.
В ходе фагоцитоза и пиноцитоза клетки теряют большую площадь плазмолеммы (см. макрофаги), которая довольно быстро восстанавливается при рециклизации мембран, за счет возвращения вакуолей и их встраивания в плазмолемму. Это происходит вследствие того, что от эндосом или вакуолей, так же как и от лизосом могут отделяться небольшие пузырьки, которые вновь сливаются с плазмолеммой.
Специфический или опосредуемый рецепторами эндоцитоз отличается от неспецифического тем, что поглощаются молекулы, для которых на плазматической мембране имеются специфические рецепторы, ассоциирующиеся только с данным типом молекул. Такие молекулы, связывающиеся с белками-рецепторами на поверхности клеток, называют лигандами.
Примером избирательного эндоцитоза является транспорт в клетку холестерина. Этот липид синтезируется в печени и в комплексе с другими фосфолипидами и белковой молекулой образует т.н.
попротеид низкой плотности (ЛНП), который секретируется клетками печени и кровеносной системой разносится по всему телу (рис. 140). Специальные рецепторы плазматической мембраны, диффузно расположенные на поверхности различных клеток, узнают белковый компонент ЛНП, и образуют специфический комплекс рецептор-лиганд. Затем комплекс перемещается в зону окаймленных ямок, окружается мембраной и погружается вглубь цитоплазмы. В ней поглощенные частицы ЛНП подвергается распаду в составе вторичной лизосомы.
Эндосомы характеризуются более низким значением рН (рН 4-5), более кислой средой, чем другие клеточные вакуоли. Это связано с наличием в их мембранах белков протонного насоса, закачивающих ионы водорода с одновременной затратой АТФ (Н+-зависимая АТФаза). Кислая среда внутри эндосом играет решающую роль в диссоциации рецепторов и лигандов. Кроме того, кислая среда является оптимальной для активации гидролитических ферментов в составе лизосом, которые активируются при слиянии лизосом с эндосомами и приводят к образованию эндолизосомы, в которой и происходит расщепление поглощенных биополимерв.
В некоторых случаях не всегда судьба диссоциированных лигандов связана с лизосомным гидролизом. Так в некоторых клетках после связывания рецепторов плазмолеммы с определенными белками, покрытые клатрином вакуоли погружаются в цитоплазму и переносятся к другой области клетки, где сливаются снова с плазматической мембраной, а связанные белки диссоциируют от рецепторов. Так осуществляется перенос, трансцитозис, некоторых белков через стенку эндотелиальной клетки из плазмы крови во межклеточную среду (рис. 141). Другой пример трансцитоза — перенос антител. Так у млекопитающих антитела матери, могут передаваться детенышу через молоко. В этом случае комплекс рецептор-антитело остается в эндосоме без изменений.
Фагоцитоз Фагоцитоз является вариантом эндоцитоза и связан с поглощением клеткой крупных агрегатов макромолекул вплоть до живых или мертвых клеток. Так же как и пиноцитоз, фагоцитоз может быть неспецифическим и специфическим, опосредуемым рецепторами на поверхности плазматической мембраны фагоцитирующих клеток. При фагоцитозе происходит образование больших эндоцитозных вакуолей — фагосом, которые сливаясь с лизосомами образуют фаголизосомы.
На поверхности клеток, способных к фагоцитозу (у млекопитающих это нейтрофилы и макрофаги) существует набор рецепторов, взаимодействующих с белками-лигандами. Так при бактериальных инфекциях антитела к белкам бактерий связываются с поверхностью бактериальных клеток, образуют слой, который узнается специфическими рецепторами на поверхности макрофагов и нейтрофилов, и в местах их связывания начинается поглощение бактерии путем обволакивания ее плазматической мембраной клетки (рис. 142).
Экзоцитоз. Плазматическая мембрана принимает участие в выведении веществ из клетки с помощью экзоцитоза — процесса, обратного эндоцитозу (см. рис. 133).
В случае экзоцитоза, внутриклеточные вакуоли или пузырьки подходят к плазматической мембране. В местах контактов плазматическая и вакуолярная мембраны сливаются, и пузырек опустошается в окружающую среду.
