Неоген период
Оглавление
Общие сведения о животных
Разделение классификации животных
Появление и эволюция животных
Протерозой. Довендская биота. Животный мир вендского периода (эдикария)
Фанерозой. Животный мир кембрийского периода. Кембрийский взрыв
Животный мир ордовикского периода
Животный мир силурийского периода
Животный мир девонского периода
Животный мир каменноугольного периода
Животный мир пермского периода
Животный мир триасового периода
Животный мир юрского периода
Животный мир мелового периода
Животный мир палеогенового периода
Животный мир неогенного периода
Животный мир миоценовой эпохи
Животный мир плиоценовой эпохи
Животный мир четвертичного периода
2.12.2. Животный мир неогенового периода (23,0 — 2,588 млн. лет назад)
Неогеновый период (23,0 — 2,588 млн.лет назад).
 Рис. 2.12.1. НеогенНеогеновый период (в переводе — новорожденный рис. 2.12.1) подразделяется на два отдела миоцен (23,0 — 5,33 млн. лет назад) и плиоцен (5,33 — 2,588 млн. лет назад). Климат неогенового периода был довольно теплым, и влажным, однако несколько более прохладным по сравнению с климатом палеогенового периода. В конце неогена он постепенно приобретает современные черты.
Рис. 2.12.1. НеогенНеогеновый период (в переводе — новорожденный рис. 2.12.1) подразделяется на два отдела миоцен (23,0 — 5,33 млн. лет назад) и плиоцен (5,33 — 2,588 млн. лет назад). Климат неогенового периода был довольно теплым, и влажным, однако несколько более прохладным по сравнению с климатом палеогенового периода. В конце неогена он постепенно приобретает современные черты.
Среди морских беспозвоночных в это время преобладают двустворчатые и брюхоногие моллюски, морские ежи. Мшанки и кораллы на юге Европы образуют рифы. Прослеживаются арктические зоогеографические провинции: северная, включавшая Англию, Нидерланды и Бельгию, южная — Чили, Патагонию и Новую Зеландию.
Сильно распространилась солоноватоводная фауна. Ее представители населяли большие мелководные моря, образовавшиеся на материках в результате наступления неогенового моря. В этой фауне совершенно отсутствуют кораллы, морские ежи и звезды. Моллюски по количеству родов и видов значительно уступают моллюскам, населявшим океан с нормальной соленостью. Однако по численности особей они во много раз превосходят океанских. Раковины небольших по размерам солоноватоводных моллюсков буквально переполняют отложения этих морей. Рыбы уже совершенно не отличаются от современных.
Рис. 2.12.2. Махайроды (Machairodus)Похожим на современный становится и сухопутный органический мир. Примитивных креодонтов вытесняют медведи, гиены, куницы, собаки, барсуки. Будучи более подвижными и имея более сложную организацию, они приспособились к разнообразным условиям жизни, перехватывали добычу у креодонтов и сумчатых хищников, а иногда и питались ими.
Наряду с видами, которые, несколько изменившись, дожили до нашего времени, появились и виды хищников, которые вымерли в неогене. К ним в первую очередь относится саблезубый тигр (рис. 2.12.2).
У потомков палеогеоновых мерикгиппусов — гиппарионов были уже такие зубы, как у современной лошади. Их небольшие боковые копыта не касались земли. Копыта же на средних пальцах делались все более крупными и широкими. Они хорошо удерживали животных на твердой почве, давали им возможность разрывать снег, чтобы извлечь из-под него корм, а так же защищаться от хищников.
Рис. 2.12.3. Мегатерий (Megatherium)Наряду с североамериканским центром развития лошадей существовал и европейский. Однако в Европе древние лошади вымерли в начале олигоцена, не оставив потомков. Вероятнее всего они были истреблены многочисленными хищниками. В Америке же древние лошади продолжали развиваться. Впоследствии они дали настоящих лошадей, которые через Берингов перешеек проникли в Европу и Азию. В Америке лошади вымерли еще в начале плейстоцена, а крупные табуны современных мустангов, свободно пасущиеся в американских прериях, являются далекими потомками лошадей, привезенных испанскими колонизаторами. Таким образом, произошел своеобразный обмен лошадьми между Новым Светом и Старым Светом.
В Южной Америке обитали гигантские ленивцы — мегатерии (до 8 м в длину рис. 2.12.3). Становясь на задние лапы, они объедали листья деревьев. Мегатерии имели толстый хвост, низкий череп с небольшим мозгом. Их передние лапы были намного короче задних. Будучи неповоротливыми, они становились легкой добычей для хищников и поэтому полностью вымерли, не оставив потомков.
Изменение климатических условий привело к образованию обширных степей, что благоприятствовало развитию копытных. От маленьких безрогих оленей, живших на болотистой почве, произошли многочисленные парнокопытные — антилопы, козлы, бизоны, бараны, газели, чьи прочные копыта были хорошо приспособлены к быстрому бегу в степях. Когда парнокопытных развелось такое количество, что начала ощущаться нехватка пищи, часть их освоила новые места обитания: скалы, лесостепи, пустыни. От живших в Африке жирафообразных безгорбых верблюдов произошли настоящие верблюды, заселившие пустыни и полупустыни Европы и Азии. Горб с питательными веществами позволял верблюдам длительное время обходиться без воды и пищи.
Рис. 2.12.4. МегалоцерасЛеса заселяли настоящие олени, из которых одни виды встречаются и в наши дни, а другие, например мегалоцерасы (рис. 2.12.4), которые были в полтора раза крупнее обычных оленей, полностью вымерли.
В лесостепных зонах обитали жирафы, вблизи озер и болот — бегемоты, свиньи, тапиры. В густых кустарниковых зарослях жили носороги, муравьеды.
Среди хоботных появляются мастодонты с прямыми длинными клыками и настоящие слоны.
На деревьях живут лемуры, обезьяны, человекообразные обезьяны. Некоторые лемуры перешли к наземному образу жизни. Передвигались они на задних ногах. Достигали 1,5 м в высоту. Питались главным образом плодами и насекомыми.
Рис. 2.12.5. Динорнис (Dinornis)Жившая в Новой Зеландии гигантская птица динорнис достигала 3,5 м в высоту (рис. 2.12.5). Голова и крылья у динорниса были маленькими, клюв недоразвитым. Он передвигался по земле на длинных сильных ногах. Дожил динорнис до четвертичного периода и, очевидно, был истреблен человеком.
В неогеновый период появляются дельфины, тюлени, моржи — виды, живущие и в современных условиях.
В начале неогенового периода в Европе и Азии было много хищных животных: собак, саблезубых тигров, гиен. Среди травоядных преобладали мастодонты, олени, однорогие носороги.
В Северной Америке хищные были представлены собаками и саблезубыми тиграми, а травоядные — титанотериями, лошадьми и оленями.
Южная Америка была несколько изолированной от Северной. Представителями ее фауны были сумчатые, мегатерии, ленивцы, броненосцы, широконосые обезьяны.
