Экология фотосинтеза
Экология фотосинтеза
A.A. Пантявин, гимназия No 1, г. Воронеж
Со времени создания нашей биолого-химической гимназии в 1990 г. педагоги кафедры биологии работают над проблемой экологизации учебных программ.
Обучение по таким программам должно способствовать формированию личности, способной принимать экологически грамотные решения в области будущей профессиональной деятельности.
С этой целью применяются такие формы обучения, как проблемная лекция, семинар-дискуссия, практические занятия по моделированию, а также разнообразные формы лабораторных работ учащихся. Цикл занятий по теме «Фотосинтез» включал проблемную лекцию «Экология фотосинтеза», семинар-дискуссию «Фотосинтез», а также лабораторную работу, во время которой учащиеся определяли изменения морфологии листьев хвойных деревьев и содержания в них хлорофилла при воздействии токсикантов.
Цель занятия: формирование представлений о факторах среды, влияющих на процесс фотосинтеза.
Оборудование и материалы: книжка-раскладка «История фотосинтеза», таблица «Опыты Дж.Пристли», инструктивные и дидактические карточки, водное растение (элодея или роголистник), NаНСО3 или (NН4)2СО3, отстоявшаяся водопроводная вода, стеклянная палочка, нитки, ножницы, электролампы мощностью 60 и 250 Вт, часы, термометры, набор светофильтров, семь высоких широкогорлых банок, штативы с пробирками, термостат марки ТГУ 01-200.
ХОД ЗАНЯТИЯ
Занятие строится как бинарный урок, на котором проблемная лекция предшествует выполнению учащимися практических работ по группам.
В начале урока учитель объявляет цель занятия. Материал излагается в форме лекции, которая знакомит учащихся с историей открытия фотосинтеза и развитием учения об этом процессе на протяжении двух столетий. Отдельной задачей в теории фотосинтеза всегда было изучение влияния внешних факторов на скорость и эффективность этого процесса.
Лекция предусматривает элементы дискуссии.
Краткое изложение лекционного материала
Учитель излагает результаты опыта бельгийского естествоиспытателя ван Гельмонта (1580–1644), проведенного в начале XVII в. (работы ван Гельмонта были опубликованы лишь в 1648 г.). Ученый выращивал иву определенного веса в горшке с определенным количеством почвы при регулярном поливе растения дождевой водой. За пять лет масса растения увеличилась на 76, 7 кг, а масса почвы уменьшилась всего на 60 г.
Учитель обращается к ученикам с вопросом.
– Как вы думаете, какой вывод на основании проведенного опыта сделал ван Гельмонт?
Учащиеся делают предположение, что масса дерева увеличилась за счет воды, использованной для полива.
На доске записывают первое исходное вещество – H2O.
Затем преподаватель знакомит учащихся с опытами Джозефа Пристли (1733–1804), о котором гимназисты знают из курса неорганической химии как об одном из первооткрывателей кислорода. Свой рассказ учитель заканчивает вопросом.
– К какому выводу пришел Дж.Пристли, проведя этот эксперимент?
Учащиеся, как и ученый, предполагают, что растения выделяют кислород, делая воздух пригодным для дыхания.
Позже голландский врач Ян Ингенхауз (1730–1799) обнаружил, что растения выделяют кислород лишь на солнечном свету и только зелеными частями.
На доске появляется запись:
H2O + солнечный свет + зеленая часть растений — —> O2.
В 1782 г. швейцарский физиолог растений Жан Сенебье (1742–1809) экспериментально доказал участие в процессе фотосинтеза углекислоты воздуха, которая служит растениям «пищей». Вскоре после этого другой швейцарский исследователь, Никола Теодор де Соссюр (1767–1845), экспериментально доказал, что синтез органических соединений растениями обеспечивается питанием углекислотой (СO2) и H2O.
Эти открытия выдающихся ученых обобщаются учащимися в виде схемы на доске:
H2O + CO2 + солнечный свет + зеленая часть растений — —> O2 + органические вещества.
Раскрывая связь физиологического процесса с его физической основой, учитель обращает внимание учащихся на сделанное немецким врачом Р.Майером (1814–1878) в 1845 г. открытие, что зеленые растения преобразуют солнечную энергию в химическую энергию синтезируемых органических соединений согласно закону сохранения и превращения энергии.
Таким образом, уже к середине прошлого века о фотосинтезе сложились представления, которые можно выразить уравнением:
H2O + CO2 + солнечный свет + зеленая часть растений — —> O2 + органические вещества + химическая энергия.
Дальнейшие открытия связаны с именами французского физиолога растений Жана Батиста Буссенго (1802–1887), который точно измерил отношение поглощаемого при фотосинтезе углекислого газа к количеству выделяемого кислорода (1:1), и немецкого ботаника Юлиуса Сакса, продемонстрировавшего в 1864 г. образование зерен крахмала при фотосинтезе.
Учитель вновь обращается с вопросом к учащимся.
– Как вы думаете, каким образом Сакс показал, что органическим веществом, запасаемым зеленым растением при фотосинтезе, является крахмал?
Ученики, проводившие качественную реакцию на крахмал, отвечают на этот вопрос и уточняют, что с тех пор эта реакция называется пробой Сакса.
Таким образом, уже к началу XX в. были известны все компоненты фотосинтеза, как исходные, так и конечные:
Н2О + (СО2)n + солнечный свет + зеленая часть растений — —> (О2)n + крахмал + химическая энергия.
Итоговым результатом диалога учителя с учащимися является таблица «История фотосинтеза»
История фотосинтеза до XX в.
|
Дата |
Ученый |
Вклад в науку |
|
1600 |
Бельгийский естествоиспытатель ван Гельмонт |
Поставил первый физиологический эксперимент, связанный с изучением питания растений |
|
1771 |
Английский химик Джозеф Пристли |
Пришел к выводу о том, что зеленые растения способны осуществлять реакции, противоположные тем дыхательным процессам, которые были обнаружены в тканях животных |
|
1779 |
Голландский врач Ян Ингенхауз |
Обнаружил, что растения выделяют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода |
|
1782 |
Швейцарский физиолог растений Жан Сенебье |
Экспериментально доказал, что все соединения углерода в растениях образуются из углекислого газа, который под влиянием света разлагается в зеленых органах растений с выделением кислорода |
|
1804 |
Швейцарский агрохимик и физиолог растений де Соссюр |
Пришел к выводу, что вода также потребляется растением при ассимиляции двуокиси углерода |
|
1845 |
Немецкий физик и физиолог Роберт Майер |
Показал, что открытый им закон сохранения энергии справедлив для явлений живой природы |
|
1864 |
Французский физиолог растений Жан Буссенго |
Установил соотношение объемов превращающегося в органические соединения СО2 и выделяемого О2 (1:1) |
|
1864 |
Немецкий ботаник Юлиус Сакс |
Продемонстрировал образование зерен крахмала при фотосинтезе |
После диалога с учащимися учитель задает следующий вопрос.
