Схема строения клеточного ядра
Ядро клетки — важнейшая ее органелла, место хранения и воспроизведения наследственной информации. Это мембранная структура, занимающая 10-40 % клетки, функции которой очень важны для жизнедеятельности эукариотов. Однако даже без наличия ядра реализация наследственной информации возможна. Примером данного процесса является жизнедеятельность бактериальных клеток. Тем не менее особенности строения ядра и его предназначение очень важны для многоклеточного организма.
Расположение ядра в клетке и его структура
Ядро располагается в толще цитоплазмы и непосредственно контактирует с шероховатой и гладкой эндоплазматической сетью. Оно окружено двумя мембранами, между которыми находится перинуклеарное пространство. Внутри ядра присутствует матрикс, хроматин и некоторое количество ядрышек.
Некоторые зрелые человеческие клетки не имеют ядра, а другие функционируют в условиях сильного угнетения его деятельности. В общем виде строение ядра (схема) представлено как ядерная полость, ограниченная кариолеммой от клетки, содержащая хроматин и ядрышки, фиксированные в нуклеоплазме ядерным матриксом.
Строение кариолеммы
Для удобства изучения клетки ядра, последнее следует воспринимать как пузырьки, ограниченные оболочками от других пузырьков. Ядро — это пузырек с наследственной информацией, находящийся в толще клетки. От ее цитоплазмы он ограждается бислойной липидной оболочкой. Строение оболочки ядра похожее на клеточную мембрану. В действительности их отличает только название и количество слоев. Без всего этого они являются одинаковыми по строению и функциям.
Строение кариолеммы (ядерной мембраны) двуслойное: она состоит из двух липидных слоев. Наружный билипидный слой кариолеммы непосредственно контактирует с шероховатым ретикулумом эндоплазмы клетки. Внутренняя кариолемма — с содержимым ядра. Между наружной и внутренней кариомембраной существует перинуклеарное пространство. Видимо, оно образовалось из-за электростатических явления — отталкивания участков глицериновых остатков.
Функцией ядерной мембраны является создание механического барьера, разделяющего ядро и цитоплазму. Внутренняя мембрана ядра служит местом фиксации ядерного матрикса — цепи белковых молекул, которые поддерживают объемную структуру. В двух ядерных мембранах существуют специальные поры: через них в цитоплазму к рибосомам выходит информационная РНК. В самой толще ядра находятся несколько ядрышек и хроматин.
Внутреннее строение нуклеоплазмы
Особенности строения ядра позволяют сравнить его с самой клеткой. Внутри ядра также присутствует особая среда (нуклеоплазма), представленная гель-золем, коллоидным раствором белков. Внутри нее есть нуклеоскелет (матрикс), представленный фибриллярными белками. Основное отличие состоит только в том, что в ядре присутствуют преимущественно кислые белки. Видимо, такая реакция среды нужна для сохранения химических свойств нуклеиновых кислот и протекания биохимических реакций.
Ядрышко
Строение клеточного ядра не может быть завершенным без ядрышка. Им является спирализованная рибосомальная РНК, которая находится в стадии созревания. Позднее из нее получится рибосома — органелла, необходимая для белкового синтеза. В структуре ядрышка выделяют два компонента: фибриллярный и глобулярный. Они различаются только при электронной микроскопии и не имеют своих мембран.
Фибриллярный компонент находится в центре ядрышка. Он представляет собой нити РНК рибосомального типа, из которых будут собираться рибосомные субъединицы. Если рассматривать ядро (строение и функции), то очевидно, что из них впоследствии будет образован гранулярный компонент. Это те же созревающие рибосомальные субъединицы, которые находятся на более поздних стадиях своего развития. Из них вскоре образуются рибосомы. Они удаляются из нуклеоплазмы через ядерные поры кариолеммы и попадают на мембрану шероховатой эндоплазматической сети.
Хроматин и хромосомы
Строение и функции ядра клетки органично связаны: здесь присутствует только те структуры, которые нужны для хранения и воспроизведения наследственной информации. Также существует кариоскелет (матрикс ядра), функцией которого является поддержание формы органеллы. Однако самой важной составляющей ядра является хроматин. Это хромосомы, играющие роль картотек различных групп генов.