С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Экзоцитоз или секреция в большинстве случаев происходит в ответ на внешний сигнал (нервный импульс, гормоны, медиаторы и др.). В ряде случаев экзоцитоз происходит постоянно (секреция фибронектина и коллагена фибробластами). Сходным образом из цитоплазмы растительных клеток выводятся некоторые полисахариды (гемицеллюлозы), участвующие в образовании клеточных стенок.
Но большинство секретируемых веществ используется другими клетками многоклеточных организмов (секреция молока, пищеварительных соков, гормонов и др.). Часть секретирующих веществ клетки используют для собственных нужд. Например, рост плазматической мембраны осуществляется за счет встраивания участков мембраны в составе экзоцитозных вакуолей, отдельные элементы гликокаликса выделяются клеткой в виде гликопротеидных молекул и т.д.
Выделенные из клеток путем экзоцитоза гидролитические ферменты могут сорбироваться в слое гликокаликса и обеспечивать примембранное внеклеточное расщепление различных биополимеров и органических молекул. Огромное значение примембранное неклеточное пищеварение имеет для животных. Было обнаружено, что в кишечном эпителии млекопитающих в зоне так называемой щеточной каемки всасывающего эпителия, особенно богатой гликокаликсом, обнаруживается огромное количество разнообразных ферментов. Часть этих же ферментов имеет панкреатическое происхождение (амилаза, липазы, различные протеиназы и др.), а часть выделяется собственно клетками эпителия (экзогидролазы, расщепляющие преимущественно олигомеры и димеры с образованием транспортируемых продуктов).
Рецепторная роль плазмалеммы. В качестве рецепторов на поверхности клетки выступают белки мембраны или элементы гликокаликса — гликопротеиды. Чувствительные участки к отдельным веществам могут быть разбросаны по поверхности клетки или собраны в небольшие зоны.
Клетки животных организмов обладают разными наборами рецепторов или же разной чувствительностью одного и того же рецептора.
Многие клеточные рецепторы способны передавать межклеточные сигналы с поверхности внутрь клетки. В настоящее время хорошо изучена система передачи сигнала клеткам с помощью некоторых гормонов, в состав которых входят пептидные цепочки. Они связываются со специфическими рецепторами на поверхности плазматической мембраны клетки. Рецепторы, после связи с гормоном активируют другой белок, лежащий уже в цитоплазматической части плазматической мембраны, — аденилатциклазу. Этот фермент синтезирует молекулу циклического АМФ из АТФ. Циклического АМФ (цАМФ) является вторичным мессенджером — активатором ферментов — киназ, вызывающих модификации других белков-ферментов. Так, при действии на печеночную клетку гормона поджелудочной железы глюкагона, вырабатываемого А-клетками островков Лангерганса стимулируется активация аденилатциклазы. Синтезированный цАМФ активирует протеинкиназу А, которая активирует каскад ферментов, в конечном счете расщепляющих гликоген (запасной полисахарид животных) до глюкозы. Действие инсулина заключается в обратном — он стимулирует вхождение глюкозы в печеночные клетки и отложение ее в виде гликогена.
Эффективность этой аденилатциклазной системы очень высока. Взаимодействие одной или нескольких молекул гормона стимулирует синтез множества молекул цАМФ, что приводит к усилению сигнала в тысячи раз. В данном случае аденилатциклазная система служит преобразователем внешних сигналов.
Другим примером рецепторной активности являются рецепторы ацетилхолина. Ацетилхолин, освобождаясь из нервного окончания, связывается с рецептором на мышечном волокне, вызывает импульсное поступление Na+ в клетку (деполяризация мембраны), открывает сразу около 2000 ионных каналов в зоне нервно-мышечного окончания.
Разнообразие и специфичность наборов рецепторов на поверхности клеток создаёт сложную систему маркеров, позволяющих отличать свои клетки (той же особи или того же вида) от чужих. Сходные клетки вступают друг с другом во взаимодействия, приводящие к слипанию поверхностей (конъюгация у простейших и бактерий, образование тканевых клеточных комплексов). При этом клетки, отличающиеся набором детерминантных маркеров или не воспринимающие их, либо исключаются из такого взаимодействия, либо у высших животных уничтожаются в результате иммунологических реакций (см. ниже).