– Какие факторы должны, судя по уравнению фотосинтеза, влиять на скорость этого процесса?
Учащиеся называют эти факторы (свет, температура, концентрация СО2 в воздухе, влажность и т.д.), а учитель записывает их на доске в порядке поступления предложений. Затем учитель организует с учащимися 40-минутную практическую работу для проверки высказанных ими предположений о влиянии света и температуры на скорость фотосинтеза. Для проведения практической работы класс делится на группы.
Первая группа получает следующую инструктивную карточку.
Экспериментальная работа №1
«Зависимость продуктивности фотосинтеза от интенсивности света»
Цель работы: изучение зависимости продуктивности фотосинтеза от интенсивности света.
Материалы и оборудование: элодея или роголистник; NаНСО3 или (NH4)2CO3; отстоявшаяся водопроводная вода; стеклянная палочка; нитки; ножницы; электролампа мощностью 200 Вт; часы; термометр.
ХОД РАБОТЫ
1. Выберите здоровые побеги элодеи или роголистника длиной около 8 см, интенсивного зеленого цвета с неповрежденной верхушкой, подрежьте их под водой, привяжите ниткой к стеклянной палочке и опустите верхушкой вниз в стакан с водой комнатной температуры (температура воды должна оставаться постоянной).
2. Для опыта взять отстоявшуюся водопроводную воду, обогащенную CO2 путем внесения NаНСО3 или (NН4)2СО3, и выставить стакан с водным растением на яркий свет. Наблюдайте за появлением пузырьков воздуха из среза растения.
3. Когда ток пузырьков станет равномерным, подсчитайте количество пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин, данные запишите в таблицу. Определите средний результат.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
|
№ п/п |
Интенсивность света |
Первое измерение |
Второе измерение |
Третье измерение |
Среднее значение |
|
|
|
|
|
|
|
4. Затем стакан с растением удалите от света на 50–60 см и повторите действия, указанные в п. 3.
5. Сравните результаты опытов и сделайте письменный вывод о различной интенсивности фотосинтеза на ярком и слабом свету.
В конце работы учащиеся первой группы отвечают на вопрос.
– Почему теневыносливые растения используют свет низкой интенсивности более эффективно, чем светолюбивые растения?
Учащиеся второй группы получают другое задание.
Экспериментальная работа №2
«Зависимость продуктивности фотосинтеза от спектрального состава света»
Цель работы: выяснение зависимости интенсивности фотосинтеза от спектрального состава света.
Материалы и оборудование: элодея или роголистник; набор светофильтров; семь высоких широкогорлых банок; отстоявшаяся водопроводная вода; ножницы; электролампа мощностью 200 Вт; часы; термометр; пробирки.
ХОД РАБОТЫ
1. Наполните пробирку на 2/3 объема отстоявшейся водопроводной водой и поместите в нее водное растение верхушкой вниз. Стебель подрежьте под водой.
2. Поместите в высокую широкогорлую банку синий светофильтр (круговой), поставьте в нее пробирку с растением и выставьте прибор на яркий свет так, чтобы он попадал на растение, проходя через светофильтр. Наблюдайте за появлением пузырьков воздуха из среза стебля растения.
3. Когда ток пузырьков станет равномерным, подсчитайте количестве пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин. Определите средний результат, данные запишите в таблицу.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
|
№ п/п |
Светофильтр |
Первое измерение |
Второе измерение |
Третье измерение |
Среднее значение |
|
|
|
|
|
|
|
4. Смените синий светофильтр на красный и повторите действия, указанные в п. 3. Следите за тем, чтобы расстояние от источника света и температура воды оставались постоянными.
5. Сравните результаты опытов и сделайте письменный вывод о зависимости интенсивности фотосинтеза от спектрального состава света.
После проведения работы учащиеся второй группы записывают ответ на вопрос.
– В каких лучах интенсивность фотосинтеза, судя по выделению кислорода, будет выше и почему?
Для учащихся третьей группы предложена следующая задача.
Экспериментальная работа №3
«Зависимость интенсивности фотосинтеза от температуры»
Цель работы: изучение зависимости интенсивности фотосинтеза от температуры.
Материалы и оборудование: элодея или роголистник; три высокие широкогорлые банки; отстоявшаяся водопроводная вода; ножницы; пробирки; электролампа мощностью 200 Вт; часы; термометр.
ХОД РАБОТЫ
1. Наполните пробирку на 2/3 объема отстоявшейся водопроводной водой и поместите в нее водное растение верхушкой вниз. Стебель подрежьте под водой.
2. Налейте в три высокие широкогорлые банки отстоявшуюся водопроводную воду разной температуры (+4 °С, +25 °С и +40 °С), поместите пробирку с растением в банку с водой с температурой +25 °С и выставьте прибор на яркий свет. Наблюдайте за появлением пузырьков воздуха из среза стебля растения.
2. Через 5 мин подсчитайте количество пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин. Определите средний результат, данные запишите в таблицу.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
|
№ п/п |
Температура |
Первое измерение |
Второе измерение |
Третье измерение |
Среднее значение |
|
|
|
|
|
|
|
4. Перенесите пробирку с растением в банку с водой другой температуры и повторите действия, указанные в п. 3. Следите за тем, чтобы расстояние от источника света и температура воды оставались постоянными.
5. Сравните результаты опытов и сделайте письменный вывод о влиянии температуры на интенсивность фотосинтеза.
После проведенных исследований учащиеся третьей группы должны ответить на вопрос.
– Почему при более высоких или более низких, чем 25 °С, температурах скорость фотосинтеза снижается?
По окончании практической работы учащиеся каждой группы докладывают результаты своих опытов, остальные учащиеся записывают их в своих тетрадях.
В конце занятия учащиеся делают итоговый вывод о влиянии различных факторов среды на интенсивность фотосинтеза.
Продолжением этой работы является семинар-дискуссия, на который выносятся следующие вопросы.
1. История фотосинтеза с начала XX в. до наших дней.
2. Влияние загрязнителей окружающей среды (сернистого газа, тяжелых металлов и др.) на фотосинтез у растений.
3. Космическая роль зеленых растений.
По этим вопросам учащиеся готовят сообщения и обсуждают их.
Результатом этой работы должно стать оформление продолжения таблицы «История фотосинтеза».
