Эндоплазматическая мембрана
Строение эндоплазматической сети
ЭПС – мембранная органелла, состоящая из плоских мембранных мешочков – цистерн, каналов и трубочек. Благодаря такому строению эндоплазматическая сеть значительно увеличивает площадь внутренней поверхности клетки и делит клетку на секции. Внутри она заполнена матриксом (умеренно плотный рыхлый материал (продукт синтеза)). Содержание различных химических веществ в секциях неодинаково, потому в клетке как одновременно, так и в определённой последовательности могут происходить различные химические реакции в незначительном объёме клетки. Эндоплазматическая сеть открывается в перинуклеарное пространство (полость между двумя мембранами кариолемы).

Мембрана эндоплазматической сети состоит из белков и липидов (в основном фосфолипидов), а так же ферментов: аденозинтрифосфатазы и ферментов синтеза мембранных липидов.
Различают два вида эндоплазматической сети:
- Гладкую (агранулярную, аЭС), представленную трубочками, которые анастамозируют между собой и не имеют на поверхности рибосом;
- Шероховатую (гранулярную, грЭС), состоящую так же из соединённых между собой цистерн, но они покрыты рибосомами.
Гранулярная ЭС свойственна всем клеткам (кроме сперматозоидов), но степень её развития разная и зависит от специализации клетки.
Сильно развита грЭС эпителиальных железистых клеток (поджелудочной железы, вырабатывающих пищеварительные ферменты, печени – синтезирующих альбумины сыворотки крови), фибробластов (клеток соединительной ткани, продуцирующих белок коллаген), плазматических клеток (продуцирование иммуноглобулинов).
Агранулярная ЭС преобладает в клетках надпочечников (синтез стероидных гормонов), в клетках мышц (обмен кальция), в клетках фундальных желез желудка (выделение ионов хлора).
Другим видом мембран ЭПС являются разветвлённые мембранные трубочки, содержащие внутри большое количество специфических ферментов, и везикулы – маленькие, окружённые мембраной пузырьки, в основном находящиеся рядом с трубочками и цистернами. Они обеспечивают перенесение тех веществ, которые синтезируются.
Функции ЭПС
Эндоплазматическая сеть – это аппарат синтеза и, частично, транспорта веществ цитоплазмы, благодаря которому клетка выполняет сложные функции.
Гладкая и шероховатая эндоплазматические сети своими мембранами и содержимым (матриксом) выполняют общие функции:
- разделительную (структурирующую), благодаря чему цитоплазма упорядоченно распределяется и не смешивается, а так же предотвращает попадание в органеллу случайных веществ;
- трансмембранное транспорт, благодаря которому осуществляется перенесение сквозь стенку мембраны необходимых веществ;
- синтез липидов мембраны с участием ферментов, содержащихся в самой мембране и обеспечивающих репродукцию эндоплазматической сети;
- благодаря разнице потенциалов, возникающая между двумя поверхностями мембран ЭС возможно обеспечение проведения импульсов возбуждения.
Кроме того, каждой из разновидностей сети свойственны свои специфические функции.
Функции гладкой (агранулярной) эндоплазматической сети
Агранулярная эндоплазматическая сеть, кроме названных функций, общих для обоих видов ЭС, выполняет ещё и свойственные только для неё функции:
- депо кальция. Во многих клетках (в скелетных мышцах, в сердце, яйцеклетках, нейронах) существуют механизмы, способные изменять концентрацию ионов кальция. Поперечнополосатая мышечная ткань содержит специализированную эндоплазматическую сеть, называемую саркоплазматическим ретикулумом. Это резервуар кальций-ионов, а мембраны этой сети содержат мощные кальциевые помпы, способные выбрасывать в цитоплазму большое количество кальция или транспортировать его в полости каналов сети за сотые доли секунды;
- синтез липидов, веществ типа холестерина и стероидных гормонов. Стероидные гормоны синтезируются в основном в эндокринных клетках половых желез и надпочечников, в клетках почек и печени. Клетки кишечника синтезируют липиды, которые выводятся в лимфу, а потом в кровь;
-
детоксикационная функция – обезвреживание єкзогенных и эндогенных токсинов;
-
ферменты органеллы берут участие в синтезе гликогена (в клетках печени).
Функции шероховатой (гранулярной) эндоплазматической сети
Для гранулярной эндоплазматической сети, кроме перечисленных общих функций, свойственны ещё и специальные:
- синтез белков на грЭС имеет некоторые особенности. Начинается он на свободных полисомах, которые в дальнейшем связываются с мебранами ЭС.