В плазматической мембране находятся специфические рецепторы, реагирующие на физические факторы. Так, в плазматической мембране фотосинтетических бактерий и синезеленых водорослей расположены белки-рецепторы (хлорофиллы), взаимодействующими с квантами света. В плазматической мембране светочувствительных клеток животных локализована фоторецепторные белки (родопсин), с помощью которых световой сигнал превращается в химический, а затем в электрический.
Межклеточное узнавание. В многоклеточных организмах клетки остаются в связи друг с другом за счет способности их поверхностей слипаться. Это свойство адгезии (соединения, сцепления) клеток определяется свойствами их поверхности и обеспечивается взаимодействием между гликопротеидами плазматических мембран. При таком межклеточном взаимодействии клеток между плазматическими мембранами всегда остается щель шириной около 20 нм, заполненная гликокаликсом.
Было установлено, что за взаимодействие однородных клеток отвечают трансмембранные гликопротеиды. Непосредственно за соединение, адгезию, клеток отвечают молекулы т.н. CAM-белков (cell adhesion molecules). Некоторые из них связывают клетки друг с другом за счет межмолекулярных взаимодействий, другие образуют специальные межклеточные соединения или контакты.
Когда соседние клетки связываются друг с другом с помощью однородных молекул адгезивных белков взаимодействия называется гомофильным, и когда в адгезии участвуют разного рода CAM на соседних клетках- гетерофильным. Встречается межклеточное связывание через дополнительные линкерные молекулы.
Из CAM-белков выделяют несколько классов. Это кадгерины, молекулы адгезии нервных клеток (иммуноглобулино-подобные N-CAM), селектины, интегрины.
Кадгерины представляют собой интегральные фибриллярные мембранные белки, которые образуют параллельные гомодимеры. Отдельные домены этих белков связаны с ионами Ca2+, что придает им определенную жесткость. Кадгеринов насчитывают более 40 видов. Так Е-кадгерин характерен для клеток преимплантированных эмбрионов и для эпителиальных клеток взрослых организмов. P-кадгерин характерен для клеток трофобласта, плаценты и эпидермиса.
Молекулы адгезии нервных клеток (N-CAM) принадлежат к суперсемейству иммуноглобулинов, они образуют связи между нервными клетками. Некоторые из N-CAM участвуют в соединении синапсов, а также при адгезии клеток иммунной системы.
Селектины также интегральные белки плазматической мембраны участвуют в адгезии эндотелиальных клеток, в связывании кровяных пластинок, лейкоцитов.
Интегрины представляют собой гетеродимеры, с a и b-цепями. Интегрины в первую очередь осуществляют связь клеток с внеклеточными субстратами, но могут участвовать и в адгезии клеток друг с другом.
Узнавание чужеродных белков. На попавшие в организм чужеродные макромолекулы (антигены), развивается иммунная реакция. Суть ее заключается в том, что часть лимфоцитов вырабатывает специальные белки — антитела, которые специфически связываются с антигенами. Так, например, макрофаги своими поверхностными рецепторами узнают комплексы антиген-антитело и поглощают их (например, поглощение бактерий при фагоцитозе).
В организме всех позвоночных также существует система рецепции чужеродных клеток или же своих, но с измененными белками плазматической мембраны, например при вирусных инфекциях или при мутациях, часто связанных с опухолевым перерождением клеток.
На поверхности всех клеток позвоночных располагаются белки, т.н. главного комплекса гистосовместимости (major histocompatibility complex — MHC). Это интегральные белки гликопротеины, гетеродимеры. Каждый индивидуум имеет свой набор таких белков MHC. Это приводит к тому, что каждая клетка данного организма отличается от клеток индивидуума этого же вида. Специальная форма лимфоцитов, Т-лимфоциты, узнают MHC своего организма и малейшие изменения в его структуре (например, связь с вирусом, или результат мутации в отдельных клетках), приводит к тому, что Т-лимфоциты узнают такие изменившиеся клетки и их уничтожают. Но уничтожают не путем фагоцитоза а выделяют из секреторных вакуолей белки-перфорины, которые встраиваются в цитоплазматическую мембрану измененной клетки, образуют в ней трансмембранные каналы, тем самым делают плазматическую мембрану проницаемой, что и приводит к гибели измененной клетки (рис. 143, 144).