Хроматин представляет собой сложный белок, который состоит из полипетида четвертичной структуры, соединенного с нуклеиновой кислотой (РНК или ДНК). В плазмидах бактерий хроматин также присутствует. Почти четверть от всего веса хроматина составляют гистоны — белки, ответственные за «упаковку» наследственной информации. Эту особенность структуры изучает биохимия и биология. Строение ядра сложное как раз из-за хроматина и наличия процессов, чередующих его спирализацию и деспирализацию.
Наличие гистонов дает возможность уплотнять и укомплектовать нить ДНК в небольшом месте — в ядре клетки. Это происходит следующим образом: гистоны образуют нуклеосомы, которые представляю собой структуру наподобие бус. Н2В, Н3, Н2А и Н4 — это главные гистоновые белки. Нуклеосома образована четырьмя парами каждого из представленных гистонов. При этом гистон Н1 является линкерным: он связан с ДНК в месте е входа в нуклеосому. Упаковка ДНК происходит в результате «наматывания» линейной молекулы на 8 белков гистоновой структуры.
Строение ядра, схема которого представлена выше, предполагает наличие соленоидподобной структуры ДНК, укомплектованной на гистонах. Толщина данного конгломерата составляет порядка 30 нм. При этом структура может уплотняться и далее, чтобы занимать меньше места и менее подвергаться механическим повреждениям, неизбежно возникающим в процессе жизни клетки.
Фракции хроматина
Структура, строение и функции ядра клетки зациклены на том, чтобы поддерживать динамические процессы спирализации и деспирализации хроматина. Потому существует две главные его фракции: сильно спирализованная (гетерохроматин) и малоспирализованная (эухроматин). Они разделены как структурно, так и функционально. В гетерохроматине ДНК хорошо защищена от любых воздействий и не может транскрибироваться. Эухроматин защищен слабее, однако гены могут удваиваться для синтеза белка. Чаще всего участки гетерохроматина и эухроматина чередуются на протяжении длины всей хромосомы.
Хромосомы
Клеточное ядро, строение и функции которого описываются в данной публикации, содержит хромосомы. Это сложный и компактно упакованный хроматин, увидеть который можно при световой микроскопии. Однако это возможно только в случае, если на предметном стекле расположена клетка в стадии митотического или мейотического деления. Одним их этапов является спирализация хроматина с образованием хромосом. Их структура предельно проста: хромосома имеет теломеру и два плеча. У каждого многоклеточного организма одного вида одинаковое строение ядра. Таблица хромосомного набора у него также аналогичная.
Реализация функций ядра
Основные особенности строения ядра связаны с выполнением некоторых функций и необходимостью их контроля. Ядро играет роль хранилища наследственной информации, то есть это своего рода картотека с записанными последовательностями аминокислот всех белков, которые могут синтезироваться в клетке. Значит, для выполнения какой-либо функции клетка должна синтезировать белок, структура которого закодирована в гене.
Чтобы ядро «понимало», какой конкретно белок нужно синтезировать в нужный час, существует система наружных (мембранных) и внутренних рецепторов. Информация от них поступает к ядру посредством молекулярных передатчиков. Наиболее часто это реализуется посредством аденилатциклазного механизма. Так на клетку воздействуют гормоны (адреналин, норадреналин) и некоторые лекарства с гидрофильной структурой.
Вторым механизмом передачи информации является внутренний. Он свойственен липофильным молекулам — кортикостероидам. Это вещество проникает через билипидную мембрану клетки и направляется к ядру, где взаимодействует с его рецептором. В результате активации рецепторных комплексов, расположенных на клеточной мембране (аденилатциклазный механизм) или на кариолемме, запускается реакция активации определенного гена. Он реплицируется, на его основании строится информационная РНК. Позднее по структуре последней синтезируется белок, выполняющий некоторую функцию.
Ядро многоклеточных организмов
В многоклеточном организме особенности строения ядра такие же, как и в одноклеточном. Хотя существуют некоторые нюансы. Во-первых, многоклеточность подразумевает, что у ряда клеток будет выделена своя специфическая функция (или несколько). Это значит, что некоторые гены постоянно будут деспирализованы, тогда как другие находятся в неактивном состоянии.
К примеру, в клетках жировой ткани синтез белков будет идти малоактивно, а потому большая часть хроматина спирализована. А в клетках, к примеру, экзокринной части поджелудочной железы, процессы биосинтеза белка идут постоянно. Потому их хроматин деспирализован. На тех участках, гены которых реплицируются чаще всего. При этом важна ключевая особенность: хромосомный набор всех клеток одного организма одинаков. Только из-за дифференциации функций в тканях некоторые из них выключаются из работы, а другие деспирализуются чаще прочих.
