Органеллы эукариотической клетки
Клетки, образующие ткани животных и растений, значительно различаются по форме, размерам и внутреннему строению. Однако все они обнаруживают сходство в главных чертах процессов жизнедеятельности, обмена веществ, в раздражимости, росте, развитии, способности к изменчивости.
Клетки всех типов содержат два основных компонента, тесно связанных между собой, — цитоплазму и ядро. Ядро отделено от цитоплазмы пористой мембраной и содержит ядерный сок, хроматин и ядрышко. Полужидкая цитоплазма заполняет всю клетку и пронизана многочисленными канальцами. Снаружи она покрыта цитоплазматической мембраной. В ней имеются специализированные структуры-органоиды, присутствующие в клетке постоянно, и временные образования — включения.Мембранные органоиды: наружная цитоплазматическая мембрана (HЦM), эндоплазматическая сеть (ЭПС), аппарат Гольджи, лизосомы, митохондрии и пластиды. В основе строения всех мембранных органоидов лежит биологическая мембрана. Все мембраны имеют принципиально единый план строения и состоят из двойного слоя фосфолипидов, в который с различных сторон ива разную глубину погружены белковые молекулы. Мембраны органоидов отличаются друг от друга лишь наборами входящих в них белков.

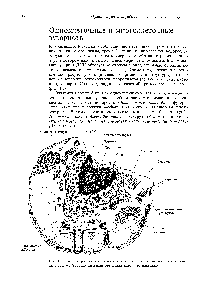
Схема строения эукариотической клетки. А — клетка животного происхождения; Б — растительная клетка:1 — ядро с хроматином и ядрышком, 2 — цитоплазматическая мембрана, 3- клеточная стенка, 4 — поры в клеточной стенке, через которые сообщается цитоплазма соседних клеток, 5 — шероховатая эндоплазматическая сеть, б — гладкая эндоплазматическая сеть, 7 — пиноцитозная вакуоль, 8 — аппарат (комплекс) Гольджи, 9 — лизосома, 10 — жировые включения в каналах гладкой эндоплазматической сети, 11 — клеточный центр, 12 — митохондрия, 13 -свободные рибосомы и полирибосомы, 14 — вакуоль, 15 — хлоропласт
Цитоплазматическая мембрана. У всех клеток растений, многоклеточных животных, у простейших и бактерий клеточная мембрана трехслойна: наружный и внутренний слои состоят из молекул белков, средний — из молекул липидов. Она ограничивает цитоплазму от внешней среды, окружает все органоиды клетки и представляет собой универсальную биологическую структуру.
некоторых клетках наружная оболочка образована несколькими мембранами, плотно прилегающими друг к другу. В таких случаях клеточная оболочка становится плотной и упругой и позволяет сохранить форму клетки, как, например, у эвглены и инфузории туфельки. У большинства растительных клеток, помимо мембраны, снаружи имеется еще толстая целлюлозная оболочка — клеточная стенка. Она хорошо различима в обычном световом микроскопе и выполняет опорную функцию за счет жесткого наружного слоя, придающего клеткам четкую форму.
На поверхности клеток мембрана образует удлиненные выросты — микроворсинки, складки, впячивания и выпячивания, что во много раз увеличивает всасывающую или выделительную поверхность. С помощью мембранных выростов клетки соединяются друг с другом в тканях и органах многоклеточных организмов, на складках мембран располагаются разнообразные ферменты, участвующие в обмене веществ. Отграничивая клетку от окружающей среды, мембрана регулирует направление диффузии веществ и одновременно осуществляет активный перенос их внутрь клетки (накопление) или наружу (выделение). За счет этих свойств мембраны концентрация ионов калия, кальция, магния, фосфора в цитоплазме выше, а концентрация натрия и хлора ниже, чем в окружающей среде. Через поры наружной мембраны из внешней среды внутрь клетки проникают ионы, вода и мелкие молекулы других веществ. Проникновение в клетку относительно крупных твердых частиц осуществляется путем фагоцитоза (от греч.
171;фаго» — пожираю, «питое» — клетка). При этом наружная мембрана в месте контакта с частицей прогибается внутрь клетки, увлекая частицу в глубь цитоплазмы, где она подвергается ферментативному расщеплению. Аналогичным путем в клетку попадают и капли жидких веществ; их поглощение называетсяпиноцитозом (от греч. «пино» — пью, «цитос» — клетка). Наружная клеточная мембрана выполняет и другие важные биологические функции.