Источник: studopedia.ru
Плазматическая мембрана
Плазмалемма — это перегородка, находящаяся внутри, непосредственно за оболочкой. Она делит клетку на определенные отсеки, которые направлены на компартменты или же органеллы. В них содержатся специализированные условия среды. Клеточная стенка полностью закрывает всю клеточную мембрану. Она выглядит как двойной слой молекул.
Основные сведения
Состав плазмалеммы — это фосфолипиды или же, как их еще называют, сложные липиды. Фосфолипиды имеют несколько частей: хвост и головку. Специалисты называют гидрофобные и гидрофильные части: в зависимости от строения животной или растительной клетки. Участки, которые именуются головкой — обращены внутрь клетки, а хвосты — наружу. Плазмалеммы по структуре являются инвариабельными и очень похожи у различных организмов; чаще всего исключение могут составить археи, у которых перегородки состоят из различных спиртов и глицерина.
Толщина плазмалеммы приблизительно 10 нм.
В малом содержании в состав биологической мембраны входят некоторые виды белков. Например, белки которые пронизывают всю мембрану насквозь, их называют интегральными. Мембраны, которые входят в состав и внешнего, и во внутреннего слоя (слой чаще всего бывает липидным), называются полуинтегральными.

Существуют перегородки, которые находятся на внешней стороне или же снаружи части, вплотную прилегающей к мембране — их называют поверхностными. Некоторые виды белка могут быть своеобразными контактными точками для клеточной мембраны и оболочки. Внутри клетки находится цитоскелет и наружная стенка. Определенные виды интегрального белка могут быть использованы как каналы в ионных транспортных рецепторах (параллельно с нервными окончаниями).
Если использовать электронный микроскоп, то можно получить данные, на основе которых можно построить схему строения всех частей клетки, а также основных составляющих и оболочек. Верхний аппарат будет состоять из трех субсистем:
- комплексное надмембранное включение;
- плазматическая мембрана;
- опорно-сократительный аппарат цитоплазмы, который будет иметь субмембранную часть.
К данному аппарату можно отнести цитоскелет клетки. Цитоплазма с органоидами и ядром называется — ядерный аппарат. Цитоплазматическая или, по-другому, плазматическая клеточная мембрана, находится под клеточной оболочкой.
Слово «мембрана» произошло от латинского слова membrum, которое можно перевести как «кожа» или «оболочка». Термин предложили более 200 лет назад и им чаще называли края клетки, но в период, когда началось использование различного электронного оборудования, установили, что плазматические цитолеммы составляют множество различных элементов оболочки.
Элементы чаще всего структурные, такие как:
- митохондрии;
- лизосомы;
- пластиды;
- перегородки.
Одна из первых гипотез относительно молекулярного состава плазмалеммы была выдвинута в 1940 году научным институтом Великобритании. Уже в 1960 году Уильям Робертс предложил миру гипотезу «Об элементарной мембране». Она предполагала, что все плазмалеммы клетки состоят из определенных частей, по сути, являются сформированными по общему принципу для всех царств организмов.
В начале семидесятых годов XX века было открыто множество данных, на основании которых в 1972 году ученые из Австралии предложили новую мозаично-жидкостную модель строения клеток.
Строение плазматической мембраны
Модель 1972-го года является общепризнанной и по сей день. То есть в современной науке, различные ученые, работающие с оболочкой, опираются на теоретический труд «Строение биологической мембраны жидкостно-мозаичной модели».
Молекулы белков связаны с липидным бислоем и пронизывают всю мембрану полностью — интегральные белки (одно из общепринятых названий — это трансмембранные белки).
Оболочка в составе имеет различные углеводные компоненты, которые будут выглядеть как полисахаридная или сахаридная цепь. Цепь, в свою очередь, будет соединена липидами и белком. Соединенные молекулами белка цепи называются гликопротеинами, а молекулами липидов — гликозидами. Углеводы находятся на внешней стороне мембраны и выполняют функции рецепторов в клетках животного происхождения.
Гликопротеин — представляют собой комплекс надмембранных функций. Его еще называют гликокаликс (от греческих слов глик и каликс, что в переводе означает «сладкий» и «чашка»). Комплекс способствует адгезии клеток.