Безъядерные клетки организма
Существуют клетки, особенности строения ядра которых могут не рассматриваться, потому как они в результате своей жизнедеятельности либо угнетают его функцию, либо вовсе избавляются от него. Простейший пример — эритроциты. Это кровяные клетки, ядро у которых присутствует только на ранних стадиях развития, когда синтезируется гемоглобин. Как только его количества достаточно для переноса кислорода, ядро удаляется из клетки, дабы облегчить ее не мешать транспорту кислорода.
В общем виде эритроцит представляет собой цитоплазматический мешок, наполненный гемоглобином. Похожая структура характерна и для жировых клеток. Строение клеточного ядра адипоцитов предельно упрощено, оно уменьшается и смещается к мембране, а процессы белкового синтеза максимально угнетаются. Эти клетки также напоминают «мешки», наполненные жиром, хотя, разумеется, разнообразие биохимических реакций в них чуть большее, чем в эритроцитах. Тромбоциты также не имеют ядра, однако их не стоит считать полноценными клетками. Это осколки клеток, необходимые для реализации процессов гемостаза.
Источник: FB.ru
Ядро есть в любой эукариотической клетке. Ядро может быть одно, или в клетке могут быть несколько ядер (в зависимости от ее активности и функции).
Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Ядерная оболочка состоит из двух мембран, разделенных перинуклеарным (околоядерным) пространством, между которыми находится жидкость. Основные функции ядерной оболочки: обособление генетического материала (хромосом) от цитоплазмы, а также регуляция двусторонних взаимоотношений между ядром и цитоплазмой.
Ядерная оболочка пронизана порами, которые имеют диаметр около 90 нм. Область поры (поровый комплекс) имеет сложное строение (это указывает на сложность механизма регуляции взаимоотношений между ядром и цитоплазмой). Количество пор зависит от функциональной активности клетки: чем она выше, тем больше пор (в незрелых клетках пор больше).
Основа ядерного сока (матрикса, нуклеоплазмы) – это белки. Сок образует внутреннюю среду ядра, играет важную роль в работе генетического материала клеток. Белки: нитчатые или фибриллярные (опорная функция), гетероядерные РНК (продукты первичной транскрипции генетической информации) и мРНК (результат процессинга).
Ядрышко – это структура, где происходят образование и созревание рибосомальных РНК (р-РНК). Гены р-РНК занимают определенные участки нескольких хромосом (у человека это 13–15 и 21–22 пары), где формируются ядрышковые организаторы, в области которых и образуются сами ядрышки. В метафазных хромосомах эти участки называются вторичными перетяжками и имеют вид сужений. Электронная микроскопия выявила нитчатый и зернистый компоненты ядрышек. Нитчатый (фибриллярный) – это комплекс белков и гигантских молекул-предшественниц р-РНК, которые дают в последующем более мелкие молекулы зрелых р-РНК. При созревании фибриллы превращаются в рибонуклеопротеиновые гранулы (зернистый компонент).
Хроматин получил свое название за способность хорошо прокрашиваться основными красителями; в виде глыбок он рассеян в нуклеоплазме ядра и является интерфазной формой существования хромосом.
Хроматин состоит в основном из нитей ДНК (40 % массы хромосомы) и белков (около 60 %), которые вместе образуют нуклеопротеидный комплекс. Выделяют гистоновые (пять классов) и негистоновые белки.
Гистонам (40 %) принадлежат регуляторная (прочно соединены с ДНК и препятствуют считыванию с нее информации) и структурная функции (организация пространственной структуры молекулы ДНК). Негистоновые белки (более 100 фракций, 20 % массы хромосомы): ферменты синтеза и процессинга РНК, репарации редупликации ДНК, структурная и регуляторная функции. Кроме этого, в составе хромосом обнаружены РНК, жиры, полисахариды, молекулы металлов.
В зависимости от состояния хроматина выделяют эухромати-новые и гетерохроматиновые участки хромосом. Эухроматин отличается меньшей плотностью, и с него можно производить считывание генетической информации. Гетерохроматин более компактен, и в его пределах информация не считывается. Выделяют конститутивный (структурный) и факультативный гетерохро-матин.