В верхнемиоценовый период между Северной Америкой и Евразией происходит обмен фауной. Много животных переселилось с материка на материк. Северную Америку заселяют мастодонты, носороги, хищники, а в Европу и Азию переселяются лошади.
С началом лигоцена в Азии, Африке и Европе расселяются безрогие носороги, мастодонты, антилопы, газели, свиньи, тапиры, жирафы, саблезубые тигры, медведи. Однако во второй половине плиоцена климат на Земле сделался прохладным, и такие животные, как мастодонты, тапиры, жирафы, переселяются на юг, и на их месте появляются быки, бизоны, олени, медведи.
В плиоцене связь между Америкой и Азией прервалась. Одновременно возобновилась связь между Северной и Южной Америкой. Североамериканская фауна переселилась в Южную Америку и постепенно вытеснила ее фауну. Из местной фауны остались лишь броненосцы, ленивцы и муравьеды, распространились медведи, ламы, свиньи, олени, собачьи, кошачьи.
Австралия была изолирована от других материков. Следовательно, значительных изменений в фауне там не произошло.[1]
2.12.1. Животный мир миоценовой эпохи (25 — 5 млн. лет назад)
В миоцене, с началом закрытия восточного Тетиса, изменения уровня моря определяли возможность интенсивных межконтинентальных фаунистических обменов между Евразией, Африкой, Индийским субконтинентом и восточносредиземноморской областью. В пределах Евразийской зоогеографической области сформировалась единая Европейско-Сибирская подобласть с южной границей в Азии приблизительно по 40oсеверной широты, ставшая гигантской ареной эволюции многих групп млекопитающих в миоцене.
Рис. 2.12.1.1. Анхитерий (Anchitherium)В конце раннего миоцена, впервые после длительного периода взаимной изоляции, происходил значительный обмен между Африкой и Евразией. Главным эффектом этого обмена было изменение африканской фауны: в ее составе появились кошачьи, собачьи, куньи, носороги, халикотерии, свиньи. Вероятно, в это же время из Африки на Индийский субконтинент эмигрировали хоботные. Первые мастодонты вселились в Европу и Северную Азию в раннем миоцене (MN3). В это же время в Евразию из Северной Америки иммигрировали анхитерии (рис. 2.12.1.1). Наиболее значительным событием в истории раннемиоценовых грызунов Европы была иммиграция азиатских по происхождению хомяков современного облика (Eumyarion, Democricetodon, Fahlbuschia, Megacricetodon, Cricetodon (рис. 2.12.1.2), Lartetomys) в среднем орлеании. С их появлением эомиидно-глиридная ассоциация трансформировалась в крицетидно-глиридную, причем во многих комплексах доминирующей группой были именно Cricetidae. В таком виде европейская фауна грызунов просуществовала вплоть до раннего валлезия (MN9). В среднем орлеании в Африку из Евразии проникли первые полорогие и жирафовые.
Рис. 2.12.1.2. Cricetodon soriaeМиоцен – время вымирания креодонтов. Однако в Азии в среднем миоцене еще присутствуют 2 рода: самый крупный представитель Proviverrinae — Dissopsalis (размером с шакала) и очень крупный представитель Hyaenodontinae — Hyainailouros (размером с тигра). Интересно отметить, что эти оба рода общие для Азии и Африки, а Hyainailouros присутствует еще и в Европе. Это подтверждает обмен фауны креодонтов между Азией и Африкой через Европу в олигоцене. Вымирание креодонтов совпадает с адаптивной радиациейCarnivora, которые являются конкурентной группой. Вымиранию креодонтов, вероятно, способствовали этологические особенности группы. Морфофункциональный анализ мозга Neohyaenodon horridus (Hyaenodontini) показал сильное морфологическое развитие вентро-каудальной зоны височной доли мозга (Савельев, Лавров, 2001). Следствием этого является повышение контроля за спонтанной активностью и агрессией. Этот крупный хищник был менее агрессивным, чем современные крупные представители отряда хищных (Carnivora).
Рис. 2.12.1.3. Pliocyon преследует верблюдовС конца раннего миоцена уже можно четко обосновать провинциальное биогеографическое подразделение евразиатских и североафриканских териофаун. Для фаун млекопитающих всех европейских провинций в целом было характерно преобладание сообществ закрытых лесных обстановок, явно субтропических в южных районах. Суб-Паратетисная провинция характеризовалась наличием гигантского “экологического коридора” открытых ландшафтов на территории от восточносредиземноморской Европы на западе через Малую Азию, Ближний Восток и северную Аравию до Афганистана на востоке и Китая на северо-востоке. Вероятно, именно через нее осуществлялись основные фаунистические обмены между Евразией и Африкой.
Взаимное влияние столкнувшихся в Евразии и Африке в миоцене различных по области происхождения экологических конкурентов вызвало бурную эволюцию ряда групп (ежовых, землеройковых, кротовых, пищуховых, беличьих, тушканчикообразных, хомякообразных и многих других грызунов, антропоидных приматов, носороговых, жвачных, хоботных, куньих, собачьих, гиеновых, кошачьих и др.) и вымирание последних “палеогеновых” реликтов (ежей-амфехинин, корнезубых зайцеобразных, аплодонтовых-просциурин, ктенодактилид-татаромиин, тахиориктоидид, гигантских носорогов, кайнотериев, креодонтов, амфиционид (рис. 2.12.1.3) и др.). К концу миоцена сформировались почти все группировки млекопитающих надродового ранга, а также очень многие из рецентных родов. В добавление к ним существовало множество впоследствии вымерших форм, так что с точки зрения териологии миоцен по праву можно считать эпохой великого разнообразия, в формировании которого одна из ведущих ролей принадлежала территории Азии. [2]
Рис. 2.12.1. 4. Зубы — большой белой акулы и кархарадонаОдна из крупнейших среди современных акул — большая белая акула, достигающая длины 10м, кажется карликом в сравнении с одним из своих предков, кархародоном мегалодоном. Эта рыба обитала на Земле с миоценовой по плейстоценовую эпоху. И хотя кархародон мегалодон известен нам лишь по ископаемым зубам, ученые все же смогли воссоздать его внешний облик. Взгляните на фотографии двух зубов (рис. 2.12.1.4) изображенных в натуральную величину. Тот, что слева, принадлежит взрослой большой белой акуле. Другой же — ископаемый зуб кархародона. Ученые сопоставили оба зуба и, основываясь на известных им размерах большой белой акулы, прикинули, что длина кархародона от носа до кончика хвоста должна была составлять не менее 20 м (рис. 2.12.1.5).
Рис. 2.12.1.5. КархарадонРаспространение степей и постепенное исчезновение лесов в миоцене сильно повлияли на рост численности популяций травоядных млекопитающих. Меньше деревьев — меньше энергии, которую они «отдают» животным.
Представьте себе на минуту обычное дерево. Большую часть своей энергии оно расходует на создание для себя опоры. С этой целью у него развивается специальный твердый ствол и множество ветвей.
В умеренном климате животные за год могут употребить в пищу лишь очень незначительную часть съедобного вещества, вырабатываемого деревом.