История фотосинтеза в XX в.
|
Год |
Ученый |
Вклад в науку |
|
1905 |
Английский физиолог растений Ф.Ф. Блэкмэн |
Высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. световую, реакцию и нефотохимическую, т.е. темновую, реакцию |
|
1932 |
Эмерсон и Арнольд |
Сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию |
|
1937 |
Р.Хилл |
Обнаружил, что изолированные хлоропласты способны высвобождать кислород в присутствии окислителя (акцептора электронов) |
|
1940 |
Г.Фишер |
Определил структурную формулу хлорофилла в опытах с последовательным разрушением молекул пигмента |
|
1941 |
Рубен и Камен |
Установили факт разложения воды, приводящего к выделению свободного кислорода при фотосинтезе |
|
1946 1953 |
Бассэм, Бенсон, Кальвин |
Определили последовательность темновых реакций фотосинтеза |
|
1954 |
Арнон, Аллен и Уотли |
Продемонстрировали процесс фотосинтеза в искусственных условиях вне клетки |
При обсуждении роли загрязнителей окружающей среды, влияющих на интенсивность фотосинтеза, учащиеся обсуждают в первую очередь роль сернистого газа (SO2) и серной кислоты (H2SO4), так как параллельно по неорганической химии они изучают тему «Серная кислота и ее производные».
Обсуждая космическую роль зеленых растений, учащиеся в итоге подводят «баланс» продуктивности фотосинтеза:
«1 см2 листа сахарной свеклы в течение дня образует из воздуха и воды с помощью солнечного света 0, 001 г сахара.
Ежегодно наземные растения связывают почти 20 млрд т, а растения морей – 25 млрд т углерода, которые расходуются на образование свыше 100 млрд т сахара. Для перевозки такого количества сахара потребовался бы железнодорожный состав длиной более чем в 50 млн км, что в 40 раз превышает общую длину всех железнодорожных линий Земли.
Для синтеза 1 кг глюкозы растениям достаточно затратить около 4, 4 кВт х ч электроэнергии, т.е. примерно столько же, сколько потребляет цветной телевизор за 15 ч.
При фотосинтезе растения используют за год почти 500 трлн кВт х ч солнечной энергии, что примерно в 200 раз превышает мировую выработку электроэнергии за год».
Определение загрязнения окружающей среды по изменениям комплекса признаков хвойных деревьев
Hayчно-исследовательская деятельность гимназистов при разработке этой темы заключается в определении морфологических и биохимических изменений у растений под действием антропогенных факторов.
Известно, что на загрязнение среды наиболее сильно реагируют хвойные древесные растения. Характерными признаками неблагополучия окружающей среды, и особенно газового состава атмосферы, является появление разного рода некрозов, уменьшение размеров многих органов (хвои, побегов текущего года и прошлых лет, их толщины, размера шишек, размера и числа заложившихся почек). Ввиду уменьшения роста побегов и хвои в длину у растений в загрязненной зоне наблюдается сближенность хвоинок (их число на 10 см побега больше, чем в чистой зоне).
Использование хвойных в качестве биоиндикаторов дает возможность оценить состояние окружающей среды. Для исследований предложен район памятника Славы, где растения оценивались как по качественным, так и по количественным показателям. Объектом исследований выбран вид ель обыкновенная. ветви дерева срезаны на высоте 2 м с части кроны, находящейся ближе к Задонскому шоссе (зоне с загрязненным воздухом). Контролем могут служить ветви деревьев, собранные в зеленой зоне города (Центральный парк культуры и отдыха).
При осмотре хвои с помощью лупы выявлено, что кончики большинства хвоинок имеют желто-коричневый цвет, что свидетельствует о загрязнении воздуха. Другие качественные признаки (колючесть, ломкость, смолистость) не позволили однозначно оценить состояние окружающей среды в исследуемом районе.
Длина хвоинок на побегах прошлого года у растений опытной группы в большинстве случаев была меньше таковой для контрольных образцов, однако количество листьев ели на 10 см побега исследуемой зоны не отличались от аналогичных показателей, характерных для чистой зоны. Толщина хвоинок варьировала от 1 до 5 мм. Количество сформировавшихся почек для опытных образцов равнялось 4±2, для контрольных 8±2.
На основании проведенных наблюдений и измерений можно сделать вывод о средней степени загрязнения воздуха в районе памятника Славы.
Определение содержания хлорофилла фотометрическим методом. Биоиндикация состояния окружающей среды
Биоиндикация – это метод оценки действия экологических факторов при помощи биологических систем. Например, воздействие SO2 на хвойные породы выражено в угнетении их роста и некрозе хвои. Содержание хлорофилла в листьях может также служить неспецифическим биоиндикационным признаком, т.к. снижение содержания хлорофилла наблюдается до появления видимых изменений листьев.
В качестве биоиндикатора выбран чувствительный к выхлопным газам автомобилей вид сосны обыкновенной, листья которой были собраны вблизи окружной автодороги.
Метод основан на регистрации оптических характеристик ацетоновой вытяжки хлорофилла, полученных с использованием фотоэлектроколориметра КФК-2. Для определения содержания хлорофилла в листьях использовали метод построения калибровочной кривой с использованием стандартного раствора Гетри.
Массовое соотношение взятого для исследований материала (листьев) и экстрагирующей жидкости (ацетона) равно 1:100.
В результате проведенных экспериментов показано, что в спектре поглощения молекул хлорофилла комнатного растения, выбранного в качестве контроля, имеются максимумы в синей (440 нм) и красной (670 нм) областях. Для ацетоновой вытяжки листьев сосны наблюдается смещение коротковолнового максимума поглощения в фиолетовую область (400 нм). Величины максимумов зависят от количества хлорофилла в вытяжке, которое рассчитывают на основании этих измерений.
Установлено, что содержание хлорофилла в контрольном и опытном образцах составило величину 0, 14±0, 02 мг/г листа, что соответствует нормальному количеству зеленого пигмента в листьях сосны.
Таким образом, не выявлено значительных изменений в содержании хлорофилла в контрольном и опытном образцах. Это свидетельствует о том, что содержание диоксида серы в воздухе и в выпадающих осадках (дожде, снеге) находится в пределах нормы.
