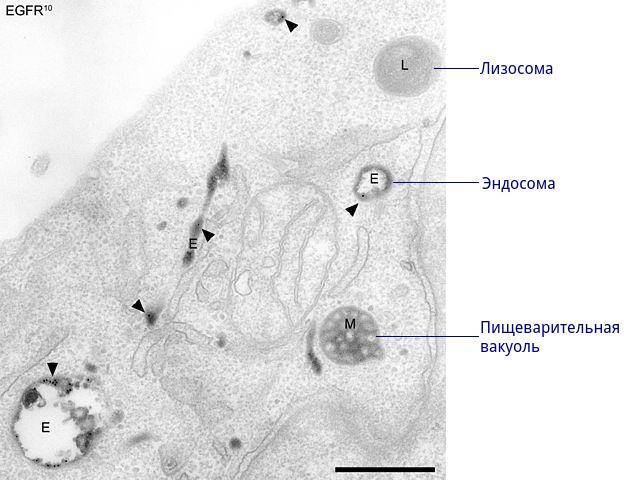
Вторичные лизосомы
Лизосомыпредставляют собой пузырьки диаметром от 100 нм до нескольких микрометров, которые обнаруживаются в цитоплазме клеток простейших, растений и животных. Они содержат широкий набор гидролитических ферментов, способных расщеплять любые биогенные вещества – белки, нуклеиновые кислоты, углеводы и липиды. Состав и количество лизосомальных гидролаз обладает видовой и тканевой специфичностью. Морфологически лизосомы неоднородны и претерпевают различные изменения при слиянии с другими органоидами. Для идентификации лизосом используется реакция на кислую фосфатазу.
Если приготовить гомогенат клеток печени и определить в нем активность кислой фосфатазы, то она окажется низкой. При обработке гомогената ультразвуком либо повторяющимися циклами замораживания-оттаивания активность фермента резко повышается. Объясняется это тем, что при обработке гомогената нарушается целостность мембран, гидролазы выходят из лизосом и активируются. Следовательно, гидролазы находятся в лизосомах в неактивном, латентном состоянии. Изучение явления латентности гидролаз и привело бельгийскго биохимика К. де Дюва к открытию лизосом (1955).
Латентность лизосомальных гидролаз обусловлена рядом факторов. Прежде всего, она объясняется наличием мембраны, которая препятствует взаимодействию ферментов с субстратами. Часть гидролаз встроено в мембрану и блокировано за счет связи с липидами. Другая часть ферментов инактивирована углеводами мембраны и матрикса. Кроме того, внутренняя среда лизосомы сильно закислена, тогда как максимальная активность гидролаз проявляется в слабо кислой среде.
Лизосомальная мембрана способна удерживать в органоиде не только ферменты, но и макромолекулярные субстраты. Низкомолекулярные продукты распада (свободные аминокислоты, сахара, нуклеотиды) выходят из лизосомы в гиалоплазму с помощью встроенных в мембрану специальных транспортных белков.
Морфологически лизосомы подразделяют на четыре типа – первичные лизосомы, вторичные лизосомы, аутофагосомы и остаточные тельца.
Первичные лизосомы образуются в пластинчатом комплексе в виде одномембранных пузырьков с бесструктурным содержимым. Они служат для временного хранения и инактивации гидролаз. Первичные лизосомы способны перемещаться в цитоплазме с помощью микротрубочек, а также сливаться с эндосомами и плазмолеммой. Диаметр их составляет 100-500 нм.
Вторичные лизосомы (фаголизосомы, пищеварительные вакуоли) образуются в результате слияния первичных лизосом с фагоцитарными или пиноцитозными вакуолями.
и этом наблюдается активация лизосомальных гидролаз, распад поступивших в клетку веществ и выведение низкомолекулярных веществ в гиалоплазму для включения в метаболические процессы клетки. Морфологически вторичные лизосомы отличаются от первичных лизосом более крупными размерами и наличием фагоцитируемого материала.
Аутолизосомы морфологически почти идентичны вторичным лизосомам, отличаясь только тем, что содержат внутри митохондрии, пластиды, рибосомы, включения и другие органоиды клетки или их фрагменты. Поэтому они рассматриваются как специальные органоиды, обеспечивающие аутофагоцитоз. Это явление наблюдается в клетках растений и животных, например, при физиологической регенерации внутриклеточных структур и гидролизе запасных питательных веществ.