На листопадных деревьях (которые ежегодно сбрасывают листву) листья вообще растут всего шесть месяцев в году, а плоды и семена (за исключением орехов) появляются только на несколько недель. Значит, леса умеренного пояса могут за год прокормить лишь ограниченное число животных.
Резкое увеличение количества травы на суше в миоценовую эпоху означало, по сути, возникновение принципиально нового источника пищи. Однако поначалу травоядным млекопитающим было непросто воспользоваться ресурсами этой бездонной кладовой. Некоторые их виды даже вымерли, поскольку не смогли приспособиться к травяному рациону. Млекопитающим, зубы которых были рассчитаны на пережевывание мягкой листвы, оказалось нелегко перейти на такой жесткий и волокнистый корм, как трава. Для таких животных травяная «диета» означала постоянное усиленное жевание, и их зубы быстро изнашивались, создавая им серьезнейшие проблемы. Ведь млекопитающие не обладают неограниченным запасом зубов, а беззубые челюсти означают для них неминуемую голодную смерть.
Несмотря на все это, в миоцене млекопитающие стали гораздо многочисленнее и разнообразнее, среди них возникло множество травоядных. К этому времени «жвачный» желудок превратился в идеальный механизм для переваривания травы. В результате в миоцене произошел своего рода взрыв, породивший новые виды травоядных, способных «жевать жвачку». Жвачные животные могут набивать себе животы громадным количеством пищи, переварить которую можно и позднее. При этом если на жвачное животное нападет хищник, оно может удрать от него, унося с собой запас корма на несколько дней вперед. Оказавшись в безопасности, животное может без лишней спешки заняться перевариванием съеденного.
В ходе этой своеобразной «жвачной революции» резко возросла численность предков нынешних антилоп, буйволов, оленей, жирафов и овец. В Северной Америке обитали антилопообразные вилороги — их причудливые рога росли у них на кончике носа.
Рис. 2.12.1.6. МеригиппусЧтобы совладать с травяным кормом, прежние зубы травоядных претерпели два основных изменения:
Во-первых, узор из бугорков на жевательной поверхности зубов значительно усложнился, и они стали самозатачивающимися. Теперь на зубах, по мере стирания, образовывались ряды прочных эмалевых ребер, которые оставались острыми всю жизнь животного.
Во-вторых, у отдельных зубов развились гораздо более широкие коронки (та часть зуба, что выдается над десной) и они сделались крупнее.
Эти новшества, как и появление корневых отверстий (которые обеспечивали приток крови к растущей части зуба), способствовали тому, что зубы животных стали расти всю их жизнь. Теперь постоянное трение зубов друг о друга больше уже не приводило к их преждевременному износу.
Необходимость приспособиться к преимущественно травяному корму — не единственная проблема, возникшая перед новыми миоценовыми травоядными. Не менее серьезные проблемы были связаны с жизнью на открытых равнинах, где травоядные постоянно находились на виду у хищников.
Чтобы выжить в подобных условиях, животные нуждались в хорошем круговом зрении, дающем широкий или даже 180-градусный обзор (при таком обзоре глаза располагаются по обеим сторонам головы, и животные видят и то, что впереди, и то, что сзади, оставаясь неподвижными). Такое зрение позволяло травоядным вовремя замечать опасность, грозящую с любого направления, что, в свою очередь, улучшало взаимодействие в стаде.
Чтобы стадо имело больше шансов уцелеть на открытой местности, в нем выработались специальные методы несения, «караульной службы» (когда отдельные животные смотрят по диагонали через все стадо), а также более совершенные системы сигналов и обмена информацией внутри стада.
Рис. 2.12.1.7. Мастадонт (Mammutidae)Длинные ноги также ценное подспорье при жизни на равнине. Они позволяют животному держать голову высоко над землей и тем самым лучше обозревать окрестности. Ну и, разумеется, длинные ноги служат своему хозяину незаменимым средством спасения в случае опасности. Поэтому в течение миоцена конечности травоядных животных постепенно приспособились к быстрому бегу.
Кости нижней части ноги со временем удлинились, а кости верхней, напротив, стали короче. Главные мышцы, ответственные за движение конечностей, также укоротились и располагались теперь выше, ближе к лопаткам и подвздошным костям, где они соединялись с туловищем животного.
При таком строении мускулатуры животное могло совершать длинные прыжки, затрачивая при этом минимум энергии.
Ноги травоядного, обитающего на равнине, весят очень мало и не обладают большой силой. Однако они идеально приспособлены для бегства от хищника и позволяют животному покрывать большие расстояния на высокой скорости, стоит ему только войти в надлежащий ритм бега.
Рис. 2.12.1.8. Дейнотерий (Deinotherium)На протяжении миоцена лошади продолжали увеличиваться в размерах. Меригиппус (рис. 2.12.1.6) по величине сравнялся с нынешним пони. Средний палец на каждой его ноге был непропорционально большим по сравнению с таким же пальцем у его предшественников, и можно сказать, что меригиппус всю свою жизнь ходил на цыпочках,— точнее, не ходил, а очень быстро бегал. Многочисленные бугорки на коренных зубах помогали ему пережевывать жесткую траву. Прежде лошади были лесными жителями и питались нежной и сочной листвой. Однако к началу миоцена они приспособились к жизни на открытых равнинах.
Теперь слоны стали куда больше похожи на их современных представителей. Мастодонт (рис. 2.12.1.7) — его называют платибелодон («лопатобивневый«) — прокладывал себе дорогу сквозь заросли подобно тяжелому бульдозеру. Из его нижней челюсти торчали широкие лопатообразные клыки, которыми он выкапывал из почвы разные растения. Дейнотерий (рис. 2.12.1.8) был намного крупнее (около 4 м в плече). Изогнутыми клыками нижней челюсти он, возможно, поддевал съедобные коренья, как большими вилами.
Рис. 2.12.1.9. Халикотерии (Chalicothenidae)Мы можем получить некоторое представление о миоценовой экосистеме, если обратимся к ее современному аналогу — восточноафриканским саваннам. Травянистая равнина обеспечивает разными видами корма разнообразных животных. В саваннах Восточной Африки зебры объедают грубые верхушки трав, а антилопы гну их поросшие листьями центральные части. Газели же отыскивают богатые протеином семена и побеги у самой земли. Бородавочники часто опускаются на колени, чтобы дотянуться до самой короткой травы или выкопать из земли съедобные луковицы и клубни. Есть в саваннах и травоядные, которые находят корм выше уровня самых высоких трав. К примеру, черный носорог питается древесной корой, тонкими ветками и листвой, а вот слон ест и траву, и листья деревьев, зачастую поглощая за день до 250 кг растительности. Ну а жирафу рост позволяет избегать какой бы то ни было конкуренции, поскольку он может обрывать веточки и листву на высоте 6 м от земли. Таким образом, различные виды растительноядных животных не претендуют на пищевые ресурсы друг друга, и корма здесь хватает на всех. Возможно, так же было и во времена миоцена: различные виды существовали за счет различных частей экосистемы.