Цитоплазма на 85 % состоит из воды, на 10 % — из белков, остальной объем приходится на долю липидов, углеводов, нуклеиновых кислот и минеральных соединений; все эти вещества образуют коллоидный раствор, близкий по консистенции глицерину. Коллоидное вещество клетки в зависимости от ее физиологического состояния и характера воздействия внешней среды имеет свойства и жидкости, и упругого, более плотного тела. Цитоплазма пронизана каналами различной формы и величины, которые получили название эндоплазматической сети. Их стенки представляют собой мембраны, тесно контактирующие со всеми органоидами клетки и составляющие вместе с ними единую функционально-структурную систему для осуществления обмена веществ и энергии и перемещения веществ внутри клетки.
В стенках канальцев располагаются мельчайшие зернышки-гранулы, называемые рибосомами. Такая сеть канальцев называется гранулярной. Рибосомы могут располагаться на поверхности канальцев разрозненно или образуют комплексы из пяти-семи и более рибосом, называемые полисомами.Другие канальцы гранул не содержат, они составляют гладкую эндоплазматическую сеть. На стенках располагаются ферменты, участвующие в синтезе жиров и углеводов.
Внутренняя полость канальцев заполнена продуктами жизнедеятельности клетки. Внутриклеточные канальцы, образуя сложную ветвящуюся систему, регулируют перемещение и концентрацию веществ, разделяют различные молекулы органических веществ и этапы их, синтеза. На внутренней и внешней поверхности мембран, богатых ферментами, осуществляется синтез белков, жиров и углеводов, которые либо используются в обмене веществ, либо накапливаются в цитоплазме в качестве включений, либо выводятся наружу.
Рибосомы встречаются во всех типах клеток — от бактерий до клеток многоклеточных организмов. Это округлые тельца, состоящие из рибонуклеиновой кислоты (РНК) и белков почти в равном соотношении. В их состав непременно входит магний, присутствие которого поддерживает структуру рибосом. Рибосомы могут быть связаны с мембранами эндоплазматической сети, с наружной клеточной мембраной или свободно лежать в цитоплазме. В них осуществляется синтез белков. Рибосомы кроме цитоплазмы встречаются в ядре клетки. Они образуются в ядрышке и затем поступают в цитоплазму.
Комплекс Гольджи в растительных клетках имеет вид отдельных телец, окруженных мембранами. В животных клетках этот органоид представлен цистернами, канальцами и пузырьками. В мембранные трубки комплекса Гольджи из канальцев эндоплазматической сети поступают продукты секреции клетки, где они химически перестраиваются, уплотняются, а затем переходят в цитоплазму и либо используются самой клеткой, либо выводятся из нее. В цистернах комплекса Гольджи происходит синтез полисахаридов и их объединение с белками, в результате чего образуются гликопротеиды.
Митохондрии — небольшие тельца палочковидной формы, ограниченные двумя мембранами. От внутренней мембраны митохондрии отходят многочисленные складки — кристы, на их стенках располагаются разнообразные ферменты, с помощью которых осуществляется синтез высокоэнергетического вещества — аденозинтрифосфорной кислоты (АТФ). В зависимости от активности клетки и внешних воздействий митохондрии могут перемещаться, изменять свои размеры, форму. В митохондриях найдены рибосомы, фосфолипиды, РНК и ДНК. С присутствием ДНК в митохондриях связывают способность этих органоидов к размножению путем образования перетяжки или почкованием в период деления клетки, а также синтез части митохондриальных белков.
Лизосомы — мелкие овальные образования, ограниченные мембраной и рассеянные по всей цитоплазме. Встречаются во всех клетках животных и растений. Они возникают в расширениях эндоплазматической сети и в комплексе Гольджи, здесь заполняются гидролитическими ферментами, а затем обособляются и поступают в цитоплазму. В обычных» условиях лизосомы переваривают частицы, попадающие в клетку путем фагоцитоза, и органоиды отмирающих клеток. Продукты лизиса выводятся через мембрану лизосомы в цитоплазму, где они включаются в состав новых молекул. При разрыве лизоеомной мембраны ферменты поступают в цитоплазму и переваривают ее содержимое, вызывая гибель клетки.