- Гранулярная эндоплазматическая сеть синтезирует: все белки клеточной мембраны (кроме некоторых гидрофобных белков, белков внутренних мембран митохондрий и хлоропластов), специфические белки внутренней фазы мембранных органелл, а так же секреторные белки, которые транспортируются по клетке и поступают во внеклеточное пространство.
- пострансляционная модификация белков: гидроксилирование, сульфатирование, фосфориллирование. Важным процессом является гликозилирование, которое происходит под действием связанного с мембраной фермента гликозилтранферазы. Гликозилирование происходит перед секрецией или транспортом веществ к некоторым участкам клетки ( комплексу Гольджи, лизосомам или плазмолемме).
- транспорт веществ по внутримембранной части сети. Синтезированные белки по промежуткам ЭС перемещаются к комплексу Гольджи, который выводит вещества из клетки.
- благодаря участию гранулярной эндоплазматической сети образуется комплекс Гольджи.
Функции зернистой эндоплазматической сети связаны с транспортом белков, которые синтезируются в рибосомах и расположены на её поверхности. Синтезированные белки поступают внутрь ЭПС, скручиваются и приобретают третичную структуру.
Белок, который транспортируется к цистернам, значительно изменяется на своём пути. Он может, например, фосфорилироваться или превращаться в гликопротеид. Обычный путь для белка – это путь через зернистую ЭПС в аппарат Гольджи, откуда он или выходит наружу клетки, или поступает к другим органеллам той же клетки, например, к лизосомам), или откладывается в виде запасных гранул.
В клетках печени как зернистая, так и незернистая эндоплазматическая сетка берут участие в процессах детоксикации ядовитых веществ, которые потом выводятся из клетки.
Как и внешняя плазматическая мембрана, эндоплазматическая сетка имеет избирательную проницаемость, вследствие чего концентрация веществ внутри и снаружи каналов сетки неодинакова. Это имеет значение для функции клетки.
Образование эндоплазматической сети
Липидные компоненты мембран эндоплазматической сети синтезируются ферментами самой сети, белковый – поступает из рибосом, расположенных на её мембранах. В гладкой (агранулярной) эндоплазматической сети нет собственных факторов синтеза белка, потому считается, что эта органелла образуется в результате потери рибосом гранулярной эндоплазматической сетью.
Источник: spravochnick.ru
Впервые эндоплазматический ретикулум был обнаружен американским учёным К. Портером в 1945 году посредством электронной микроскопии.
Эндоплазматический ретикулум (ЭПР) (лат. reticulum — сеточка) или эндоплазматическая сеть (ЭПС) — внутриклеточный органоид эукариотической клетки, представляющий собой разветвленную систему соединённых между собой каналов и полостей, ограниченных одинарной мембраной, поверхность которой составляет более 50% площади всех клеточных мембран. Мембрана ЭПС тоньше чем плазмалемма и содержит более высокую концентрации. белка. Непосредственным продолжение ЭПС является наружная ядерная мембрана.
На поверхности мембран ЭПС происходит большая часть реакций метаболизма, протекающих в клетке. ЭПС разделяет цитоплазму на отдельные отсеки. по каналам ЭПС происходит упорядоченный обмен веществами и энергией между различными компонентами клетки.
ЭПС – генератор мембран для плазмолеммы, ап гольджи и лизосом.
Гранулярная или шероховатая эпс.
наружная обращеная к цитоплазме, сторона грЭПС покрыта рибосомами (котор имеют вид мелк гранул; поступают из ядра благодаря связи мембраны с наруж мембр ядра).
грЭПС – образ уплощенными мембранными цистернами и трубочками на наружной поверхности которых располог рибосомы и полисомы, придающие мембране зернист вид.
Мембраны содерж белки (которые обеспеч связывание рибосом, уплощение цистерн).
Полость грЭПС сообщ с перенуклеарн пространство. Благодаря грЭПС происход отделение вновь синтезированных белковых молекул от гиалоплазмы.
грЭПС хорошо развита в клетках, специализирующихся на белковом синтезе.
ФУНКЦИИ: 1)биосинтез всех мембранных белков, предназначенных для экспорта из клетки.
2) в грЭПС происход посттрансляционный процессинг белков. (созревание белка). белки приобрет характер для них третичную или четвертичную структуру. потом транспортир в комплекс гольджи — > потом в другие органойды.