Функции плазматической мембраны
Барьерная
Помогает отделить внутренние составляющие клеточной массы от тех веществ, которые находятся извне. Предохраняет организм от попадания различных веществ, которые будут являться для него чужеродными, и помогает поддерживать внутриклеточный баланс.
Транспортная
Клетка имеет свой «пассивный транспорт» и использует его для уменьшения расхода энергии. Транспортная функция работает в следующих процессах:
- эндоцитоз;
- экзоцитоз;
- натриевый и калиевый обмен.
На внешней стороне мембраны находится рецептор, на участке которого происходит смешивание гормонов и различных регуляторных молекул.
Пассивный транспорт — процесс, при котором вещество проходит через мембрану, при этом энергия не затрачивается. Иными словами, вещество доставляется из области клетки с высокой концентрацией, в ту сторону, где концентрация будет более низкая.
Существует два вида:
- Простая диффузия — присуща маленьким нейтральным молекулам H2O, CO2 и О2 и некоторыми гидрофобным органическим веществам с низкой молекулярной массой и соответственно без проблем проходят через фосфолипиды мембраны. Эти молекулы могут проникать через мембрану вплоть до того времени, пока градиент концентрации будет стабилен и неизменен.
- Облегченная диффузия — характерна для различных молекул гидрофильного типа. Они также могут проходить через мембрану согласно градиенту концентрации. Однако, процесс будет осуществляться с помощью различных белков, которые будут образовывать специфические каналы ионных соединений в мембране.
Активный транспорт — это перемещение различных составляющих через стенку мембраны в противовес градиенту. Такое перенесение требует значительных затрат энергетических ресурсов в клетке. Чаще всего именно активный транспорт является основным источником потребления энергии.
Выделяют несколько разновидностей активного транспорта при участии белков-переносчиков:
- Натриево-калиевый насос. Получение клеткой необходимых минералов и микроэлементов.
- Эндоцитоз — процесс, при котором происходит захват клеткой твердых частиц (фагоцитоз) или же различных капель любой жидкости (пиноцитоз).
- Экзоцитоз — процесс, при котором происходит выделение из клетки определенных частиц во внешнюю окружающую среду. Процесс является противовесом эндоцитоза.
Термин «эндоцитоз» произошел от греческих слов «энда» (изнутри) и «кетоз» (чаша, вместилище). Процесс характеризует захват внешнего состава клеткой и осуществляется при производстве мембранных пузырьков. Этот термин был предложен в 1965 году профессором цитологии из Бельгии Кристианом Бэйлсом, он изучал поглощение различных веществ клетками млекопитающих, а также фагоцитоз и пиноцитоз.
Фагоцитоз
Происходит при захвате клеткой определенных твердых частиц или же живых клеток. А пиноцитоз — это процесс, при котором капли жидкости захватываются клеткой. Фагоцитоз (от греческих слов «пожиратель» и «вместилище») — процесс при котором очень маленькие объекты живой природы захватываются и поглощаются, так же как и твердые части различных одноклеточных организмов.
Открытие процесса принадлежит физиологу из России — Вячеславу Ивановичу Мечникову, который определил непосредственно процесс, при этом он проводил различные испытания с морскими звездами и крошечными дафниями.
В основе питания одноклеточных гетеротрофных организмов лежит их способность переваривать, а также захватывать различные частицы.
Мечников описал алгоритм поглощения бактерии амебой и общий принцип фагоцитоза:
- адгезия — прилипание бактерий к мембране клетки;
- поглощение;
- образование пузырька с бактериальной клеткой;
- откупоривание пузырька.
Исходя из этого, процесс фагоцитоза состоит из таких этапов:
- Поглощаемая частица крепится к мембране.
- Окружение поглощаемой частицы мембраной.
- Образование мембранного пузырька (фагосома).
- Открепление мембранного пузырька (фагосомы) во внутреннюю часть клетки.
- Объединение фагосомы и лизосомы (переваривание), а также внутреннее перемещение частиц.
Можно наблюдать полное или частичное переваривание.
В случае частичного переваривания чаще всего образуется остаточное тельце, которое будет находиться внутри клетки некоторое время. Те остатки, которые будут непереварены, изымаются (эвакуируются) из клетки путем экзоцитоза. В процессе эволюции эта функция предрасположенности к фагоцитозу постепенно отделилась и перешла от различных одноклеточных к специализированным клеткам (таким как пищеварительная у кишечнополостных и губок), а после к особым клеткам у млекопитающих и человека.