Рис. 2.12.1.10. Эгиптопитек (Egiptopitek)В тот же период появились и другие новоселы. В начале миоцена ряды птиц пополнились новыми видами попугаев, пеликанов, голубей и дятлов. Чуть позже к ним присоединились первые вороны и соколы. Бурно эволюционировали такие новые млекопитающие, как мыши, крысы, морские свинки и дикобразы. Появилась и странная группа лошадеобразных животных — их называют халикотериями (рис. 2.12.1.9). Большими когтями, напоминающими копыта, они выкапывали съедобные коренья.
Отныне животные могли свободно перемещаться из Африки в Европу или Азию и обратно. Вскоре возникло своего рода двустороннее движение, при котором слоны мигрировали из Африки в Евразию и Северную Америку, а кошки, буйволы, жирафы и свиньи путешествовали в обратном направлении.
Первые приматы были маленькими зверьками, похожими на землероек. Они появились на Земле около 65 млн лет назад. Эволюция продолжила работу в этом направлении, и к середине олигоцена сформировались две основные группы приматов — обезьяны Нового Света (в Южной Америке) и обезьяны Старого Света (в Африке и Азии).
Рис. 2.12.1.11. Дриопитек (Dryopithecus)Вскоре от африканской ветви произошла еще одна группа обезьян, ставшая родоначальницей человекообразных обезьян и в конечном итоге — человека. Мозг человекообразных обезьян крупнее, чем у прочих их сородичей. Кроме того, у них нет хвоста, а длинные и сильные руки отлично приспособлены для лазанья по деревьям и перепрыгивания с ветки на ветку.
В руки ученых попали ископаемые останки небольшой человекообразной обезьяны, которую назвали эгиптопитеком («египетской обезьяной»). Она обитала в Африке в олигоценовую эпоху, около 27 млн лет назад (рис. 2.12.1.10).
Никто не может с уверенностью утверждать, что именно эгиптопитек был родоначальником современных человекообразных обезьян, но это вовсе не исключено.
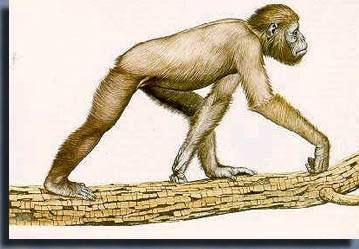
Вскоре после начала миоцена (около 24 млн лет назад) появилась и другая, более высокоразвитая человекообразная обезьяна — дриопитек (рис. 2.12.1.11), похожий на нынешних шимпанзе. Эти животные быстро перебрались со своей африканской родины, через сухопутные «мосты» в Европу и Азию. По всей видимости, дриопитек ходил на двух ногах, однако бегал и лазил по деревьям при помощи всех четырех конечностей. Возможно, он даже переносил в руках пищу. Итак, история человечества должна была вот-вот начаться. [3]
|>> Животный мир плиоцена>>
А.С.Антоненко
Источник: www.wwlife.ru
Общая информация
Геологические периоды Земли (таблица приведена ниже) представляют собой последовательность событий, которые произошли в процессе развития планеты с момента образования на ней земной коры. С течением времени на поверхности происходят различные процессы, такие как возникновение и разрушение форм рельефа, погружение участков суши под воду и поднятие их, оледенение, а также появление и исчезновение разных видов растений и животных и т. д. Наша планета носит на себе явные следы своего образования. Ученые утверждают, что способны с математической точностью их фиксировать в различных слоях пород.
Основные группы отложений
Геологи, пытаясь восстановить историю планеты, изучают пласты горных пород. Принято данные отложения делить на пять основных групп, выделяя следующие геологические эры Земли: древнейшую (архейскую), раннюю (протерозойскую), древнюю (палеозойскую), среднюю (мезозойскую) и новую (кайнозойскую). Считается, что граница между ними проходит по крупнейшим эволюционным явлениям, произошедшим на нашей планете. Последние три эры, в свою очередь, делятся на периоды, так как в данных отложениях наиболее ярко сохранились останки растений и животных. Каждому этапу свойственны события, которые оказали решающее влияние на сегодняшний рельеф Земли.
Древнейший этап
Архейская эра Земли отличалась довольно бурными вулканическими процессами, в результате которых на поверхности планеты оказались магматические гранитные породы – основа для формирования материковых плит. В то время здесь существовали только микроорганизмы, которые могли обходиться без кислорода. Предполагается, что отложения архейской эры покрывают практически цельным щитом отдельные площади материков, в них много железа, серебра, платины, золота и руд других металлов.
Ранний этап
Протерозойская эра также характерна высокой вулканической активностью. В этот период формировались горные массивы так называемой байкальской складчатости. До наших дней они практически не сохранились, сегодня представляют собой всего лишь отдельные несущественные поднятия на равнинах. В данный период Землю населяли простейшие микроорганизмы и сине-зеленые водоросли, появились первые многоклеточные. Протерозойский пласт горных пород богат полезными ископаемыми: слюдой, рудами цветных металлов и железными рудами.
Древний этап
Первый период палеозойской эры ознаменовался образованием горных хребтов каледонской складчатости. Это привело к существенному сокращению морских бассейнов, а также возникновению огромных участков суши. До наших дней сохранились отдельные хребты того периода: на Урале, в Аравии, Юго-Восточном Китае и Центральной Европе. Все эти горы «изношенные» и невысокие. Вторая половина палеозоя также характерна горообразовательными процессами. Здесь формировались хребты герцинской складчатости. Данная эпоха была более мощной, возникали обширные горные массивы на территориях Урала и Западной Сибири, Маньчжурии и Монголии, Центральной Европы, а также Австралии и Северной Америки. Сегодня они представлены весьма невысокими глыбовыми массивами. Животные палеозойской эры – это пресмыкающиеся и земноводные, моря и океаны заселены рыбами. Среди растительного мира преобладали водоросли. Палеозойская эра (каменноугольный период) характерна большими месторождениями каменного угля и нефти, которые возникли именно в эту эпоху.
Средний этап
Начало мезозойской эры характеризуется периодом относительного спокойствия и постепенного разрушения горных систем, созданных ранее, погружения под воду равнинных территорий (часть Западной Сибири). Вторая половина этого периода ознаменовалась образованием хребтов мезозойской складчатости. Появились весьма обширные гористые страны, которые и сегодня имеют такой же облик. В качестве примера можно привести горы Восточной Сибири, Кордильеры, отдельные участки Индокитая и Тибета. Земля была густо покрыта буйной растительностью, которая постепенная отмирала и перегнивала. Благодаря жаркому и влажному климату происходило активное образование торфяников и болот. Это была эпоха гигантских ящеров – динозавров. Обитатели мезозойской эры (травоядные и хищные животные) распространились по территории всей планеты. В это же время появляются и первые млекопитающие.