Остаточные тельца (телолизосомы) представляют собой лизосомы с уплотненным структурированным материалом. Они содержат меньше гидролаз и служат местом накопления липидов, пигментов и других продуктов метаболизма.
Лизосомы представляют особый тип мембранного органоида, который осуществляет в клетке процессы распада. В специализированных клетках лизосомы могут обеспечивать внутриклеточное пищеварение, защиту от микроорганизмов и их продуктов, реконструкцию клетки. При осуществлении этих функций лизосомы взаимодействуют с другими мембранными органоидами – ПС, пластинчатым комплексом, эндосомами, включениями и плазмолеммой.
Функция внутриклеточного пищеварения, которое осуществляется с помощью лизосом, особенно выражена у простейших и беспозвоночных. Пищеварительные клетки имеются, например, в кишечном эпителии у моллюсков и клещей. Они содержат стабильные пиноцитозные вакуоли, которые постоянно снабжаются лизосомальными ферментами.
Защитная роль клеток-фагоцитов была обнаружена И. И. Мечниковым у беспозвоночных еще в конце XIX в. У позвоночных он описал два типа фагоцитов: микрофаги и макрофаги. В цитоплазме микрофагов содержится большое количество упакованных в мембрану гранул, которые представляют собой не что иное, как разновидность первичных лизосом. При активации клетки из них выходят гидролазы, которые разрушают внеклеточные субстраты. Микрофаги также способны к ограниченному фагоцитозу бактериальных клеток. К микрофагам у млекопитающих относятся нейтрофильные гранулоциты крови, которые обеспечивают защиту организма от бактерий. Макрофаги отличаются крупными размерами, способностью к амебоидному движению и активному фагоцитозу. Они содержат интенсивно функционирующие вторичные лизосомы, которые, кроме полного разрушения поступающих извне веществ, участвуют также в частичном протеолизе белковых антигенов для их последующего представления другим иммунокомпетентным клеткам. У млекопитающих к макрофагам относятся моноциты крови, которые после выхода из капилляров превращаются в гистиоциты соединительной ткани, альвеолярные и перитонеальные макрофаги, остеокласты костной ткани, клетки Купфера в печени и другие формы тканевых макрофагов.
Реконструктивная функция лизосом широко распространена как у животных, так и у растений. Она заключается в ограниченной аутофагии цитоплазматических структур для регуляции обмена веществ, развития тканей и органов, а также в условиях недостатка энергии и питательных веществ. Примерами могут служить гидролиз запасных питательных веществ при прорастании семян, частичный протеолиз тироглобулина эпителиальными клетками щитовидной железы с образованием гормонов трийодтиронина и тироксина, разрушение межклеточных контактов при физиологической регенерации ороговевающего эпителия. Особенно отчетливо эта функция лизосом проявляется в разрушении ими цитоплазматических структур в условиях голодания (эндогенное питание).
Значение лизосом для нормального функционирования организма человека подчеркивается существованием “болезней накопления”, которые обусловлены мутациями, полностью или частично нарушающими работу лизосомальных гидролаз. В результате в лизосомах наблюдается избыточное накопление соответствующих субстратов, что приводит к изменению размеров, формы и тонкой структуры этого органоида. При болезни Уолмена, например, отсутствует кислая липаза, что приводит к накоплению в лизосомах эфиров холестерина и триглицеридов и формированию липидных включений. Болезнь Помпа связана с недостатком гликозидазы, что вызывает накопление гликогена. Особенно тяжелым заболеванием является псевдохерлерова дистрофия, при которой в фибробластах отсутствуют почти все лизосомальные ферменты, кроме протеаз. В результате блокируется обмен таких компонентов соединительных тканей как протеогликаны и сложные липиды.
Источник: helpiks.org
Лизосомы (от гр. lysis — «разложение, растворение, распад» и soma — «тело») — это пузырьки которые имеют средний диаметр от двухсот до четырёхсот микромиллиметров. У них одномембранная оболочка, которая снаружи иногда покрыта волокнистым белковым слоем. Лизосомы содержат в себе набор ферментов (кислых гидролаз), которые производят гидролитическое (в присутствии воды) расщепление веществ (нуклеиновых кислот, белков, жиров, углеводов) при низких значениях рН.