К фагоцитозу предрасположены лимфоциты и лейкоциты в крови. Сам процесс фагоцитоза нуждается в больших затратах энергии и напрямую объединен с активностью внешней клеточной мембраны и лизосомы, при которых находятся пищеварительные ферменты.
Пиноцитоз
Пиноцитоз — это захват поверхностью клетки какой-либо жидкости, в которой находятся различные вещества. Открытие явления пиноцитоза принадлежит ученому Фицджеральду Льюису. Произошло это событие в 1932 году.
Пиноцитоз — это один из основных механизмов, при котором в клетку попадают высокомолекулярные соединения, например, различные гликопротеины или же растворимые белки. Пиноцитозная активность, в свою очередь, невозможна без физиологического состояния клетки и зависит от ее состава и состава окружающей среды. Самый активный пиноцитоз мы можем наблюдать у амебы.
У человека пиноцитоз наблюдается в клетках кишечника, в сосудах, почечных канальцах, а также в растущих ооцитах. Для того чтобы изобразить процесс пиноцитоза, которой будет осуществляться с помощью лейкоцитов человека, можно сделать выпячивание плазматической мембраны. При этом части будут отшнуровываться и отделяться. Процесс пиноцитоза нуждается в затрате энергии.
Этапы процесса пиноцитоза:
- На наружной клеточной плазмалемме появляются тонкие наросты, которые окружают капли жидкости.
- Этот участок внешней оболочки становится тоньше.
- Образование мембранного пузырька.
- Стенка прорывается (проваливается).
- Пузырек перемещается в цитоплазме и может слиться с различными пузырьками и органоидами.
Экзоцитоз
Термин произошел от греческих слов «экзо» — наружный, внешний и «цитоз» — сосуд, чаша. Процесс заключается в выделении клеточной частью определенных частиц во внешнюю среду. Процесс экзоцитоза является противоположным пиноцитозу.
В процессе экоцитоза из клетки выходят пузырьки внутриклеточной жидкости и переходят на внешнюю мембрану клетки. Содержимое внутри пузырьков может выделяться наружу, а мембрана клетки сливается с оболочкой пузырьков. Таким образом, большинство макромолекулярных соединений будет происходить именно этим способом.
Экзоцитоз выполняет ряд задач:
- доставка молекул на внешнюю клеточную мембрану;
- транспортировка по всей клетке веществ, которые будут нужны для роста и увеличения площади мембраны, например, определенных белков или же фосфолипидов;
- освобождение или соединение различных частей;
- выведение вредных и токсических продуктов, которые появляются при метаболизме, например, соляной кислоты секретируемой клетками слизистой оболочки желудка;
- транспортировка пепсиногена, а также сигнальных молекул, гормонов или нейромедиаторов.
Специфические функции биологических мембран:
- генерация импульса, происходящего на нервном уровне, внутри мембраны нейрона;
- синтез полипептидов, а также липидов и углеводов шероховатой и гладкой сети эндоплазматической сетки;
- изменение световой энергии и ее преобразование в энергию химическую.
Источник: LivePosts.ru
Мы начнем гистологию с изучения эукариотической клетки, являющейся самой простой системой, наделенной жизнью. При исследовании клетки в световом микроскопе мы получаем информацию о ее размере, форме, и эта информация связана с наличием у клеток ограниченных мембраной границ. С развитием электронной микроскопии (ЭМ) наши представления о мембране, как о четко ограниченной линии раздела между клеткой и окружающей средой изменились, ибо оказалось, что на поверхности клетки имеется сложная структура, состоящая из следующих 3-х компонентов:
1. Надмембранный компонент (гликокаликс) (5 — 100 нм);
2. Плазматическая мембрана (8 — 10 нм);
3. Подмембранный компонент (20 – 40 нм).
При этом 1 и 3 компоненты вариабельны и зависят от типа клеток, наиболее статичным представляется строение плазматической мембраны, которую мы и рассмотрим.