3) гЭПС выполняет ф-ю пространственного разделения ферментных систем. резделени клетки с помощью мембран на отдел отсеки – компарменты.
4) обеспеч транспорт синтезируемых веществ в аппарат гольджи.
Гладкая или агранулярная ЭПС.
не имеет рибосом. Сост из сильно ветвящихся канальцев и мелких вакуолей диаметром 20-100 нм. гЭПС — трёхмерная замкнутая сеть мембранных анастамозирующих турбочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутсвт.
На цитоплазмотической поверхности гЭПС синтезируется большая часть липидов клетки, которые вход в состав всех её мембран. Часть синтезир на гЭПС белков и липидов встраивается в неё, но увеличения общей площади мембраны при этом не происход. на гЭПС соверш синтез и распад многих углеводов, включ полисахариды, образ стеройдные гормоны.
В Гэпс накаплив многие ядовит в-ва, подлежащ удален из клетки.
гЭПС наиболее развита в клетках с интенсивным жировыми углеводным обменом.
ФУНКЦИИ: 1) синтез липидов; (на мембранах) 2) синтез гликогена (в клетках печени)
3) синтез холестерина и других стеройдов 4) детоксикация эндогенных и экзогенных в-в. (в клетках печени) 5) накопление ионов Са. гЭПС в Миш клетках играет роль депо ионов кальция, необходимых для мыш сокращ. 6) компартментализация (эпс раздел клетку на отдел отсеки) 7) транспорт синтезируемых веществ 8) в мегакариоцитах элементы гЭПС образуют демаркационные каналы, разделяющие формирующие тромбоциты. 9) восстановление кариолеммы в телофазе митоза.
РИБОСОМЫ
Рибосомы впервые были описаны как уплотненные частицы, или гранулы, клеточным биологом румынского происхождения Джорджем Паладе в середине 1950-х годов [1]. В 1974 г. Паладе, Клод и Кристиан Де Дюв получили Нобелевскую премию по физиологии и медицине «за открытия, касающиеся структурной и функциональной организации клетки». Термин «рибосома» был предложен Ричардом Робертсом в 1958 вместо «рибонуклеобелковая частица микросомальной фракции» [
Рибосома — важнейший органоид живой клетки сферической или слегка овальной формы, диаметром 100-200. В эукариотических клетках рибосомы располагаются на мембранах эндоплазматического ретикулума, хотя могут быть локализованы и в неприкрепленной форме в цитоплазме. Синтез рибосом у эукариот происходит в специальной внутриядерной структуре — ядрышке. Рибосомы представляют собой нуклеопротеид. Рибосомная РНК составляет около 70 % всей РНК клетки.
Рибосома- место синтеза белка. Каждая рибосома сост из 2х частей (субъединиц) – большой и малой. Построены они из равных частей (по массе) белка ирнк. РНК входящ в сост рибосом наз рибосомальной. рРНК синтез в ядрышке.
Основным методом выделения рибосом является осаждение центрифугированием. Этот метод позволяет выделить два основных типа рибосом, которые называются 70S-рибосомами и 8OS-рибосомами.
— сведбсрг — единица, характеризующая скорость осаждения в центрифуге; чем больше число S. тем выше скорость осаждения). 70S — рибосомы обнаруживаются у прокариот и в хлоропластах и митохондриях эукариот. 8OS-рибосомы, несколько более крупные, находятся в цитоплазме эукариот. В процессе синтеза белка рибосомы движутся вдоль мРНК. Процесс идет более эффективно, если вдоль мРНК движется не одна, а несколько рибосом. Такие цепи рибосом на мРНК называют полирибосомами, или полисомами.
Источник: StudFiles.net
Эндоплазматическая сеть (ЭПС) – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название обусловлено тем, что её многочисленные элементы (цистерны, трубочки, пузырьки) образуют единую, непрерывную трехмерную сеть.
Степень развития ЭПС варьирует в различных клетках, и даже в разных участках одной и той же клетки, и зависит от функциональной активности клеток.
Различают две разновидности ЭПС (рис.4):
• гранулярную ЭПС (грЭПС) и
• гладкую, или агранулярную ЭПС (аЭПС), которые связаны между собой в переходной области.
Рис.4.