Источник: znakka4estva.ru
Точка пересечения световой кривой с осью ординат находится ниже нулевой отметки по оси абсцисс. Пока интенсивность света
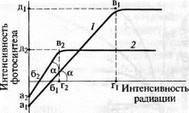
Рис. 6.15. Световые кривые фотосинтеза для светолюбивых (/) и теневыносливых растений (2) (по Т. К. Горышиной, 1979):
аи а2 — ниже оси абсцисс: растения в темноте, фотосинтез отсутствует, идет только дыхание; бь б2 — «компенсационные точки» (фотосинтез уравновешивает дыхание); в(, в2 — перегиб светой кривой фотосинтеза, она выходит на плато насыщения; Г|, проекция точек в(, в2 на ось абсцисс, характеризует «насыщающую» интенсивность света, выше которой свет не повышает интенсивность фотосинтеза; д,, д2 — проекция точек в,, в2 на ось ординат — наибольшая интенсивность фотосинтеза для данного вида в данном местообитании; а — углы наклона световой кривой к абсциссе, отражающей степень увеличения фотосинтеза при возрастании радиации (в области сравнительно низкой интенсивности света)
мала, идет выделение С02, а не поглощение его, так как фотосинтез не компенсирует дыхание. При некотором значении освещенности световая кривая пересекает линию абсцисс. Это значит, что количество поглощенного в фотосинтезе диоксида углерода равно количеству С02, выделенному при дыхании. За этой точкой уже преобладает фотосинтез, т.е. с повышением освещенности потребление С02 будет превышать его выход при дыхании. Минимальное освещение (при нормальном содержании С02), при котором интенсивность фотосинтеза (поглощения С02) уравновешивает дыхание (выделение С02), называют точкой компенсации (или компенсационным освещением). При такой освещенности для фотосинтеза используется столько С02, сколько высвобождается при дыхании. Поэтому интенсивно дышащие растения нуждаются в большем количестве света. Так, теневыносливые виды характеризуются более низкой интенсивностью дыхания, что позволяет им расти при меньшей освещенности. Светолюбивые растения лучше используют большую освещенность и в этих условиях имеют гораздо более высокую продуктивность фотосинтеза. Поэтому сельскохозяйственные растения, от которых требуется высокая урожайность, должны быть светолюбивыми (рис. 6.16).
Компенсационная точка варьирует даже у разных листьев одной кроны. Выросшие в тени дышат слабее световых, поэтому компенсация у них наступает при меньшем освещении. Теневые листья лучше используют слабый свет, и насыщение у них наступает очень рано (примерно при 10 клк). Растения или органы с низкой точкой компенсации способ-
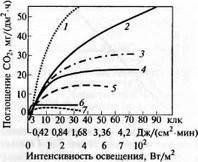
Рис. 6.16. Влияние света на нетто-фотосинтез различных растении при оптимальной температуре и естественном содержании С02 в воздухе
(по В.Лархеру, 1978):
1 — сорго; 2 — кукуруза; 3 — пшеница; 4 — светолюбивые травы; 5 — бук; 6 — теневыносливые травы; 7 — тенелюбивые мхи, планктонные водоросли
ны к более полному использованию света, но при освещенности ниже критической они погибают. Так происходит отмирание нижних листьев и очищение ствола от ветвей.
Точка компенсации суммирует действие разных факторов на дыхание и фотосинтез. Знание ее необходимо при изучении продуктивности растений, поскольку она указывает на границу между запасанием и расходованием органики. Ниже нее наступает голодание. Компенсационная точка обычно определяется при концентрации С02 0,03 % и температуре 20 °С. При постоянной концентрации С02 она повышается с ростом температуры, так как при потеплении дыхание увеличивается сильнее фотосинтеза. Поэтому при пониженной освещенности (например, зимой в оранжереях) необходима умеренная положительная температура и повышение ее может снизить темпы роста растений. Улучшение водного режима и снабжения С02 сдвигает компенсационную точку в сторону меньшей освещенности, а старение листьев — в сторону большей.
Угол наклона восходящей части кривой к оси абсцисс показывает эффективность использования света растением (свидетельствует о скорости увеличения интенсивности фотосинтеза по мере возрастания освещенности). В этой области фотосинтез определяется скоростью световых реакций: чем выше квантовый выход фотосинтеза, тем круче поднимается световая кривая. Это важный показатель функционального состояния растений.
Перегиб световой кривой указывает на два важных параметра. Во-первых, минимальный уровень освещения, обеспечивающий в данных условиях максимальную интенсивность фотосинтеза. Дальнейший рост освещенности не увеличивает его, поэтому световая кривая выходит на плато и происходит световое насыщение фотосинтеза. Во-вторых, уровень плато показывает максимальную величину фотосинтеза, которой достигает растение в данных условиях. Он определяет продуктивность растения при достаточном освещении, однако в разных условиях этот показатель сильно колеблется.
У большинства растений световые кривые постепенно выходят на плато, но светолюбивые виды при повышении концентрации С02 и оптимальном сочетании других факторов плато не имеют, образуя идущие вверх линии, лишь несколько отгибающиеся. У теневыносливых видов плато выражено хорошо, и чем меньше интенсивность света, при которой наступает перегиб световой кривой, тем более теневынослив вид. С повышением концентрации С02 уровень плато теневыносливых видов может повышаться, а перегиб смещаться в сторону повышенной освещенности. При сравнении световых кривых особо выделяются С4-виды. У них световое насыщение не достигается, и в ясные дни они используют свет полностью даже в полдень. И при средней освещенности эти растения фотосинтезируют интенсивнее С3-видов. Цикл Кальвина у С3-растений работает не так эффективно, поэтому их световая кривая делает изгиб раньше. Среди них есть и такие растения, фотосинтез которых при излишней освещенности снижается, т.е. для них существует оптимум освещения. Это споровые сеянцы древесных пород и травы сомкнутых лесов.
Световая кривая планктонных водорослей имеет колоколовидную форму: после достижения оптимума она падает (область перенасыщения), так как интенсивность света больше насыщающей подавляет их

Рис. 6.17. Зависимость между интенсивностью света и фотосинтезом (по Р. Риклефсу, 1979):
/ — зеленые водоросли; 2 — диатомовые водоросли; 3 — белый дуб; 4 — ладанная сосна; 5 — кизил
фотосинтетическую активность (рис. 6.17). Поэтому наибольшая интенсивность фотосинтеза при высоком стоянии солнца отмечается не у самой поверхности, а несколько глубже (в озерах на глубине — 2—5 м, в открытом море — 10—15 м) в зависимости от силы света над водой и ее мутности. Вблизи поверхности воды свет избыточен, и здесь, особенно в солнечные дни, может происходить ингибирование фотосинтеза. Вероятно, это обусловлено в основном тем, что скорость поглощения радиации пигментами так высока, что ее энергия не может быть использована водорослями нормальным путем и расходуется на деструктивные фото- окислительные реакции. В пасмурные же дни и в бедное светом время года торможения фотосинтеза светом в водоемах нет и область оптимума смещается до самой поверхности.
Параметры световых кривых специфичны для разных по отношению к свету экологических групп растений, они значительно меняются в зависимости от действия других экологических факторов, стадии развития растения, площади и расположения листьев. Таким образом, световые кривые несут много важной экологической информации.
Источник: myzooplanet.ru
На примере растений разных экологических групп, жизненных форм (деревьев, кустарников, трав и т. д.) и условий обитания (тропики, умеренный пояс, Арктика, пустыня) накоплено достаточно материала, характеризующего особенности фотосинтеза (О. В. Заленский, 1977; К. Я. Биль, 1993).