22. Микротрубочки представляют собой полые неветвящиеся цилиндры диаметром 20-25 нм, состоящие из мономеров белка тубулина. Микротрубочки участвуют в поддержании формы клетки, определяют ее полярность, разграничивают функционально отличающиеся друг от друга участки цитоплазмы, входят в состав клеточного центра, ресничек, а также веретена деления при митозе и мейозе.
Микрофибриллы и микрофиламенты — нитчатые структуры различной длины, но самые короткие и самые тонкие, толщиной от 5 до 10 нм, образованные белками (актин). Располагаются по всей цитоплазме, но особенно развиты у апикального полюса клетки, где образуют терминальную сеть. Встречаются в конусах роста, микроворсинках и на других участках клетки, находящихся в состоянии локомоторной активности, выполняют опорную и двигательную функции, в различных тканях образуя специальные органеллы.
Специальные органеллы — постоянные структуры, присущие клеткам определенных тканей. К ним относятся реснички, жгутики, тонофибриллы, нейрофибриллы, микроворсинки.
Реснички и жгутики — органеллы движения. Реснички развиты в клетках эпителия дыхательных путей и некоторых отделов половых трактов, а жгутики имеются у спермиев. Структура ресничек и жгутиков сходна, отличаются они лишь размерами. Реснички имеют длину 5-10 мкм, жгутики — 150 мкм. И те и другие представляют собой длинные тонкие выросты цитоплазмы, покрытые цитолеммой, имеющие в основании базальное тельце, а по всей длине — аксонему. Базальное тельце представляет собой центриоль, у которой можно наблюдать дополнительные структуры, характерные для материнской центриоли, а иногда можно видеть и дочернюю центриоль, расположенную под прямым углом к материнской. Основная нить ресничек и жгутиков — аксонема организована подобно центриоли, но содержит девять дублетов (пар) периферических и две центральные микротрубочки. Периферические и центральные микротрубочки связаны между собой в единую подвижную систему. При движении ресничек и жгутиков происходит скольжение микротрубочек друг по другу, в результате чего органеллы изгибаются.
Тонофибриллы, миофибриллы, нейрофибриллы — разновидности микрофибрилл и микрофиламентов, характерные для той или иной ткани. Тонофибриллы развиты в эпителиальных тканях, где образуют скелет клеток. Миофибриллы развиты в мышечных тканях, и определяют сократимость мышечных клеток и волокон, а нейрофибриллы — в нервных клетках. Предполагается, что они участвуют в токе аксоплазмы и проведении нервного импульса.
Микроворсинки — выросты цитоплазмы, одетые цитолеммой и содержащие внутри пучок микрофиламентов. Они увеличивают всасывательную поверхность клетки. Особенно хорошо развиты в кишечном эпителии.
Включения. Это необязательные компоненты клетки, появляющиеся и исчезающие в зависимости от интенсивности и характера обмена веществ в клетке и от условий существования организма. Включения имеют вид зерен, глыбок, капель, вакуолей, гранул различной величины и формы. Их химическая природа очень разнообразна. В зависимости от функционального назначения включения объединяют в группы: трофические, включения, секреты и инкреты, пигменты, экскреты и др. Среди трофических включений (запасных питательных веществ) важную роль играют жиры и углеводы. Белки как трофические включения используются лишь в редких случаях (в яйцеклетках в виде желточных зерен). Пигментные включения (желчные пигменты, мочевина и другие)
Источник: StudFiles.net
Особенности строения ядра
Заполнено ядро жидкостью и несколькими структурными элементами. В нем выделяют оболочку, набор хромосом, нуклеоплазму, ядрышка. Оболочка двухмембранная, между мембранами находится перенуклеарное пространство.

Внешняя мембрана сходна по строению с эндоплазматическим ретикулумом. Она связана с ЭПР, который будто ответвляется от ядерной оболочки. Снаружи на ядре находятся рибосомы.
Внутренняя мембрана прочная, так как в ее состав входит ламина. Она выполняет опорную функцию и служит местом крепления для хроматина.
Мембрана имеет поры, обеспечивающие обменные процессы с цитоплазмой. Ядерные поры состоят из транспортных белков, которые поставляют в кариоплазму вещества путем активного транспорта. Пассивно сквозь поровые отверстия могут пройти только небольшие молекулы. Также каждая пора прикрыта поросомой, которая регулирует обменные процессы в ядре.