Новый этап
Кайнозойская эра, сменившая средний этап, длится по сей день. Начало данного периода ознаменовано возрастанием активности внутренних сил планеты, что привело к общему поднятию огромных участков суши. Эта эпоха характеризуется возникновением горных массивов альпийской складчатости в пределах Альпийско-Гималайского пояса. В этот период приобрел современные очертания Евразийский материк. Кроме того, произошло значительное омоложение древних массивов Урала, Тянь-Шаня, Аппалачей и Алтая. Резко поменялся климат на Земле, начались периоды мощных покровных оледенений. Движения ледниковых масс изменяли рельеф материков Северного полушария. В результате сформировались холмистые равнины с огромным количеством озер. Животные кайнозойской эры – это млекопитающие, пресмыкающиеся и земноводные, многие представители начальных периодов дошли и до наших дней, другие вымерли (мамонты, шерстистые носороги, саблезубые тигры, пещерные медведи и прочие) по тем или иным причинам.
Что такое геологический период?
Геологический этап как единицу геохронологической шкалы нашей планеты принято делить на периоды. Давайте посмотрим, что об этом термине сказано в энциклопедии. Период (геологический) – это крупный интервал геологического времени, при котором образовывались горные породы. В свою очередь, он подразделяется на меньшие единицы, которые принято называть эпохами.
Первые этапы (архейский и протерозойский) в связи с полным отсутствием или незначительным количеством в них отложений животного и растительного характера делить на дополнительные участки не принято. Палеозойская эра включает в себя кембрийский, ордовикский, силурский, девонский, карбонский и пермский периоды. Этот этап характеризуется наибольшим количеством подинтервалов, остальные ограничились всего тремя. Мезозойская эра включает в себя триасовый, юрский и меловой этапы. Кайнозойская эра, периоды которой наиболее изучены, представлена палеогеном, неогеном и четвертичным подинтервалом. Рассмотрим подробнее некоторые из них.
Триас
Триасовый период – это первый подинтервал мезозойской эры. Его продолжительность составила около 50 миллионов лет (начало — 251-199 миллионов лет назад). Для него характерно обновление морской и наземной фауны. Вместе с тем продолжают существовать немногочисленные представители палеозоя, такие как спириферид, табулят, некоторое пластинчатожаберные и др. Среди беспозвоночных весьма многочисленны аммониты, давшие множество новых форм, важных для стратиграфии. Среди кораллов преобладают шестилучевые формы, из брахиопод – теребратулиды и ринхонелиды, в группе иглокожих — морские ежи. Позвоночные животные в основном представлены рептилиями – крупными ящеротазовыми динозаврами. Широко распространены текодонты – сухопутные быстро передвигающиеся рептилии. Кроме того, в триасовый период появляются первые крупные обитатели водной среды – ихтиозавры и плезиозавры, однако своего расцвета они достигают только в юрском периоде. Также в это время возникли и первые млекопитающие, которые были представлены мелкими формами.
Флора в триасовый период (геологический) утрачивает элементы палеозоя и приобретает исключительно мезозойский состав. Здесь преобладают папоротниковые породы растений, саговообразные, хвойные и гинкговые. Климатические условия характеризуются значительным потеплением. Это приводит к высыханию многих внутренних морей, а в оставшихся существенно возрастает уровень солености. Кроме того, весьма сокращаются площади внутриконтинентальных водоемов, в результате чего развиваются пустынные ландшафты. Например, к этому периоду относят Таврическую формацию Крымского полуострова.
Юра
Юрский период получил свое название благодаря Юрским горам в Западной Европе. Он составляет среднюю часть мезозоя и наиболее плотно отражает главные особенности развития органики этой эры. В свою очередь, его принято разделять на три отдела: нижний, средний и верхний.
Животный мир этого периода представлен широко распространившимися беспозвоночными – головоногими моллюсками (аммонитами, представленными многочисленными видами и родами). Они по скульптуре и характеру раковин резко отличаются от представителей триаса. Кроме того, в юрский период происходит расцвет другой группы моллюсков – белемнитов. В это время значительного развития достигают шестилучевые рифообразующие кораллы, морские губки, лилии и ежи, а также многочисленные пластинчатожаберные. Зато полностью исчезают виды палеозойского брахиопода. Морская фауна позвоночных видов значительно отличается от триаса, она достигает огромного разнообразия. В юрский период широко развиты рыбы, а также водные рептилии – ихтиозавры и плезиозавры. В это время происходит переход с суши и приспосабливание к морской среде крокодилов и черепах. Огромного разнообразия достигают различные виды наземных позвоночных – рептилий. Среди них к своему расцвету приходят динозавры, которые представлены травоядными, хищными и другими формами. Большинство из них достигает 23 метров в длину, например, диплодок. В отложениях этого периода встречается новый вид рептилий – летающие ящеры, которые получили название «птеродактили». В это же время появляются и первые птицы. Флора юры достигает пышного расцвета: голосеменные растения, гинкговые, цикадовые, хвойные (араукарии), беннеттиты, саговниковые и, конечно же, папоротники, хвощи и плауны.
Неоген
Неогеновый период – это второй период кайнозойской эры. Начался он 25 миллионов лет назад и закончился 1,8 миллион лет назад. В это время произошли существенные изменения в составе фауны. Возникает большое разнообразие брюхоногих и двустворчатых моллюсков, кораллов, фораминиферов и кокколитофоридов. Широкое развитие получили земноводные, морские черепахи и костистые рыбы. В неогеновый период большого разнообразия достигают и наземные позвоночные формы. Например, появились быстропрогрессирующие гиппарионовые виды: гиппарионы, лошади, носороги, антилопы, верблюды, хоботовые, олени, бегемоты, жирафы, грызуны, саблезубые тигры, гиены, человекообразные обезьяны и прочие.
Под воздействием различных факторов в это время бурно эволюционирует органических мир: возникают лесостепи, тайга, горные и равнинные степи. В тропических областях – саванны и влажные леса. Климатические условия приближаются к современным.
Геология как наука
Геологические периоды Земли изучает наука – геология. Появилась она относительно недавно — в начале 20 века. Однако, несмотря на свою молодость, она смола пролить свет на многие спорные вопросы об образовании нашей планеты, а также происхождении населяющих ее существ. В этой науке мало гипотез, в основном используются только результаты наблюдений и факты. Спору нет, хранящиеся в земных слоях следы развития планеты в любом случае дадут более точную картину прошлого, чем любая сочиненная книга. Однако уметь прочитать эти факты и правильно их понять дано не всем, поэтому даже в этой точной науке время от времени могут возникать ошибочные интерпретации тех или иных событий. Там, где присутствуют следы огня, можно с уверенностью утверждать, что огонь был; а где есть следы воды, с такой же уверенностью можно утверждать, что вода была и так далее. И тем не менее ошибки тоже случаются. Чтобы не быть голословными, рассмотрим один такой пример.
«Морозные узоры на стеклах»
В 1973 году журнал «Знание – сила» опубликовал статью знаменитого биолога А. А. Любимцева «Морозные узоры на стеклах». В ней автор обращает внимание читателей на поразительное сходство ледяных узоров с растительными структурами. В качестве эксперимента он сфотографировал узор на стекле и показал фотографию знакомому ботанику. И тот без замедления опознал на снимке окаменелый след чертополоха. С позиции химии эти узоры возникают вследствие газофазной кристаллизации паров воды. Однако нечто подобное происходит и при получении пиролитического графита методом пиролиза метана, разбавленного водородом. Так, было установлено, что в стороне от этого потока образуются дендритные формы, которые весьма похожи на растительные остатки. Объясняется это тем, что существуют общие законы, которые управляют образованием форм в неорганической материи и живой природе.