В пособиях по физиологии растений указанной теме уделяется достаточно внимания, что делает излишним ее широкое обсуждение. Так, разделение растений на светолюбивые и теневыносливые основано на их способности адаптироваться к разным световым условиям. Фотосинтетический электронный транспорту светолюбивых растений интенсивнее, чем у теневыносливых.
Светолюбивые растения больше вытягиваются при недостатке света, чем теневыносливые. В. Н. Любименко более чем на 600 видах показал, что листья у вторых по сравнению с первыми содержат повышенное количество хлорофилла. Даже одни и те же растения, хорошо освещенные, содержат 1,5—3 мг хлорофилла на 1 г листовой пластинки, тогда как растущие в тени — 4—6 мг/г. У теневых растений и концентрация хлорофилла на один хлоропласт больше и сами хлоропласта крупнее. Однако интенсивность фотосинтеза не связана с количеством хлорофилла. У освещенных листьев хлорофилл a преобладает над хлорофиллом b, а у затененных соотношение a и b падает. Состав пигментов меняется и у других растений. Так, по мере увеличения глубины водоема у водорослей происходит увеличение содержания бурых пигментов (фукоксантин), у наиболее же глубоководных (красные водоросли) —красных пигментов (фикоэретрин). У красных водорослей отношение фикоэретрин/хлорофилл оказывается тем больше, чем глубже они обитают в водоеме. Следовательно, явление «хроматической адаптации» — общее для зеленых растений.
Многими исследованиями показано, что растения разных экологических групп содержат неодинаковое количество хлорофилла. Очень часто содержание хлорофилла определяет энергетический выход у листьев — накопление энергии в единицу времени (О. Хитт, 1973). В то же время разные виды растений по числу и размерам хлоропластов в 1 мм2 (А. Т. Мокроносов, 1981; и др.) отличаются, что часто коррелируется с образованием сухого вещества и содержанием хлоропластов. Так, у Tropaeolum major число хлоропластов на 1 мм2 составляет 383 тыс., а сухое вещество (г/м2 листовой поверхности за 10 ч) — 4,47; у Phaseolus multiflorous —283 тыс. и соответственно 3,22; у Ricinus communis — 495 тыс. и 5,29; у Helianthus annuus — 465 тыс. и 5,57. Число хлоропластов на 1 см2 листа варьирует от 2 до 90 млн. Много пластид в листьях ксерофитов аридных пустынь, обитающих в условиях высокой интенсивности света: у астрагала хивинского — 89 млн., у осоки вздутой — 72 млн.
В связи с экологией фотосинтеза растений представляют интерес данные по оценке стока углерода как показателя годичной продуктивности, выраженного величиной нетто-фотосинтеза с учетом затрат на дыхание (П. Ю. Вороник и др., 1995). Для этого определяют прежде всего проективное содержание хлорофилла — хлорофилльный индекс. Обнаружена высокая корреляция (r = 0,72) между годичным фотосинтетическим стоком углерода и проективным содержанием хлорофилла в растительном покрове — фитоценозах. Так, продуктивность растительности сухопутной России оценивается в 1,6 Гга, тогда как планеты —50 Гга в год. Именно наличие корреляции между проективным содержанием хлорофилла того или иного типа фитоценоза и годичной его продуктивностью фотосинтеза позволяет подойти к оценке приспособленности фитоценозов к климатическим факторам и судить об их возможных сукцессионных последствиях характерно, что биоразнообразие мало влияет на конечный результат расчета, это облегчает задачу оценки продуктивности фитоценоза с учетом содержания хлорофилла у 2—3 доминирующих видов в проективном покрытии его.
Экологические особенности видов, их приспособленность к среде отражаются и на продуктах фотосинтеза. В ходе эволюции фототрофности конечные продукты становятся все более однообразными («специализация» веществ). Так, у бактерий и водорослей они более разнообразны, чем у высших растений. При сравнении фотосинтеза лишайников и высших растений показано, что при одинаковом наборе ассимилятов метаболизм лишайников отличается некоторыми специфическими особенностями: 1) увеличением в продуктах фотосинтеза доли маннита и, сахарозы; 2) высокой фотосинтетической термостабильностью; 3) восстановлением CO2 до сахароспиртов даже при температуре от —15 до —20° С.
Мир высших растений характеризуется исключительным морфофизиологическим разнообразием, что отражается и на организацию донорно-акцепторных отношений (А. Т. Мокроносов, 1981), имеющих приспособительное значение. Приспособительные изменения фотосинтеза можно обнаружить и при анализе положения Координационных точек нетто-фотосинтеза при световом насыщении, границ жизни и фотосинтеза у разных экологических групп растений; Оптимум фотосинтеза определяется температурными границами жизни вида и популяций как результат их приспособленности.
В пределах каждой группы растений наблюдается еще световая адаптация. Так, водоросли в целом относятся к теневыносливым растениям, у которых насыщение фотосинтеза происходит при низких интенсивностях света. Например, у сифоновых водорослей, обитающих в скелетах коралловых рифов, насыщение фотосинтеза достигается при освещении 10 лк, а у хлореллы и некоторых синезеленых — при 0,2 лк. У одних и тех же организмов световое насыщение фотосинтеза меняется по сезонам года: у динофлагеллят оно происходит летом при 34,7, весной и осенью — 2,5, зимой — 1 тыс. лк. Световая адаптация фотосинтеза связана со многими событиями: изменением концентрации пигмента, активностью ферментных систем, положением и числом хлоропластов, инактивацией молекул хлорофилла. Некоторые водоросли способны синтезировать хлорофилл даже в темноте, у некоторых в темноте происходит его разрушение. Так, Scenedesmus acuta в течение 8 лет в темноте сохранила зеленый цвет и вскоре после выставления на свет стала фиксировать CO2. У некоторых водорослей адаптация к световым условиям бывает растянута во времени от 2—3 сут (Cyelotella) до 2 недель (Oscillatoria rubens). В зависимости от времени года меняется даже тип фотосинтеза и содержание хлорофилла в листьях. На примере однолетних растений из семейства марьевых, произрастающих в условиях недостатка влаги показано, что листья весенней генерации осуществляют фотосинтез С4-типа, а летней генерации — по смешанному С4— САМ-типу (В. Ю. Любимов, К. Я. Биль, 1986). В фотосинтетической адаптации растений важное значение имеет число и распределение пластид в палисадной паренхиме. Оказалось, что анатомический тип листа больше влияет на распределение, величину и структурные вариации пластид, чем экологические особенности растений (Ю. В. Гамалей, Т. Р. Милашвилли, 1986).
В прошлом теплый климат протерозоя и палеозоя и высокая влажность воздуха, высокое содержание CO2 и низкое содержание O2 более благоприятствовали фотосинтезу, чем нынешние условия (А. А. Ничипорович, 1973). Количество углерода, доступное для фототрофов, ныне сократилось примерно в 800 раз по сравнению с периодом возникновения жизни (В. В. Межерин, 1976).