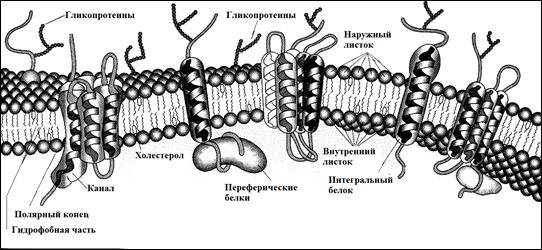
Плазматическая мембрана. Изучение плазмолеммы в условиях ЭМ привело к заключению об однотипности ее структурной организации, при которой она имеет вид триламинарной линии, где внутренний и наружный слои электронноплотные, а расположенный между ними – более широкий слой представляется электроннопрозрачным. Такой тип структурной организации мембраны свидетельствует об ее химической гетерогенности. Не касаясь дискуссии по этому вопросу, оговорим, что плазмолемма состоит из трех типов веществ: липидов, белков и углеводов.
Липиды, входящие в состав мембран, обладают амфифильными свойствами за счет присутствия в их составе как гидрофильных, так и гидрофобных групп. Амфипатический характер липидов мембраны способствует образованию липидного бислоя. При этом в фосфолипидах мембраны выделяют два домена:
а) фосфатная– голова молекулы, химические свойства этого домена определяют его растворимость в воде и его называют гидрофильным;
б) ацильные цепи, представляющие собой этерифицированные жирные кислоты – это гидрофобный домен.
Типы мембранных липидов: основным классом липидов биологических мембран являются фосфолипиды, они формируют каркас биологической мембраны. См. рис.1
Рис. 1: Типы мембранных липидов
Биомембраны – это двойной слой амфифильных липидов(липидный бислой). В водной среде такие амфифильные молекулы самопроизвольно образуют бислой, в котором гидрофобные части молекул ориентированы друг к другу, а гидрофильные к воде. См. рис. 2
Рис. 2: Схема строения биомембраны
В состав мембран входят липиды следующих типов:
1. Фосфолипиды;
2. Сфинголипиды — “головки” + 2 гидрофобных “хвоста”;
3. Гликолипиды.
Холестерин (ХЛ)– находится в мембране в основном в срединной зоне бислоя, он амфифилен и гидрофобный (за исключением одной гидроксигруппы). Липидный состав влияет на свойства мембран: отношение белок/липиды близок 1:1, однако миелиновые оболочки обогащены липидами, а внутренние мембраны – белками.
Способы упаковки амфифильных липидов:
1. Бислои (липидная мембрана);
2. Липосомы — это пузырек с двумя слоями липидов, при этом как внутренняя, так и наружная поверхности являются полярны;
3. Мицеллы – третий вариант организации амфифильных липидов – пузырек, стенка которого образована одним слоем липидов, при этом их гидрофобные концы обращены к центру мицеллы и их внутренняя среда является не водной, агидрофобной.
Наиболее распространенной формой упаковки молекул липидов является образование ими плоского бислоя мембран. Липосомы и мицеллы – это скорые транспортные формы, обеспечивающие перенос веществ в клетку и из нее. В медицине липосомы используют для переноса водорастворимых, а мицеллы – для переноса жирорастворимых веществ.
Белки мембраны
1. Интегральные (включены в липидные слои);
2. Периферические. См. рис. 3
Интегральные (трансмембранные белки):
1. Монотопные– (например, гликофорин. Они пересекают мембрану 1 раз), и являются рецепторами, при этом их наружный – внеклеточный домен – относится к распознающей части молекулы;
2.Политопные– многократно пронизывают мембрану – это тоже рецепторные белки, но они активизируют путь передачи сигнала внутрь клетки;
3.Мембранные белки, связанные с липидами;
4. Мембранные белки, связанные с углеводами.
Рис. 3: Белки мембраны
Периферические белки:
Не погружены в липидный бислой и не соединены с ним ковалентно. Они удерживаются за счет ионных взаимодействий. Периферические белки ассоциированы с интегральными белками в мембране за счет взаимодействия — белок-белковые взаимодействия.
1. Спектрин, который расположен на внутренней поверхности клетки;
2.Фибронектин, локализован на наружной поверхности мембраны.
Белки – обычно составляют до 50% массы мембраны. При этом интегральные белкивыполняют следующие функции:
а) белки ионных каналов;
б) рецепторные белки.