В течение длительного времени геологи датировали каждый период геологический, опираясь на следы растительных и животных форм, найденных в залежах угля. И вот только несколько лет назад появились утверждения некоторых ученых о том, что такой способ неверный и что все найденные окаменелости — ни что иное, как побочный продукт формирования земных слоев. Спору нет, нельзя все мерить одинаково, однако необходимо более тщательно подходить к вопросам датировки.
А было ли всемирное оледенение?
Рассмотрим еще одно категоричное утверждение ученых, и не только геологов. Всем нам, начиная со школы, внушали о всемирном оледенении, которое накрыло нашу планету, в результате чего вымерли многие виды животных: мамонты, шерстистые носороги и многие другие. А современное подрастающее поколение воспитывается на квадрологии «Ледниковый период». Ученые в один голос утверждают, что геология — точная наука, не допускающая теорий, а использующая только проверенные факты. Однако это далеко не так. Здесь, как и во многих областях науки (истории, археологии и прочих), можно наблюдать закостенелость теорий и непоколебимость авторитетов. Например, еще с конца девятнадцатого века в кулуарах науки ведутся горячие споры о том, было ли оледенение или нет. В середине двадцатого века известным ученым-геологом И. Г. Пидопличко был опубликован четырехтомный труд «О ледниковом периоде». В этой работе автор поэтапно доказывает о несостоятельности версии о всемирном оледенении. Он опирается не на труды других ученых, а на лично им проведенные геологические раскопки (причем часть из них он проводил, будучи бойцом Красной Армии, участвуя в боях против немецких захватчиков) по всей территории Советского Союза и Западной Европы. Он доказывает, что ледник не мог покрывать весь континент, а носил только локальный характер, и что не он стал причиной вымирания многих видов животных, а совсем иные факторы – это и катастрофические события, приведшие к смещению полюсов («Сенсационная история Земли», А. Скляров); и хозяйственная деятельность самого человека.
Мистика, или Почему ученые не замечают очевидного
Несмотря на неопровержимые доказательства, приведенные Пидопличко, ученые не спешат отказываться от принятой версии оледенения. А дальше еще интереснее. Работы автора были изданы в начале 50-х годов, однако со смертью Сталина все экземпляры четырехтомника были изъяты из библиотек и университетов страны, сохранились только в запасниках библиотек, а получить их оттуда весьма не просто. В советское время все желающие взять эту книгу в библиотеке брались на учет спецслужбами. И даже сегодня существуют определенные проблемы в получении этого печатного издания. Однако благодаря интернету каждый желающий может ознакомиться с работами автора, который подробно разбирает периоды геологической истории планеты, объясняет происхождение тех или иных следов.
Геология — точная наука?
Считается, что геология — наука исключительно опытная, которая выводит заключения только из того, что видит. Если случай сомнительный, то она ничего не утверждает, высказывает мнение, допускающее обсуждения, и откладывает окончательное решение до тех пор, пока не будут получены однозначные наблюдения. Однако, как показывает практика, точные науки тоже ошибаются (например, физика или математика). И тем не менее ошибки не являются катастрофой, если их вовремя принимать и исправлять. Зачастую они не носят глобальный характер, а имеют локальное значение, нужно только иметь мужество, чтобы принять очевидное, сделать правильные выводы и двигаться дальше, на встречу новым открытиям. Современные же ученые показывают кардинально противоположное поведение, ведь большинство светил науки в свое время получили титулы, награды и признание за свою деятельность, и сегодня им совсем не хочется расставаться с ними. И такое поведение замечено не только в геологии, но и в других сферах деятельности. Только сильные люди не боятся признавать своих промахов, они радуются возможности развиваться дальше, ведь обнаружение ошибки — это не катастрофа, а наоборот, новая возможность.
Источник: FB.ru
Климат и география
Как и в предыдущий палеоген, в период неогена наблюдалась тенденция к глобальному охлаждению, особенно на более высоких широтах (известно, что сразу же после окончания неогена в эпоху плейстоцена, Земля подверглась серии ледниковых периодов, смешанных с более теплыми «межледниковыми эпохами»). Географически неоген был важен для наземных мостов, которые открывались между различными континентами: именно во время позднего неогена Северная и Южная Америка соединились центральноамериканским перешейком; Африка находилась в непосредственном контакте с южной Европой через сухой бассейн Средиземного моря; восточная Евразия и Западная Северная Америка присоединились к Сибири сухопутным мостам; медленное столкновение индийского субконтинента с Азией привело к образованию Гималайских гор.
Животный мир неогена
Млекопитающие
Глобальные климатические тенденции, в сочетании с распространением различных трав, сделали неогеновый период золотым веком открытых прерий и саванн.
Эти обширные пастбища стимулировали эволюцию парнокопытных и непарнокопытных животных, включая доисторических лошадей и верблюдов (которые произошли в Северной Америке), а также оленей, свиней и носорогов. Во время более позднего неогена взаимосвязи между Евразией, Африкой и Северной, и Южной Америкой создали почву для запутанной сети видов, что привело к почти полному исчезновению южноамериканской и австралийской мегафауны.
С точки зрения человека наиболее важным этапом неогенового периода была продолжающаяся эволюция обезьян и гоминид. В миоценовую эпоху огромное количество видов гоминидов обитало в Африке и Евразии; в течение последующего плиоцена большинство этих гоминид (в том числе прямых предков современных людей) были сгруппированы в Африке. Именно после неогенового периода, в эпоху плейстоцена, появились первые человеческие существа (род Homo) на планете.
Птицы
Некоторые из летающих и нелетающих видов птиц неогена были поистине огромными (например, аргентависы и остеодонторнисы превышали 20 кг). Конец неогена означал исчезновение большинства нелетающих хищных птиц из Южной Америки и Австралии. Эволюция птиц продолжалась быстрыми темпами, причем большинство современных видов хорошо представлены в конце неогена.
Рептилии
На протяжении большей части неогенового периода доминировали гигантские крокодилы, размеры которых не соответствовали с размерам их меловых предков.
Этот период в 20 млн лет также стал свидетелем продолжающейся эволюции доисторических змей и (особенно) доисторических черепах, последняя группа которых начала достигать действительно впечатляющих размеров к началу плейстоценовой эпохи.
Морская фауна
Хотя доисторические киты начали развиваться в предыдущий палеогеновый период, они не стали исключительно морскими существами до неогена, что также свидетельствовало о продолжающейся эволюции первых ластоногих (семейство млекопитающих, включая тюленей и моржей), а также доисторических дельфинов, с которыми киты тесно связаны. Доисторические акулы сохранили свой статус на вершине морской пищевой цепи; например, мегалодон уже появился в конце палеогена и продолжил свое доминирование на протяжении всего неогена.