Основная функция — внутриклеточное переваривание различных химических соединений и клеточных структур. Существуют:
- первичные или же неактивные,
- вторичные лизосомы, в которых и осуществляется процесс переваривания.
Вторичные лизосомы образуются из первичных. Они разделяются на:
- гетеролизосомы,
- аутолизосомы.
В гетеролизосомах проходит процесс переваривания материала, который поступает в клетку извне путем активного транспорта (пиноцитоза и фагоцитоза). В аутолизосомах разрушаются собственные клеточные структуры, завершившие свой жизненный цикл.
Вторичные лизосомы, которые уже перестали переваривать материал, называются остаточными тельцами. В них нет гидролаз, содержится непереваренный материал. При нарушении целостности мембраны лизосом или при заболевании клетки гидролазы начинают поступать внутрь клетки из лизосом и производят ее самопереваривание (автолиз). Этот же процесс лежит в основе процесса естественной гибели всех клеток (апоптоза).
Лизосомы, строение и функции которых являются внутриклеточно формирующимися секреторные вакуоли, заполненные гидролитическими ферментами, необходимые для процессов фаго- и аутофагоцитоза. На светооптическом уровне лизосомы можно распознать и определить степень их развития в клетке по активности гистохимической реакции на кислую фосфатазу — ключевой лизосомальный энзим.
При электронной микроскопии лизосомы выглядят как пузырьки, ограниченные от гиалоплазмы мембраной. Существует четыре основных вида лизосом:
- первичные илизосомы,
- вторичные лизосомы,
- аутофагосомы,
- остаточные тельца.
Первичные лизосомы— это мелкие мембранные пузырьки, которые имеют диаметр около ста нм, заполненные гомогенным мелкодисперсным содержимым, являющим собой набор гидролитических ферментов. В лизосомах есть около сорока ферментов. Например, протеазы, нуклеазы, гликозидазы, фосфорилазы, сульфатазы. Их оптимальный режим действия рассчитан на кислую среду (рН 5). Лизосомальные мембраны содержат специальные белки-носители для транспорта из лизосомы в гиалоплазму продуктов гидролитического расщепления — аминокислот, сахаров и нуклеотидов. Мембрана лизосом устойчива по отношению к гидролитическим ферментам.
Вторичные лизосомы образуются при слиянии первичных лизосом с эндоцитозными либо с пиноцитозными вакуолями. Если сказать иначе, то вторичные лизосомы — это внутриклеточные пищеварительные вакуоли, ферменты которых поставляются первичными лизосомами, а материал для переваривания — эндоцитозной (пиноцитозной) вакуолью. Строение вторичных лизосом очень разное и может меняться во время гидролитического расщепления содержимого. Ферменты лизосом расщепляют попавшие в клетку биологические вещества, после чего образуются мономеры, которые транспортируются через мембрану лизосомы в гиалоплазму, где утилизируются или включаются в разнообразные синтетические и метаболические реакции. Если взаимодействию с первичными лизосомами и гидролитическому расщеплению их ферментами подвергаются собственные структуры клетки (стареющие органеллы, включения и пр.), формируется аутофагосома. Аутофагоцитоз считается естественным процессом в жизнедеятельности клетки и ей отводится важная роль в обновлении ее структур при внутриклеточной регенерации.
Источник: vseobiology.ru
Строение лизосомы
Элементарная мембрана лизосомы отграничивает от цитоплазмы несколько десятков гидролитических (пищеварительных) ферментов, расщепляющих белки, жиры, углеводы и нуклеиновые кислоты. Ферменты относятся к группам протеаз, липаз, нуклеаз, фосфатаз и др.
В отличие от гиалоплазмы, внутренняя среда лизосом имеет кислую реакцию, а содержащиеся здесь ферменты активны только при низком pH.
Изоляция ферментов лизосом необходима, иначе, оказавшись в цитоплазме, они могут разрушить клеточные структуры.