В указанные эры масштабы фотосинтеза превосходили скорость биологического окисления органического вещества, что способствовало процветанию жизни. Одним из сдерживающих факторов фотосинтеза ныне стало обеднение атмосферы CO2.
Стабилизацией физико-химического механизма не завершается эволюция фотосинтеза, а продолжается его биологический этап в направлении поиска экологических новшеств для интенсивного использования самого процесса. Так, с возникновением морфологического разнообразия растений в ходе эволюции параллельно происходит экологическая их дивергенция по интенсивности фотосинтеза. Переход от одноклеточных к многоклеточным и особенно к дифференцированным водорослям, а также смена структурной организации жизненных форм и тип растительности, образование многокомпонентных биогеоценозов представляют собой ступени на пути интенсификации фотосинтеза как планетарного явления. Подобным образом постоянно усиливается разнообразие адаптивных возможностей фотосинтеза разных объектов и сообществ, что является проявлением экологической, эволюции растений.
Источник: www.activestudy.info
A. A. Пантявин, гимназия No 1, г. Воронеж
Со времени создания нашей биолого-химической гимназии в 1990 г. педагоги кафедры биологии работают над проблемой экологизации учебных программ.
Обучение по таким программам должно способствовать формированию личности, способной принимать экологически грамотные решения в области будущей профессиональной деятельности.
С этой целью применяются такие формы обучения, как проблемная лекция, семинар-дискуссия, практические занятия по моделированию, а также разнообразные формы лабораторных работ учащихся. Цикл занятий по теме “Фотосинтез” включал проблемную лекцию “Экология фотосинтеза”, семинар-дискуссию “Фотосинтез”, а также лабораторную работу, во время которой учащиеся определяли изменения морфологии листьев хвойных деревьев и содержания в них хлорофилла при воздействии токсикантов.
Цель занятия: формирование представлений о факторах среды, влияющих на процесс фотосинтеза.
Оборудование и материалы: книжка-раскладка “История фотосинтеза”, таблица “Опыты Дж. Пристли”, инструктивные и дидактические карточки, водное растение (элодея или роголистник), NаНСО3 или (NН4)2СО3, отстоявшаяся водопроводная вода, стеклянная палочка, нитки, ножницы, электролампы мощностью 60 и 250 Вт, часы, термометры, набор светофильтров, семь высоких широкогорлых банок, штативы с пробирками, термостат марки ТГУ 01-200.
ХОД ЗАНЯТИЯ
Занятие строится как бинарный урок, на котором проблемная лекция предшествует выполнению учащимися практических работ по группам.
В начале урока учитель объявляет цель занятия. Материал излагается в форме лекции, которая знакомит учащихся с историей открытия фотосинтеза и развитием учения об этом процессе на протяжении двух столетий. Отдельной задачей в теории фотосинтеза всегда было изучение влияния внешних факторов на скорость и эффективность этого процесса.
Лекция предусматривает элементы дискуссии.
Краткое изложение лекционного материала
Учитель излагает результаты опыта бельгийского естествоиспытателя ван Гельмонта (1580-1644), проведенного в начале XVII в. (работы ван Гельмонта были опубликованы лишь в 1648 г.). Ученый выращивал иву определенного веса в горшке с определенным количеством почвы при регулярном поливе растения дождевой водой. За пять лет масса растения увеличилась на 76, 7 кг, а масса почвы уменьшилась всего на 60 г.
Учитель обращается к ученикам с вопросом.
– Как вы думаете, какой вывод на основании проведенного опыта сделал ван Гельмонт?
Учащиеся делают предположение, что масса дерева увеличилась за счет воды, использованной для полива.
На доске записывают первое исходное вещество – H2O.
Затем преподаватель знакомит учащихся с опытами Джозефа Пристли (1733-1804), о котором гимназисты знают из курса неорганической химии как об одном из первооткрывателей кислорода. Свой рассказ учитель заканчивает вопросом.
– К какому выводу пришел Дж. Пристли, проведя этот эксперимент?
Учащиеся, как и ученый, предполагают, что растения выделяют кислород, делая воздух пригодным для дыхания.
Позже голландский врач Ян Ингенхауз (1730-1799) обнаружил, что растения выделяют кислород лишь на солнечном свету и только зелеными частями.
На доске появляется запись:
H2O + солнечный свет + зеленая часть растений – –> O2.
В 1782 г. швейцарский физиолог растений Жан Сенебье (1742-1809) экспериментально доказал участие в процессе фотосинтеза углекислоты воздуха, которая служит растениям “пищей”. Вскоре после этого другой швейцарский исследователь, Никола Теодор де Соссюр (1767-1845), экспериментально доказал, что синтез органических соединений растениями обеспечивается питанием углекислотой (СO2) и H2O.
Эти открытия выдающихся ученых обобщаются учащимися в виде схемы на доске:
H2O + CO2 + солнечный свет + зеленая часть растений – –> O2 + органические вещества.
Раскрывая связь физиологического процесса с его физической основой, учитель обращает внимание учащихся на сделанное немецким врачом Р. Майером (1814-1878) в 1845 г. открытие, что зеленые растения преобразуют солнечную энергию в химическую энергию синтезируемых органических соединений согласно закону сохранения и превращения энергии.
Таким образом, уже к середине прошлого века о фотосинтезе сложились представления, которые можно выразить уравнением:
H2O + CO2 + солнечный свет + зеленая часть растений – –> O2 + органические вещества + химическая энергия.
Дальнейшие открытия связаны с именами французского физиолога растений Жана Батиста Буссенго (1802-1887), который точно измерил отношение поглощаемого при фотосинтезе углекислого газа к количеству выделяемого кислорода (1:1), и немецкого ботаника Юлиуса Сакса, продемонстрировавшего в 1864 г. образование зерен крахмала при фотосинтезе.
Учитель вновь обращается с вопросом к учащимся.
– Как вы думаете, каким образом Сакс показал, что органическим веществом, запасаемым зеленым растением при фотосинтезе, является крахмал?
Ученики, проводившие качественную реакцию на крахмал, отвечают на этот вопрос и уточняют, что с тех пор эта реакция называется пробой Сакса.
Таким образом, уже к началу XX в. были известны все компоненты фотосинтеза, как исходные, так и конечные:
Н2О + (СО2)n + солнечный свет + зеленая часть растений – –> (О2)n + крахмал + химическая энергия.