Гранулярная ЭПС образована мембранными трубочками и уплощенными цистернами, на наружной (обращенной в сторону гиалоплазмы) поверхности которых расположены рибосомы. Прикрепление рибосом происходит благодаря интегральным рецепторным белкам мембран грЭПС – рибофоринам. Эти же белки формируют гидрофобные каналы в мембране грЭПС для проникновения синтезированной белковой цепочки в просвет цистерн.
Основная функция грЭПС: сегрегация (отделение) вновь синтезированных белковых молекул от гиалоплазмы.
Таким образом, грЭПС обеспечивает:
• биосинтез белков, предназначенных для экспорта из клетки;
• биосинтез ферментов лизосом
• биосинтез мембранных белков.
Белковые молекулы накапливаются внутри просвета цистерн, приобретают вторичную и третичную структуру, а также подвергаются начальным посттрансляционным изменениям – гидроксилированию, сульфатированию, фосфорилированию и гликозилированию (присоединение к белкам олигосахаридов с образованием гликопротеинов).
ГрЭПС присутствует во всех клетках, но в наибольшей степени эта сеть развита в клетках, специализирующихся на белковом синтезе, таких как, клетки поджелудочной железы, вырабатывающих пищеварительные ферменты; фибробласты соединительной ткани, синтезирующих коллаген; плазматические клетки, продуцирующих иммуноглобулины. В этих клетках элементы грЭПС образуют параллельные скопления цистерн; при этом просвет цистерн часто расширен. Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов грЭПС.
Агранулярная ЭПС представляет собой трехмерную сеть мембранных трубочек, канальцев, пузырьков, на поверхности которых рибосомы отсутствуют.
Функции агрЭПС
• участие в синтезе липидов, в том числе мембранных, холестерина и стероидов;
• метаболизм гликогена;
• нейтрализация и детоксикация эндогенных и экзогенных токсичных веществ;
• накопление ионов Са (особенно в специализированной виде аЭПС – саркоплазматической сети мышечных клеток).
АгрЭПС хорошо развита:
• в клетках, активно продуцирующих стероидные гормоны – клетки коркового вещества надпочечников, интерстициальные гландулоциты яичка, клетки желтого тела яичника.
• в клетках печени, где её ферменты участвуют в метаболизме гликогена, а также в процессах, которые обеспечивают нейтрализацию и детоксикацию эндогенных биологически активных веществ (гормонов) и экзогенных вредных веществ (алкоголя, лекарственных веществ и др.).
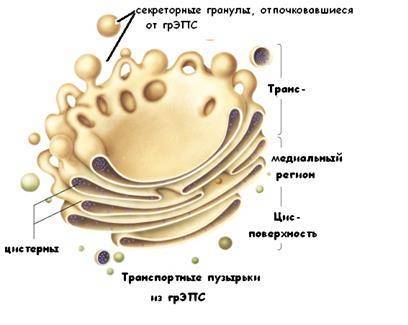
КОМПЛЕКС ГОЛЬДЖИ – мембранная органелла, образованная тремя основными элементами (рис.5): скоплениями уплощенных цистерн, мелкими (транспортными) пузырьками и конденсирующими вакуолями.
Комплекс этих элементов называется диктиосомой.
Рис.5.
Цистерны имеют вид изогнутых дисков с несколько расширенными периферическими отделами. Цистерны образуют группу в виде стопки из 3-30 элементов. От периферических расширений цистерн отщепляются пузырьки и вакуоли.
Пузырьки – мелкие (диаметр 40-80 нм), окруженные мембраной сферические элементы с содержимым умеренной электронной плотности. Вакуоли – крупные (диаметр 0.1-1.0 мкм), сферические образования, отделяющиеся от зрелой поверхности комплекса Гольджи в некоторых железистых клетках. Вакуоли содержат секреторный продукт, находящийся в процессе конденсации.
Комплекс Гольджи обладаетполярностью: в каждой диктиосоме выделяют две поверхности:
• формирующаяся (незрелая,илицис-поверхность) и
• зрелая (транс-поверхность).
Цис-поверхность выпуклой формы обращена в сторону ЭПС и связана с ней системой мелких транспортных пузырьков, отщепляющих от ЭПС. Таким образом, белки в транспортных пузырьках проникают через цис-поверхность.
Каждая группа медиальных цистерн внутри стопки отличается особым составом ферментов, и для каждой группы характерны свои реакции обработки белков. Обработанные вещества выходят в вакуолях с вогнутой транс-поверхности.
Источник: poznayka.org