Растительный мир неогена
В неогеновый период наблюдались две основные тенденции в растительной жизни. Во-первых, падение глобальных температур стимулировало рост массивных лиственных лесов, которые заменили джунгли и дождевые леса в высоких северных, и южных широтах. Во-вторых, всемирное распространение трав идет рука об руку с эволюцией травоядных млекопитающих, кульминацией которых являются сегодняшние лошади, коровы, овцы, олени и других пасущиеся и жвачные животные.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info
|
Подразделение неогеновой системы
Неогеновый период является временем максимального проявления альпийской эпохи складчатости. Наряду с образованием величайших горных складчатых сооружений в это время продолжается общее поднятие платформ. Морские трансгрессии в неогене уже не имеют большого размаха, и моря приобретают характер замкнутых, зачастую опресненных бассейнов, нередко постепенно превращающихся в озера и лагуны. Фауна в каждом из них приобретает своеобразные черты, что весьма усложняет сопоставление осадков и объясняет отсутствие единой стратиграфической схемы неогена.
В настоящее время для Юга СССР и Западной Европы принимаются следующие подразделения неогена ().
Подразделение неогеновой системы
Отдел Подотдел Ярус Примечание Верхнеплиоценовый N2 Апшеронский N2ap Акчагыльский N2ak Плиоценовый N2 Среднеплиоценовый 1| Куяльницкий N2kl Киммерийский N2k Нижнеплиоценовый N2 Понтический N2pn Часто пользуются местными подраз Верхнемиоценовый Nj Мэотический Njm Сарматский Nts делениями .(свиты, слои, реже серии) Миоценовый Мг Среднемиоценовый Nj Тортонский Njt Нижне миоценовый N J Гельветский Njh Бурдигальский Njb
Палеогеография, осадконакопление и структура земной коры
Тектонические движения, проявившиеся на протяжении палеогенового периода, заметно усложнили структуру земной коры, особенно в пределах геосинклинальных областей, но все же в начале неогенового периода продолжают существовать те же геосинклинальные области и платформы, что и раньше. В результате значительных общих поднятий и связанных с ними регрессий, происшедших к концу палеогенового периода, в начале нового, неогенового периода платформенные массивы — Евразийский и Северо-Американский в северном полушарии, и «обломки» некогда обширной Гондваны — Южно-Американ- ский, Африканский, Австралийский и другие материки стали почти повсеместно сушей. В это время район Зондского архипелага представлял собой сплошную сушу, соединенную с Евразийским платформенным массивом.
Полагают, что в неогеновом периоде существовала сухопутная связь между Северо-Американским и Евразийским платформенными массивами в районе Британских островов и Гренландии, а также межд Европой и Африкой в районе Средиземного моря. Средиземноморская геосинклинальная область в начале неогенового периода была занята морем островного характера: в виде островов среди него возвышались горы, возникшие в палеогеновом периоде. По обеим сторонам Тихого океана протягивались узкие геосинклинальные области — Восточно- Тихоокеанская и Западно-Тихоокеанская.
На протяжении неогенового периода в геосинклинальных областях с новой силой проявились альпийская складчатость и горообразование, придавшие альпийским горным сооружениям в основном современный облик; это высокие и самые молодые горы на земной поверхности, причем в этих молодых складчатых областях всегда горные хребты соответствуют положительным структурам — антиклинориям, а впадины и депрессии — отрицательным структурам, синклинориям.
Складкообразование достигло наибольшей силы в конце миоценовой и начале плиоценовой эпох. В конце плиоценовой эпохи оно заметно ослабело, но зато стали проявляться с большей силой дизъюнктивные (разрывные) дислокации, охватившие как области недавней, альпийской складчатости, так и участки палеозойских и мезозойских складчатых сооружений, к этому времени уже значительно или полностью денудированных.
В результате такие участки древних горных сооружений подверглись повторному поднятию и до настоящего времени они по своей высоте не уступают или даже превышают многие альпийские складчатые сооружения. К таким, вторично приподнятым по линиям разломов «глыбовым» горным сооружениям принадлежат каледонские структуры Скандинавии, каледонские и герцинские структуры Урала, Средней и Центральной Азии, Сибири (Тянь-Шань, Алтай Саяны и т. д.), Австралии, киммерийские структуры Северо-Востока СССР, Китая и Северной Америки ( 137). В пределах Гондваны продолжались расколы и формирование системы грабенов на Африканской платформе, сопровождавшиеся вулканическими извержениями.
Наиболее сложно происходило тектоническое развитие Средиземноморской геосинклинальной области. По своему развитию и современному тектоническому состоянию она существенно отличается от относящихся к альпийскому же циклу Западно-Тихоокеанской и Восточно- Тихоокеанской геосинклинальных областей. В пределах Средиземноморской области на протяжении неогенового периода имело место несколько фаз складчатости. В результате к концу периода под влиянием альпийской складчатости она почти закончила этап своего геосинклинального развития или, как говорят, пришла к стадии общей стабилизации, хотя некоторые черты геосинклинального развития сохранились в ее пределах и до сих пор (разрывные и складчатые дислокации, землетрясения, вулканические извержения).
Что же касается остальных геосинклинальных областей, расположенных по берегам Тихого окана, то, несмотря на образование в их пределах огромных по протяженности молодых альпийских горных сооружений, в них сохранились до сих пор все характерные черты геосинклинального развития, причем на довольно молодой своей стадии, что подтверждается рядом особенностей. Таковыми являются: сильно расчлененный рельеф поверхности земной коры (чередование глубоких и узких океанических впадин с высокоподнятыми островными дугами), резко дифференцированный характер тектонических движений, чрезвычайно высокая и в настоящее время сейсмичность (глубокофокусные землетрясения) и интенсивная вулканическая деятельность.
Альпийская складчатость в неогеновом периоде сопровождалась местами активным внедрением .и излиянием магмы .в воздымающихся складчатых структурах, особенно в Средиземноморской геосинклинальной зоне. С неогеновыми интрузиями связаны промышленные месторождения меди, свинца, цинка, молибдена, вольфрама и других цветных металлов. В частности, полиметаллические месторождения такого возраста имеются на Кавказе. В процессе формирования альпийских складчатых сооружений между возвышающимися дугами складчатых гор возникали обширные изолированные котловины, иногда значительной глубины. В качестве таких котловин рассматриваются современные Средиземное, Черное и другие моря, а также моря Зондского архипелага и Тихоокеанского побережья Азии. Наряду с такими котловинами, занятыми морями, образовались краевые (предгорные) и межгорные прогибы, заполнившиеся мощными толщами песчаных и глинистых осадков и являющиеся в настоящее время предгорными равнинами и низменностями (Предкарпатский, Предкавказский, Месопотамский и др.). Часть из них представляет собой занесенные осадками моря. В палеогеновых и особенно в неогеновых отложениях ряда таких прогибов обнаружены огромные скопления нефти. В неогеновое время не было сколько-нибудь значительных трансгрессий, и моря охватывали только краевые участки платформ, главным образом в районах, прилегающих к Средиземноморской геосинклинальной области. Так, временами южные окраины Русской платформы заливались водами морей, располагавшихся в основном в пределах соседней, Средиземноморской геосинклинальной области. В начале периода, в раннем и среднем миоцене, эти моря являлись еще прямым продолжением морей Средиземноморской области Европы, но с начала позднего миоцена новые поднятия и складкообразование в Альпах, на Карпатах, Балканах и в Закавказье привели к полному отделению бассейнов Юго-Восточной Европы. Возникло громадное замкнутое сарматское озеро-море, которое протягивалось от Венгрии на западе через Молдавию и юг Украины, вдоль Предкавказья до Аральского моря, охватывая и область современного Черного моря и Каспия. В конце миоценовой эпохи (мэотическом веке) это море несколько сократилось, но вплоть до начала плиоценовой эпохи (понтический век) оно оставалось единым.