Итоговым результатом диалога учителя с учащимися является таблица “История фотосинтеза”
История фотосинтеза до XX в.
| Дата | Ученый | Вклад в науку |
| 1600 | Бельгийский естествоиспытатель ван Гельмонт | Поставил первый физиологический эксперимент, связанный с изучением питания растений |
| 1771 | Английский химик Джозеф Пристли | Пришел к выводу о том, что зеленые растения способны осуществлять реакции, противоположные тем дыхательным процессам, которые были обнаружены в тканях животных |
| 1779 | Голландский врач Ян Ингенхауз | Обнаружил, что растения выделяют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода |
| 1782 | Швейцарский физиолог растений Жан Сенебье | Экспериментально доказал, что все соединения углерода в растениях образуются из углекислого газа, который под влиянием света разлагается в зеленых органах растений с выделением кислорода |
| 1804 | Швейцарский агрохимик и физиолог растений де Соссюр | Пришел к выводу, что вода также потребляется растением при ассимиляции двуокиси углерода |
| 1845 | Немецкий физик и физиолог Роберт Майер | Показал, что открытый им закон сохранения энергии справедлив для явлений живой природы |
| 1864 | Французский физиолог растений Жан Буссенго | Установил соотношение объемов превращающегося в органические соединения СО2 и выделяемого О2 (1:1) |
| 1864 | Немецкий ботаник Юлиус Сакс | Продемонстрировал образование зерен крахмала при фотосинтезе |
После диалога с учащимися учитель задает следующий вопрос.
– Какие факторы должны, судя по уравнению фотосинтеза, влиять на скорость этого процесса?
Учащиеся называют эти факторы (свет, температура, концентрация СО2 в воздухе, влажность и т. д.), а учитель записывает их на доске в порядке поступления предложений. Затем учитель организует с учащимися 40-минутную практическую работу для проверки высказанных ими предположений о влиянии света и температуры на скорость фотосинтеза. Для проведения практической работы класс делится на группы.
Первая группа получает следующую инструктивную карточку.
Экспериментальная работа №1
“Зависимость продуктивности фотосинтеза от интенсивности света”
Цель работы: изучение зависимости продуктивности фотосинтеза от интенсивности света.
Материалы и оборудование: элодея или роголистник; NаНСО3 или (NH4)2CO3; отстоявшаяся водопроводная вода; стеклянная палочка; нитки; ножницы; электролампа мощностью 200 Вт; часы; термометр.
ХОД РАБОТЫ
1. Выберите здоровые побеги элодеи или роголистника длиной около 8 см, интенсивного зеленого цвета с неповрежденной верхушкой, подрежьте их под водой, привяжите ниткой к стеклянной палочке и опустите верхушкой вниз в стакан с водой комнатной температуры (температура воды должна оставаться постоянной).
2. Для опыта взять отстоявшуюся водопроводную воду, обогащенную CO2 путем внесения NаНСО3 или (NН4)2СО3, и выставить стакан с водным растением на яркий свет. Наблюдайте за появлением пузырьков воздуха из среза растения.
3. Когда ток пузырьков станет равномерным, подсчитайте количество пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин, данные запишите в таблицу. Определите средний результат.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
| № п/п | Интенсивность света | Первое измерение | Второе измерение | Третье измерение | Среднее значение |
4. Затем стакан с растением удалите от света на 50-60 см и повторите действия, указанные в п. 3.
5. Сравните результаты опытов и сделайте письменный вывод о различной интенсивности фотосинтеза на ярком и слабом свету.
В конце работы учащиеся первой группы отвечают на вопрос.
– Почему теневыносливые растения используют свет низкой интенсивности более эффективно, чем светолюбивые растения?
Учащиеся второй группы получают другое задание.
Экспериментальная работа №2
“Зависимость продуктивности фотосинтеза от спектрального состава света”
Цель работы: выяснение зависимости интенсивности фотосинтеза от спектрального состава света.
Материалы и оборудование: элодея или роголистник; набор светофильтров; семь высоких широкогорлых банок; отстоявшаяся водопроводная вода; ножницы; электролампа мощностью 200 Вт; часы; термометр; пробирки.
ХОД РАБОТЫ
1. Наполните пробирку на 2/3 объема отстоявшейся водопроводной водой и поместите в нее водное растение верхушкой вниз. Стебель подрежьте под водой.
2. Поместите в высокую широкогорлую банку синий светофильтр (круговой), поставьте в нее пробирку с растением и выставьте прибор на яркий свет так, чтобы он попадал на растение, проходя через светофильтр. Наблюдайте за появлением пузырьков воздуха из среза стебля растения.
3. Когда ток пузырьков станет равномерным, подсчитайте количестве пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин. Определите средний результат, данные запишите в таблицу.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
| № п/п | Светофильтр | Первое измерение | Второе измерение | Третье измерение | Среднее значение |
4. Смените синий светофильтр на красный и повторите действия, указанные в п. 3. Следите за тем, чтобы расстояние от источника света и температура воды оставались постоянными.
5. Сравните результаты опытов и сделайте письменный вывод о зависимости интенсивности фотосинтеза от спектрального состава света.
После проведения работы учащиеся второй группы записывают ответ на вопрос.
– В каких лучах интенсивность фотосинтеза, судя по выделению кислорода, будет выше и почему?
Для учащихся третьей группы предложена следующая задача.
Экспериментальная работа №3
“Зависимость интенсивности фотосинтеза от температуры”
Цель работы: изучение зависимости интенсивности фотосинтеза от температуры.
Материалы и оборудование: элодея или роголистник; три высокие широкогорлые банки; отстоявшаяся водопроводная вода; ножницы; пробирки; электролампа мощностью 200 Вт; часы; термометр.
ХОД РАБОТЫ
1. Наполните пробирку на 2/3 объема отстоявшейся водопроводной водой и поместите в нее водное растение верхушкой вниз. Стебель подрежьте под водой.
2. Налейте в три высокие широкогорлые банки отстоявшуюся водопроводную воду разной температуры (+4 °С, +25 °С и +40 °С), поместите пробирку с растением в банку с водой с температурой +25 °С и выставьте прибор на яркий свет. Наблюдайте за появлением пузырьков воздуха из среза стебля растения.
2. Через 5 мин подсчитайте количество пузырьков, выделившихся за 1 мин. Подсчет проведите 3 раза с перерывом в 1 мин. Определите средний результат, данные запишите в таблицу.
Таблица. количество пузырьков О2, выделившихся за 1 мин из водного растения
| № п/п | Температура | Первое измерение | Второе измерение | Третье измерение | Среднее значение |
4. Перенесите пробирку с растением в банку с водой другой температуры и повторите действия, указанные в п. 3. Следите за тем, чтобы расстояние от источника света и температура воды оставались постоянными.
5. Сравните результаты опытов и сделайте письменный вывод о влиянии температуры на интенсивность фотосинтеза.
После проведенных исследований учащиеся третьей группы должны ответить на вопрос.
– Почему при более высоких или более низких, чем 25 °С, температурах скорость фотосинтеза снижается?
По окончании практической работы учащиеся каждой группы докладывают результаты своих опытов, остальные учащиеся записывают их в своих тетрадях.