Количество ядер в разных по специализации клетках различно. В большинстве случаев клетки одноядерные, но есть ткани, построенные из многоядерных клеток (печеночная или ткань мозга). Есть клетки лишенные ядра – это зрелые эритроциты.
У простейших выделяют два типа ядер: одни отвечают за сохранение информации, другие – за синтез белка.
Ядро может прибывать в состоянии покоя (период интерфазы) или деления. Переходя в интерфазу, имеет вид сферического образования с множеством гранул белого цвета (хроматина). Хроматин бывает двух видов: гетерохроматин и эухроматин.
Эухроматин – это активный хроматин, который сохраняет деспирализированное строение в покоящемся ядре, способен к интенсивному синтезу РНК.
Гетерохроматин – это участки хроматина, которые находятся в конденсированном состоянии. Он может при необходимости переходить в эухроматиновое состояние.
При использовании цитологического метода окрашивания ядра (по Романовскому-Гимзе) выявлено, что гетерохроматин меняет цвет, а эухроматин нет. Хроматин построен из нуклеопротеидных нитей, названных хромосомами. Хромосомы несут в себе основную генетическую информацию каждого человека. Хроматин — форма существования наследственной информации в интерфазном периоде клеточного цикла, во время деления он трансформируется в хромосомы.
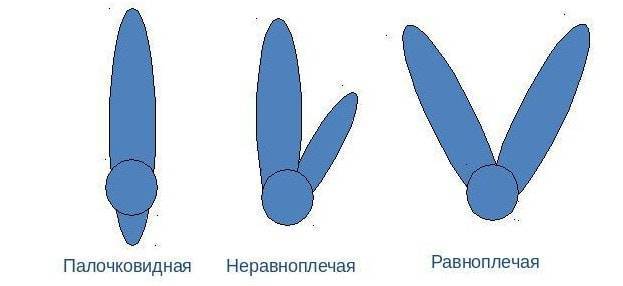
Строение хромосом
Каждая хромосома построена из пары хроматид, которые находятся параллельно друг к другу и связаны только в одном месте – центромере. Центромера разделяет хромосому на два плеча. В зависимости от длины плеч выделяют три вида хромосом:
- Равноплечие;
- разноплечие,
- одноплечие.
Некоторые хромосомы имеют дополнительный участок, который крепится к основному нитевидными соединениями – это сателлит. Сателлиты помогают идентифицировать разные пары хромосом.
Метафазное ядро представляет собой пластинку, где располагаются хромосомы. Именно в эту фазу митоза изучается количество и строение хромосом. Во время метафазы сестринские хромосомы двигаются в центр и распадаются на две хроматиды.
Строение ядрышка
В ядре также находится немембранное образование — ядрышко. Ядрышки представляют собой уплотненные, округлые тельца, способные преломлять свет. Это основное место синтеза рибосомальной РНК и необходимых белков.
Число ядрышек различно в разных клетках, они могут объединяться в одно крупное образование или существовать отдельно друг от друга в виде мелких частиц. При активации синтетических процессов объем ядрышка увеличивается. Оно лишено оболочки и находится в окружении конденсированного хроматина. В ядрышке также содержатся металлы, в большей мере цинк. Таким образом, ядрышко – это динамичное, меняющееся образование, необходимое для синтеза РНК и транспорта ее в цитоплазму.
Нуклеоплазма заполняет все внутреннее пространство ядра. В нуклеоплазме находится ДНК, РНК, протеиновые молекулы, ферментативные вещества.
Функции ядра в клетке
- Принимает участие в синтезе белка, рибосомной РНК.
- Регулирует функциональную активность клетки.
- Сохранение генетической информации, точная ее репликация и передача потомству.
Роль и значение ядра
Ядро является главным хранилищем наследственной информации и определяет фенотип организма. В ядре ДНК существует в неизмененном виде благодаря репарационным ядерным ферментам, которые способны ликвидировать поломки и мутации. Во время клеточного деления ядерные механизмы обеспечивают точное и равномерное расхождение генетической информации в дочерние клетки.
Источник: animals-world.ru
Тонкая структура клеточного ядра
Хроматин
Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы — структуры, на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой.