Образование лизосом
Лизосомы образуются в комплексе Гольджи. Ферменты (по-сути белки) лизосом синтезируются на шероховатой эндоплазматической сети, после чего транспортируются в Гольджи с помощью везикул (пузырьков, ограниченных мембраной). Здесь белки модифицируются, приобретают свою функциональную структуру, упаковываются в другие пузырьки – первичны лизосомы, – которые отрываются от аппарата Гольджи. Далее, превращаясь во вторичные лизосомы, выполняют функцию внутриклеточного переваривания. В некоторых клетках первичные лизосомы секретируют свои ферменты за пределы цитоплазматической мембраны.
Функции лизосом
О функциях лизосом говорит уже их название: lysis — расщепление, soma — тело.
При попадании в клетку питательных веществ, каких-либо микроорганизмов лизосомы принимают участие в их переваривании. Кроме того, они разрушают ненужные структуры самой клетки и даже целые органы организмов (например, хвост и жабры в процессе развития многих земноводных).
Ниже дается описание основным, но не единственным функциям лизосом.
Переваривание частиц, поступивших в клетку путем эндоцитоза
Путем эндоцитоза (фогоцитоза и пиноцитоза) в клетку поступают относительно крупные материалы (питательные вещества, бактерии и др.). При этом цитоплазматическая мембрана впячивается внутрь клетки, во впячивание попадает структура или вещество, после чего впячивание отшнуровывается во внутрь, и образуется пузырек (эндосома), окруженный мембраной, – фагоцитарный (с твердыми частицами) или пиноцитарный (с растворами).
С таким пузырьком далее сливается первичная лизосома. В результате образуется вторичная лизосома, в которой происходит процесс переваривания (происходят реакции ферментативного расщепления).

Подобным способом может происходить усваивание пищи (например, у амеб). В данном случае вторичную лизосому также называют пищеварительной вакуолью. Переваренные вещества поступают из вторичной лизосомы в цитоплазму. Другой вариант — переваривание попавших в клетку бактерий (наблюдается у фагоцитов — специализированных для защиты организма лейкоцитов).
Оставшиеся во вторичной лизосоме ненужные вещества удаляются из клетки путем экзоцитоза (обратен эндоцитозу). Лизосома с непереваренными подлежащими удалению веществами называются остаточным тельцем.
Автофагия
Путем автофагии (аутофагии) клетка избавляется от ненужных ей собственных структур (различных органелл и др.).
Сначала такой органоид окружается элементарной мембраной, отделившейся от гладкой ЭПС. После этого образовавшийся пузырек сливается с первичной лизосомой. Образуется вторичная лизосома, которая называется автофагической вакуолью. В ней происходит переваривание клеточной структуры.
Автофагия особенно выражена в клетках, находящихся в процессе дифференциации.
Автолиз
Под автолизом понимают саморазрушение клетки. Характерен при метаморфозах, омертвении тканей.
Автолиз наступает, когда содержимое многих лизосом высвобождается в цитоплазму. Обычно в достаточно нейтральной среде гиалоплазмы ферменты лизосом, которым необходима кислая среда, становятся неактивными. Однако, когда разрушается много лизосом, то кислотность среды повышается, а ферменты остаются активными и расщепляют клеточные структуры.
Источник: biology.su
История открытия[править | править код]
В 1949—1952 годах биохимик Кристиан де Дюв и его студенты, изучавшие действие инсулина в клетках печени крыс, случайно обнаружили неожиданное различие в активности кислой фосфатазы в зависимости от способа выделения. Кислая фосфатаза использовалась ими в качестве стандарта, основным предметом их изучения был фермент глюкозо-6-фосфатаза, вовлечённый в метаболизм инсулина. В ходе экспериментов выяснилось, что при фракционировании клеточного содержимого на центрифуге кислая фосфатаза была ассоциирована с микросомальной фракцией, но проявляла только десятую часть активности в сравнении с простым клеточным экстрактом, причём после нескольких дней хранения микросомальной фракции в холодильнике активность кислой фосфатазы возрастала. При обнаружении этого феномена первым объяснением было то, что произошла какая-то техническая ошибка. Однако повторение эксперимента неизменно воспроизводило первоначальную картину. Это позволило предположить существование неких окружённых мембраной клеточных частиц, которые содержат внутри себя фермент. С 1952 по 1955 год было открыто ещё несколько кислых гидролаз, связанных с микросомальной фракцией. В 1955 году, который считается годом открытия лизосом, К. де Дюв предложил название «лизосома» для клеточной органеллы, которая окружена мембраной, внутри которой поддерживается низкий pH и внутри которой находится ряд ферментов, оптимально работающих в кислой среде[7][8]. В том же 1955 году американский цитолог Алекс Новиков (англ.)русск. из Вермонтского университета США, блестяще владевший техникой микроскопии, посетил лабораторию К. де Дюве и смог получить первые электронные фотографии этих органелл, используя препарат частично очищенных лизосом. Позднее в 1961 году Алекс Новиков с помощью гистохимического выявления кислой фосфатазы и электронной микроскопии подтвердил локализацию этого фермента в лизосомах[9][10]. В 1963 году бельгийский биохимик Генри Хэрс, ранее работавший в группе К. де Дюве, обнаружил недостаточность лизосомного фермента α-глюкозидазы у пациентов с болезнью Помпе и высказал предположение о связи других генетических заболеваний с нарушением работы лизосом[11]. В настоящее время более 50 наследственных заболеваний связывают с лизосомной недостаточностью[12].