В конце занятия учащиеся делают итоговый вывод о влиянии различных факторов среды на интенсивность фотосинтеза.
Продолжением этой работы является семинар-дискуссия, на который выносятся следующие вопросы.
1. История фотосинтеза с начала XX в. до наших дней.
2. Влияние загрязнителей окружающей среды (сернистого газа, тяжелых металлов и др.) на фотосинтез у растений.
3. Космическая роль зеленых растений.
По этим вопросам учащиеся готовят сообщения и обсуждают их.
Результатом этой работы должно стать оформление продолжения таблицы “История фотосинтеза”.
История фотосинтеза в XX в.
| Год | Ученый | Вклад в науку |
| 1905 | Английский физиолог растений Ф. Ф. Блэкмэн | Высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т. е. световую, реакцию и нефотохимическую, т. е. темновую, реакцию |
| 1932 | Эмерсон и Арнольд | Сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию |
| 1937 | Р. Хилл | Обнаружил, что изолированные хлоропласты способны высвобождать кислород в присутствии окислителя (акцептора электронов) |
| 1940 | Г. Фишер | Определил структурную формулу хлорофилла в опытах с последовательным разрушением молекул пигмента |
| 1941 | Рубен и Камен | Установили факт разложения воды, приводящего к выделению свободного кислорода при фотосинтезе |
|
1946 1953 |
Бассэм, Бенсон, Кальвин | Определили последовательность темновых реакций фотосинтеза |
| 1954 | Арнон, Аллен и Уотли | Продемонстрировали процесс фотосинтеза в искусственных условиях вне клетки |
При обсуждении роли загрязнителей окружающей среды, влияющих на интенсивность фотосинтеза, учащиеся обсуждают в первую очередь роль сернистого газа (SO2) и серной кислоты (H2SO4), так как параллельно по неорганической химии они изучают тему “Серная кислота и ее производные”.
Обсуждая космическую роль зеленых растений, учащиеся в итоге подводят “баланс” продуктивности фотосинтеза:
“1 см2 листа сахарной свеклы в течение дня образует из воздуха и воды с помощью солнечного света 0, 001 г сахара.
Ежегодно наземные растения связывают почти 20 млрд т, а растения морей – 25 млрд т углерода, которые расходуются на образование свыше 100 млрд т сахара. Для перевозки такого количества сахара потребовался бы железнодорожный состав длиной более чем в 50 млн км, что в 40 раз превышает общую длину всех железнодорожных линий Земли.
Для синтеза 1 кг глюкозы растениям достаточно затратить около 4, 4 кВт х ч электроэнергии, т. е. примерно столько же, сколько потребляет цветной телевизор за 15 ч.
При фотосинтезе растения используют за год почти 500 трлн кВт х ч солнечной энергии, что примерно в 200 раз превышает мировую выработку электроэнергии за год”.
Определение загрязнения окружающей среды по изменениям комплекса признаков хвойных деревьев
Hayчно-исследовательская деятельность гимназистов при разработке этой темы заключается в определении морфологических и биохимических изменений у растений под действием антропогенных факторов.
Известно, что на загрязнение среды наиболее сильно реагируют хвойные древесные растения. Характерными признаками неблагополучия окружающей среды, и особенно газового состава атмосферы, является появление разного рода некрозов, уменьшение размеров многих органов (хвои, побегов текущего года и прошлых лет, их толщины, размера шишек, размера и числа заложившихся почек). Ввиду уменьшения роста побегов и хвои в длину у растений в загрязненной зоне наблюдается сближенность хвоинок (их число на 10 см побега больше, чем в чистой зоне).
Использование хвойных в качестве биоиндикаторов дает возможность оценить состояние окружающей среды. Для исследований предложен район памятника Славы, где растения оценивались как по качественным, так и по количественным показателям. Объектом исследований выбран вид ель обыкновенная. ветви дерева срезаны на высоте 2 м с части кроны, находящейся ближе к Задонскому шоссе (зоне с загрязненным воздухом). Контролем могут служить ветви деревьев, собранные в зеленой зоне города (Центральный парк культуры и отдыха).
При осмотре хвои с помощью лупы выявлено, что кончики большинства хвоинок имеют желто-коричневый цвет, что свидетельствует о загрязнении воздуха. Другие качественные признаки (колючесть, ломкость, смолистость) не позволили однозначно оценить состояние окружающей среды в исследуемом районе.
Длина хвоинок на побегах прошлого года у растений опытной группы в большинстве случаев была меньше таковой для контрольных образцов, однако количество листьев ели на 10 см побега исследуемой зоны не отличались от аналогичных показателей, характерных для чистой зоны. Толщина хвоинок варьировала от 1 до 5 мм. Количество сформировавшихся почек для опытных образцов равнялось 4±2, для контрольных 8±2.
На основании проведенных наблюдений и измерений можно сделать вывод о средней степени загрязнения воздуха в районе памятника Славы.
Определение содержания хлорофилла фотометрическим методом. Биоиндикация состояния окружающей среды
Биоиндикация – это метод оценки действия экологических факторов при помощи биологических систем. Например, воздействие SO2 на хвойные породы выражено в угнетении их роста и некрозе хвои. Содержание хлорофилла в листьях может также служить неспецифическим биоиндикационным признаком, т. к. снижение содержания хлорофилла наблюдается до появления видимых изменений листьев.
В качестве биоиндикатора выбран чувствительный к выхлопным газам автомобилей вид сосны обыкновенной, листья которой были собраны вблизи окружной автодороги.
Метод основан на регистрации оптических характеристик ацетоновой вытяжки хлорофилла, полученных с использованием фотоэлектроколориметра КФК-2. Для определения содержания хлорофилла в листьях использовали метод построения калибровочной кривой с использованием стандартного раствора Гетри.
Массовое соотношение взятого для исследований материала (листьев) и экстрагирующей жидкости (ацетона) равно 1:100.
В результате проведенных экспериментов показано, что в спектре поглощения молекул хлорофилла комнатного растения, выбранного в качестве контроля, имеются максимумы в синей (440 нм) и красной (670 нм) областях. Для ацетоновой вытяжки листьев сосны наблюдается смещение коротковолнового максимума поглощения в фиолетовую область (400 нм). Величины максимумов зависят от количества хлорофилла в вытяжке, которое рассчитывают на основании этих измерений.
Установлено, что содержание хлорофилла в контрольном и опытном образцах составило величину 0, 14±0, 02 мг/г листа, что соответствует нормальному количеству зеленого пигмента в листьях сосны.
Таким образом, не выявлено значительных изменений в содержании хлорофилла в контрольном и опытном образцах. Это свидетельствует о том, что содержание диоксида серы в воздухе и в выпадающих осадках (дожде, снеге) находится в пределах нормы.
Источник: belreferatov.net