После понтического века море юга Европы окончательно разделилось на ряд изолированных бассейнов — Средиземное, Черное, Каспийское моря и др., которые то сокращались до размеров меньше современных, то расширяли свои пределы. Так, ,в позднем плиоцене возникла обширная трансгрессия Каспийского (акчагыльского) моря, когда оно покрыло всю Прикаспийскую низменность и внедрилось узкими длинными заливами в уже существовавшие тогда долины рек Волги и Камы. Плиоценовые моря Черноморско-Каспийской области были временами сильно опреснены, и их осадки наряду с морской содержат и пресноводную фауну моллюсков.
В результате образования альпийских гор, значительного повторного поднятия многих областей древних денудированных складчатых сооружений, а также общего поднятия платформ рельеф земной поверхности в сравнении с предыдущими периодами приобрел совершенно новые черты, которые в основном сохранились по настоящее время.
Климатические особенности неогенового периода восстанавливаются довольно хорошо. В Западной Европе и на юге Русской платформы в начале периода существовал теплый умеренный или субтропический климат, но с зимним сезоном; это подтверждается составом растительных и животных остатков и другими признаками. К северу от этой зоны, вплоть до Гренландии, климат был умеренным. Во второй половине периода, в плиоценовую эпоху, климат в Европе стал на всем протяжении континентальным, умеренным (но все же более теплым, чем в современную эпоху). В конце неогенового периода наступило похолодание, и климатическая обстановка стала походить на современную. Некоторые ученые полагают даже, что климат в Западной Европе был более суровым, чем в настоящее время.
В связи с широким установлением континентального режима и воз- дыманием гор среди неогеновых отложений значительное развитие получили континентальные осадки и осадки внутренних морей и озер. В первой и начале второй половины периода в сравнительно узких эпи- континентальных бассейнах, располагавшихся вдоль геосинклинальных областей, отлагались ракушечные известняки, глины и пески. На окраинах и заливах таких бассейнов, расположенных в зоне теплого и сухого климата, отлагались соли и гипс (например, в Предкарпатье и Закарпатье, в Закаспийской области). На суше формировались во многих местах речные и озерные песчано-глинистые отложения, нередко угленосные, как в США, на Дальнем Востоке, в Западной Европе, на юге Русской платформы.
На тех участках геосинклинальных областей, где морской режим продолжал сохраняться на протяжении всего периода вдш большей его части, отлагались мощные толщи глин и песков с морской фауной. В краевых (предгорных) и межгорных прогибах и впадинах за счет разрушения уже существующих и еще воздымающихся горных сооружений накапливались очень мощные толщи обломочных пород типа мо- ласс; они отличаются очень обильной нефтеносностью и местами газоносностью. Такие нефтеносные толщи прослеживаются с небольшими перерывами как вдоль Средиземноморской, так и вдоль тихоокеанских геосинклинальных областей. К ним приурочены крупнейшие месторождения нефти на юге Западной Европы, Карпат, Кавказа, Ближнего и Среднего Востока (Иран, Ирак, Саудовская Аравия и др.), Бирмы, Малайского архипелага, Японии, Сахалина, Калифорнии, Центральной Америки (Мексика, Венесуэла, Колумбия и др.), Аргентина и т. д.
Нередко морские и озерные отложения неогена содержат мощные пласты оолитовых бурых железняков большого промышленного значения (Керченский полуостров, Тургайский прогиб, Западно-Сибирская низменность и др.).
Неогеновые отложения в СССР известны примерно в тех же местах, что и палеогеновые, но развиты они на значительно меньшей площади. На Русской платформе они представлены вдоль ее южных окраин. Они почти полностью отсутствуют на Сибирской платформе. Зато неогеновые толщи пород хорошо выражены на северных и восточных склонах Карпат и Горного Крыма, в Предкавказье и Закавказье, на обширных пространствах Средней Азии и Западно-Сибирской низменности. Хорошо представлен неоген на Камчатке, Курильских островах и Сахалине.
Органический мир
Состав неогеновой фауны и флоры во многом сходен с современным, но географическое распределение их было иное. В Южной Европе в начале неогена (в миоцене) продолжала существовать теплолюбивая флора, представленная как вечнозелеными растениями — пальмами, секвойями, болотными кипарисами, папоротниками, так и растениями с опадающей листвой—.дубами, кленами, тополями, буком, акацией и др. К северу распространялась растительность умеренного климата — лиственная и хвойная. В плиоцене площадь, занятая теплолюбивыми растениями, сильно сократилась; они продолжали существовать лишь на крайнем юге Европы. На остальной части Западной Европы, в Восточной Европе и на юге Сибири существовала флора теплого умеренного .пояса, а на севере росли хвойные леса. Таким образом, в неогене определились фитогеографические провинции, близкие современным, за исключением тундровой зоны.
В морской фауне неогена, как и в палеогене, широко представлены такие классы, как пелециподы и гастроподы; за ними по обилию форм следуют морские ежи, мшанки и др. Среди пелеципод и гастропод много современных родов (Cardium, Mactra, Venus, Tapes, Limnocardium, Con- geria, Spirialis и др.), но они выражены иными видами ( 138).
Большим разнообразием отличались млекопитающие. В неогеновом периоде возникли многие современные семейства и роды млекопитающих, но наряду с ними существовали семейства и роды, свойственные только этому периоду. Изучение неогеновых млекопитающих (и растений), которые сравнительно быстро распространялись на суше, показало, что некоторые ныне разобщенные материки время от времени получали сухопутную связь друг с другом и снова ее теряли. Такие связи устанавливаются, например, между Азией и Америкой через Берингов пролив и между Европой и Америкой через Исландию и Гренландию.
В неогеновом периоде широкое распространение получили семейства хищных, копытных и хоботных. В это время появились медведи, гиены, куницы, собаки, мастодонты, носороги, свиньи, быки, овцы, жирафы, человекообразные обезьяны, а в конце периода (плиоцен) — слоны, гиппопотамы, гиппарионы и настоящие лошади. Зоогеографи- ческие провинции неогенового периода были в общих чертах сходны с современными, но отличались очертаниями своих границ и размерами. В Австралии по-прежнему продолжалось развитие сумчатых.
В связи с резко выраженной изоляцией неогеновых бассейнов фауна образует комплексы, имеющие чисто местное значение. Среди руководящих ископаемых формы, имеющие широкое пространственное распространение, малохарактерны.
|
Источник: www.bibliotekar.ru