В 1974 году за свой вклад в раскрытие структурной и функциональной организации клетки К. де Дюв был удостоен Нобелевской премией по медицине[13].
Признаки лизосом[править | править код]
Лизосомы являются гетерогенными по форме, размеру, ультраструктурным и цитохимическим особенностям. В клетках животных размер лизосом составляет обычно менее 1 мкм, хотя в некоторых типах клеток, например, в макрофагах, размер лизосом может превышать несколько микрон. Лизосомы, как правило, имеют сферическую, овальную, иногда тубулярную форму[14]. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных). Лизосомы у животных обычно составляют не более 5 % внутриклеточного объёма[15].
Один из признаков лизосом — наличие в них ряда ферментов (кислых гидролаз), способных расщеплять белки, углеводы, липиды и нуклеиновые кислоты. К числу ферментов лизосом относятся катепсины (тканевые протеазы), кислая рибонуклеаза, фосфолипаза и др. Кроме того, в лизосомах присутствуют ферменты, которые способны отщеплять от органических молекул сульфатные (сульфатазы) или фосфатные (кислая фосфатаза) группы. Всего полость лизосомы содержит около 60 растворимых кислых гидролитических ферментов[2].
Для лизосом характерна кислая реакция внутренней среды, которая обеспечивает оптимум работы лизосомных гидролаз[14]. Обычно pH в лизосомах составляет около 4,5-5, то есть концентрация протонов в них на два порядка выше, чем в цитоплазме. Это обеспечивается активным транспортом протонов, который осуществляет встроенный в мембраны лизосом белок-насос протонная АТФаза[15]. Помимо протонного насоса в мембрану лизосом встроены белки-переносчики для транспорта в цитоплазму продуктов гидролиза макромолекул: аминокислот, сахаров, нуклеотидов, липидов[16].
Высокая активность кислой фосфатазы ранее использовалась как один из маркеров лизосом. В настоящее время более надежным маркером считается присутствие специфических мембранных гликопротеидов — LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома.
Образование лизосом и их типы[править | править код]
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе[17]. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.
Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путём фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
- Ранняя эндосома — в неё поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
- Поздняя эндосома — в неё из ранней эндосомы поступают пузырьки с материалом, поглощённом при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.
- Лизосома — в неё из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.
- Фагосома — в неё попадают более крупные частицы (бактерии и т. п.), поглощённые путём фагоцитоза. Фагосомы обычно сливаются с лизосомой.
- Аутофагосома — окружённый двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.
- Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окружённые одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путём отпочковывания от ядерной оболочки.
- Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал (в частности, липофусцин). В нормальных клетках сливаются с наружной мембраной и путём экзоцитоза покидают клетку. При старении или патологии накапливаются.
Функции лизосом[править | править код]
Функциями лизосом являются:
- переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток)
- аутофагия — уничтожение ненужных клетке структур, к примеру, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки
- автолиз — самопереваривание клетки, приводящее к её гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
- растворение внешних структур (см, например, остеокласты)
Внутриклеточное пищеварение и участие в обмене веществ[править | править код]
У многих протистов и у животных, имеющих внутриклеточное пищеварение, лизосомы участвуют в переваривании пищи, захваченной путём эндоцитоза. При этом лизосомы сливаются с пищеварительными вакуолями. У протистов непереваренные остатки пищи обычно удаляются из клетки при слиянии пищеварительной вакуоли с наружной мембраной.