Пластиды есть только в растительных клетках и встречаются, у большинства зеленых растений. В пластидах синтезируются и накапливаются органические вещества. Различают пластиды трех видов: хлоропласты, хромопласты и лейкопласты.
Хлоропласты — зеленые пластиды, содержащие зеленый пигмент хлорофилл. Они находятся в листьях, молодых стеблях, незрелых плодах. Хлоропласты окружены двойной мембраной. У высших растений внутренняя часть хлоропластов заполнена полужидким веществом, в котором параллельно друг другу уложены пластинки. Парные мембраны пластинок, сливаясь, образуют стопки, содержащие хлорофилл (рис. 6). В каждой стопке хлоропластов высших растений чередуются слои молекул белка и молекул липидов, а между ними располагаются молекулы хлорофилла. Такая слоистая структура обеспечивает максимум свободных поверхностей и облегчает захват и перенос энергии в процессе фотосинтеза.
Хромопласты — пластиды, в которых содержатся растительные пигменты (красный или бурый, желтый, оранжевый). Они сосредоточены в цитоплазме клеток цветков, стеблей, плодов, листьев растений и придают им соответствующую окраску. Хромопласты образуются из лейкопластов или хлоропластов в результате накопления пигментов каротиноидов.
Лейкопласты-бесцветные пластиды, располагающиеся в неокрашенных частях растений: в стеблях, корнях, луковицах и др. В лейкопластах одних клеток накапливаются зерна крахмала, в лейкопластах других клеток — масла, белки.
Все пластиды возникают из своих предшественников — пропластид. В них выявлена ДНК, которая контролирует размножение этих органоидов.
Клеточный центр, или центросома, играет важную роль при делении, клетки и состоит из двух центриолей. Он встречается у всех клеток животных и растений, кроме цветковых, низших грибов и некоторых, простейших. Центриоли в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В делящейся клетке первым делится клеточный центр, одновременно образуется ахроматиновое веретено, ориентирующее хромосомы при расхождении их к полюсам. В дочерние клетки отходит по одной центриоле.
У многих растительных и животных клеток имеются органоиды специального назначения: реснички, выполняющие функцию движения (инфузории, клетки дыхательных путей), жгутики (простейшие одноклеточные, мужские половые клетки у животных и растений и др.). Включения —временные элемеаты, возникающие в клетке на определенной стадии ее жизнедеятельности в результате синтетической функции.
и либо используются, либо выводятся из клетки. Включениями являются также запасные питательные вещества: в растительных клетках-крахмал, капельки жира, блки, эфирные масла, многие органические кислоты, соли органических и неорганических кислот; в животных клетках — гликоген (в клетках печени и мышцах), капли жира (в подкожной клетчатке); Некоторые включения накапливаются в клетках как отбросы — в виде кристаллов, пигментов и др.
Вакуоли — это полости, ограниченные мембраной; хорошо выражены в клетках растений и имеются у простейших. Возникают в разных участках расширений эндоплазматической сети. И постепенно отделяются от нее. Вакуоли поддерживают тургорное давление, в них сосредоточен клеточный или вакуолярный сок, молекулы которого определяют его осмотическую концентрацию. Считается, что первоначальные продукты синтеза — растворимые углеводы, белки, пектины и др. — накапливаются в цистернах эндоплазматической сети. Эти скопления и представляют собой зачатки будущих вакуолей.
Цитоскелет. Одной из отличительных особенностей эукариотической клетки является развитие в ее цитоплазме скелетных образований в виде микротрубочек и пучков белковых волокон. Элементы цитоскелета тесно связаны с наружной цитоплазматической мембраной и ядерной оболочкой, образуют сложные переплетения в цитоплазме. Опорные элемеиты цитоплазмы определяют форму клетки, обеспечивают движение внутриклеточных структур и перемещение всей клетки.
Ядро клетки играет основную роль в ее жизнедеятельности, с его удалением клетка прекращает свои функции и гибнет. В большинстве животных клеток одно ядро, но встречаются и многоядерные клетки (печень и мышцы человека, грибы, инфузории, зеленые водоросли). Эритроциты млекопитающих развиваются из клеток-предшественников, содержащих ядро, но зрелые эритроциты утрачивают его и живут недолго.