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначимых или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфорилированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
Ядрышко
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Ядерный матрикс
Ядерным матриксом некоторые исследователи называют нерастворимый внутриядерный каркас. Считается, что матрикс построен преимущественно из негистоновых белков, формирующих сложную разветвленную сеть, сообщающуюся с ядерной ламиной. Возможно, ядерный матрикс принимает участие в формировании функциональных доменов хроматина. В геноме клетки имеются специальные незначащие А-Т-богатые участки прикрепления к ядерному матриксу (англ. S/MAR — Matrix/Scaffold Attachment Regions), служащие, как предполагается, для заякоривания петель хроматина на белках ядерного матрикса. Впрочем, не все исследователи признают существование ядерного матрикса.
Эволюционное значение клеточного ядра
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (так как рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Модификациям подвергаются концы молекулы мРНК. К 5′ -концу молекулы прикрепляется 7-метилгуанин (так называемый кэп). К 3′-концу нематрично присоединяются несколько десятков остатков аденина (полиаденирование).
Процессинг мРНК тесно сопряжен с синтезом этих молекул и необходим для контроля качества. Непроцессированная или не полностью процессированная мРНК не сможет выйти из ядра в цитоплазму или будет нестабильна и быстро деградирует. У прокариот нет таких механизмов контроля качества, и из-за этого прокариотические мРНК имеют меньший срок жизни — нельзя допустить, чтобы неправильно синтезированная молекула мРНК, если такая появится, транслировалась в течение долгого времени.
Происхождение ядра
Клеточное ядро является важнейшей чертой эукариотических организмов, отличающей их от прокариот и архей. Несмотря на значительный прогресс в цитологии и молекулярной биологии, происхождение ядра не выяснено и является предметом научных споров. Выдвинуто 4 основных гипотезы происхождения клеточного ядра, но ни одна из них не получила широкой поддержки.[1]
Гипотеза, известная как «синтропная модель», предполагает что ядро возникло в результате симбиотических взаимоотношений между археей и бактерией (ни археи, ни бактерии не имеют оформленных клеточных ядер). По этой гипотезе, симбиоз возник, когда древняя архея (сходная с современными метаногенными археями), проникла в бактерию (сходную с современными Миксобактериями). Впоследствии архея редуцировалась до клеточного ядра современных эукариот. Эта гипотеза аналогична практически доказанным теориям происхождения митохондрий и хлоропластов, которые возникли в результате эндосимбиоза прото-эукариот и аэробных бактерий.[2] Доказательством гипотезы является наличие одинаковых генов у эукариот и архей, в частности генов гистонов. Также миксобактерии быстро передвигаются, могут образовывать многоклеточные структуры и имеют киназы и G-белки, близкие к эукариотическим.[3]
Согласно второй гипотезе, прото-эукариотическая клетка эволюционировала из бактерии без стадии эндосимбиоза. Доказательством модели является существование современных бактерий из отряда Planctomycetes, которые имеют ядерные структуры с примитивными порами и другие клеточные компартменты, ограниченные мембранами (ничего похожего у других прокариот не обнаружено).[4]
Согласно гипотезе вирусного эукариогенеза, окруженное мембраной ядро, как и другие эукариотические элементы, произошли вследствие инфекции прокариотической клетки вирусом. Это предположение основывается на наличии общих черт у эукариот и некоторых вирусов, а именно геноме из линейных цепей ДНК, кэпировании мРНК и тесном связывании генома с белками (гистоны эукариот принимаются аналогами вирусных ДНК-связывающих белков). По одной версии, ядро возникло при фагоцитировании (поглощении) клеткой большого ДНК-содержащего вируса.[5] По другой версии, эукариоты произошли от древних архей, инфицированных поксвирусами. Это гипотеза основана на сходстве ДНК-полимеразы современных поксвирусов и эукариот.[6][7] Также предполагается, что нерешенный вопрос о происхождении пола и полового размножения может быть связан с вирусным эукариогенезом.[8]
Наиболее новая гипотеза, названная экзомембранной гипотезой, утверждает, что ядро произошло от одиночной клетки, которая в процессе эволюции выработала вторую внешнюю клеточную мембрану; первичная клеточная мембрана после этого превратилась в ядерную мембрану, и в ней образовалась сложная система поровых структур (ядерных пор) для транспорта клеточных компонентов, синтезированных внутри ядра.[9]
Источник: dic.academic.ru