Многие клетки животных, у которых преобладает полостное пищеварение (например, хордовые) получают питательные вещества из межклеточной жидкости или плазмы крови с помощью пиноцитоза. Эти вещества также вовлекаются в обмен веществ клетки после их переваривания в лизосомах. Хорошо изученный пример такого участия лизосом в обмене веществ — получение клетками холестерина. Холестерин, приносимый кровью в виде ЛПНП, поступает внутрь пиноцитозных везикул после соединения ЛПНП с рецепторами ЛПНП на мембране. Рецепторы возвращаются к мембране из ранней эндосомы, а ЛПНП поступают в лизосомы. После этого ЛПНП перевариваются, а высвободившийся холестерин через мембрану лизосом поступает в цитоплазму.
Косвенно лизосомы участвуют в обмене, обеспечивая десенсибилизацию клеток к воздействию гормонов. При длительном действии гормона на клетку часть рецепторов, связавших гормон, поступают в эндосомы и затем деградируют внутри лизосом. Снижение числа рецепторов понижает чувствительность клетки к гормону.
Для крупных вакуолей растений характерна запасающая функция — в них могут накапливаться ионы, пигменты (например, антоцианы), вторичные метаболиты, белки (в алейроновых зернах эндосперма злаков). Внутри вакуолей (например, в прорастающих семенах) у растений происходят и процессы переваривания запасенных белков.
Аутофагия[править | править код]
Обычно различают два типа аутофагии — микроаутофагия и макроаутофагия. При микроаутофагии, как при образовании мультивезикулярных телец, образуются впячивания мембраны эндосомы или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путём клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды; так, у дрожжей описана микроаутофагия пероксисом и частичная микроаутофагия ядер, при которой клетка сохраняет жизнеспособность.
При макроаутофагии участок цитоплазмы (часто содержащий какие-либо органоиды) окружается мембранным компартментом, похожим на цистерну эндоплазматической сети. В результате этот участок оказывается отгорожен от остальной цитоплазмы двумя мембранами. Затем такая аутофагосома сливается с лизосомой, и её содержимое переваривается. Видимо, макроаутофагия также неизбирательна, хотя часто подчеркивается, что с помощью неё клетка может избавляться от «отслуживших свой срок» органоидов (митохондрий, рибосом и др.).
Третий тип аутофагии — шаперон-зависимая. При этом способе происходит направленный транспорт частично денатурировавших белков из цитоплазмы сквозь мембрану лизосомы в её полость.
Автолиз[править | править код]
Ферменты лизосом нередко высвобождаются при разрушении мембраны лизосомы. Обычно при этом они инактивируются в нейтральной среде цитоплазмы. Однако при одновременном разрушении всех лизосом клетки может произойти её саморазрушение — автолиз. Различают патологический и обычный автолиз. Распространенный вариант патологического автолиза — посмертный автолиз тканей.
В норме процессы автолиза сопровождают многие явления, связанные с развитием организма и дифференцировкой клеток. Так, автолиз клеток описывается как механизм разрушения тканей у личинок насекомых при полном превращении, а также при рассасывании хвоста у головастика. Правда, эти описания относятся к периоду, когда различия между апоптозом и некрозом ещё не были установлены, и в каждом случае требуется выяснять, не лежит ли на самом деле в основе деградации органа или ткани апоптоз, не связанный с автолизом.
У растений автолизом сопровождается дифференциация клеток, которые функционируют после смерти (например, трахеид или члеников сосудов). Частичный автолиз происходит и при созревании клеток флоэмы- члеников ситовидных трубок.
Клиническое значение[править | править код]
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером лизосомных болезней накопления может служить болезнь Гоше, болезнь Помпе, Болезнь Тея — Сакса. Всего известно более 50 наследственных заболеваний, связанных с нарушением функции лизосомы[12].
Повреждение лизосом некротизированных клеток, в том числе гранулоцитов, даёт начало воспалительному процессу[18].
См. также[править | править код]
- Вакуоль
- Фагоцитоз
- Эндоцитоз
- Лизосомные болезни накопления
- Восковидные липофусцинозы нейронов
Источник: ru.wikipedia.org