А периферические мембранные белки (фибриллярные, глобулярные) выполняют следующие функции:
а) наружные (рецепторные и адгезионные белки);
б) внутренние – белки цитоскелета (спектрин, анкирин), белки системы вторых посредников.
Ионные каналы– это сформированные интегральными белками каналы, они формируют небольшую пору, через которую по электрохимическому градиенту проходят ионы. Наиболее известные каналы – это каналы для Nа, К, Са, Сl.
Существуют и водные каналы – это аквопорины (эритроциты, почка, глаз).
Надмембранный компонент – гликокаликс, толщина 50 нм. Это углеводные участки гликопротеинов и гликолипидов, обеспечивающие отрицательный заряд. Под ЭМ – это рыхлый слой умеренной плотности, покрывающий наружную поверхность плазмолеммы. В состав гликокаликса помимо углеводных компонентов входят периферические мембранные белки (полуинтегральные). Функциональные участки их находятся в надмембранной зоне — это иммуноглобулины. См. рис. 4
Функция гликокаликса:
1. Играют рольрецепторов;
2. Межклеточное узнавание;
3. Межклеточные взаимодействия (адгезивные взаимодействия);
4. Рецепторы гистосовместимости;
5. Зона адсорбции ферментов (пристеночное пищеварение);
6.Рецепторы гормонов.
Рис. 4: Гликокаликс и подмембранные белки
Подмембранный компонент — самая наружная зона цитоплазмы, обычно обладает относительной жесткостью и эта зона особенно богата филаментами (d = 5-10 нм). Предполагают, что интегральные белки, входящие в состав клеточной мембраны, прямо или косвенно связаны с актиновыми филаментами, лежащими в подмембранной зоне. При этом экспериментально доказано, что при агрегации интегральных белков, находящийся в этой зоне актин и миозин также агрегируют, что указывает на участие актиновых филамент в регуляции формы клетки.
| <== предыдущая лекция | | | следующая лекция ==> |
| РЫХЛАЯ ВОЛОКНИСТАЯ СОЕДИНИТЕЛЬНАЯ ТКАНЬ | | |
Источник: helpiks.org
Плазматическая мембрана клетки. Строение плазматической мембраны. Функции плазматической мембраны.
Животные клетки ограничены плазматической мембраной. На ее строении, очень сходном со строением многих внутриклеточных мембран, мы остановимся несколько подробнее. Основной матрикс мембраны состоит из липидов, главным образом фосфатидил-холина. Эти липиды состоят из головной гидрофильной группы, к которой присоединены длинные гидрофобные углеводородные цепи. В воде такие липиды спонтанно формируют двуслойную пленку толщиной 4-5 нм, в которой гидрофильные группы обращены к водной среде, а гидрофобные углеводородные цепи располагаются в два ряда, образуя безводную липидную фазу. Клеточные мембраны представляют собой липидные бислои именно такого типа и содержат гликолипиды, холестерол и фосфолипиды.
Гидрофильная часть гликолипидов образована олигосахаридами. Гликолипиды всегда располагаются на наружной поверхности плазматической мембраны, причем олигосахаридная часть молекулы ориентирована подобно волоску, погруженному в окружающую среду. Разбросанные среди фосфолипидов в почти равном с ними количестве молекулы холестерола стабилизируют мембрану. Распределение различных липидов во внутреннем и наружном слоях мембраны неодинаково, и даже в пределах одного слоя имеются участки, в которых концентрируются отдельные виды липидов. Такое неравномерное распределение, вероятно, имеет какое-то, пока еще неясное, функциональное значение.
Главными функциональными элементами, погруженными в сравнительно инертный липидныи матрикс мембраны, являются белки. Белок по массе составляет от 25 до 75% в различных мембранах, но, поскольку белковые молекулы намного крупнее, чем липидные, 50% по массе эквивалентны соотношению: 1 молекула белка на 50 молекул липида. Одни белки пронизывают мембрану от ее наружной до внутренней поверхности, другие же закреплены в каком-то одном слое. Белковые молекулы обычно ориентированы так, что их гидрофобные группы погружены в липидную мембрану, а полярные гидрофильные группы на поверхности мембраны погружены в водную фазу. Многие белки наружной поверхности мембраны представляют собой гликопротеины; их гидрофильные сахаридные группы обращены во внеклеточную среду.
Источник: otvet.mail.ru