Ядро окружено двойной мембраной, пронизанной порами, посредством которых оно тесно связано с каналами эндоплазматической сети и цитоплазмой. Внутри ядра находится хроматин — спирализованные участки хромосом. В период деления клетки они превращаются в палочковидные структуры, хорошо различимые в световой микроскоп. Хромосомы — это сложный комплекс белков с ДНК, называемый нуклеопротеидом.
Функции ядра состоят в регуляции всех жизненных отправлений клетки, которую оно осуществляет при помощи ДНК и РНК-материальных носителей наследственной информации. В ходе подготовки к делению клетки ДНК удваивается, в процессе митоза хромосомы расходятся и передаются дочерним клеткам, обеспечивая преемственность наследственной информации у каждого вида организмов.
Кариоплазма — жидкая фаза ядра, в которой в растворенном виде находятся продукты жизнедеятельности ядерных структур
Ядрышко — обособленная, наиболее плотная часть ядра. В состав ядрышка входят сложные белки и РНК, свободные или связанные фосфаты калия, магния, кальция, железа, цинка, а также рибосомы. Ядрышко исчезает перед началом деления клетки и вновь формируется в последней фазе деления.
Таким образом, клетка обладает тонкой и весьма сложной организацией. Обширная сеть цитоплазматических мембран и мембранный принцип строения органоидов позволяют разграничить множество одновременно протекающих в клетке химических реакций. Каждое из внутриклеточных образований имеет свою структуру и специфическую функцию, но только при их взаимодействии возможна гармоничная жизнедеятельность клетки.На основе такого взаимодействия вещества из окружающей среды поступают в клетку, а отработанные продукты выводятся из нее во внешнюю среду — так совершается обмен веществ. Совершенство структурной организации клетки могло возникнуть только в результате длительной биологической эволюции, в процессе которой выполняемые ею функции постепенно усложнялись.
Простейшие одноклеточные формы представляют собой и клетку, и организм со всеми его жизненными проявлениями. В многоклеточных организмах клетки образуют однородные группы — ткани. В свою очередь ткани формируют органы, системы, и их функции определяются общей жизнедеятельностью целостного организма.
Помимо организмов с типичной клеточной организацией (эукариотические клетки) существуют относительно простые, доядерные, илипрокариотические, клетки — бактерии и синезеленые, у которых отсутствуют оформленное ядро, окруженное ядерной мембраной, и высокоспециализированные внутриклеточные органоиды. Особую форму организации живого представляют вирусы и бактериофаги (фаги). Их строение крайне упрощено: они состоят из ДНК (либо РНК) и белкового футляра. Свои функции обмена веществ и размножения вирусы и фаги осуществляют только внутри клеток другого организма: вирусы — внутри клеток растений и животных, фаги — в бактериальных клетках как паразиты на, генетическом уровне.
Источник: www.examen.ru
Органеллы эукариотических клеток
Прокариотическая клетка не имеет внутренних перегородок. Она состоит как бы из одного отсека, отграниченного от внешней среды плазматической мембраной. У эукариотической клетки таких отсеков несколько, т. е. эти клетки разделены на различные, окруженные мембранами области (компартменты). Комплименты эукариотической клетки называются органами. Органеллы специализируются на выполнении различных функций. Разделение клетки на компартменты, возможно, является следствием больших размеров современной эукариотической клетки: в среднем она больше бактериальной в 10 — 30 раз. При этом принципиально важными оказываются два момента:
• увеличение объема клетки сопровождается существенно меньшим увеличением площади поверхности ее плазматической мембраны, тогда как именно на мембранах протекают многие важнейшие метаболические процессы;
• с увеличением объема клетки затрудняется координация ее метаболизма, в частности рациональное распределение по объему множества катаболических и анаболических процессов, «доставка» нужных веществ в нужное место.
Формирование компартментов позволяет пространственно разделить «блоки» метаболизма.
В цитозоле любой (не только растительной) современной эукариотической клетки находятся следующие органеллы:
1) ядро — содержит основную часть генома и является местом синтеза ДНК и РНК;
2) эндоплазматический ретикулум (ЭР) — место синтеза большинства липидов клетки, а также белков, предназначенных для органелл или секреции;
3) аппарат Гольджи (АГ)— место сортировки и модификации белков и липидов, получаемых от эндоплазматического ретикулума;
4) митохондрии — «энергетические станции клетки», основное место синтеза АТФ, имеют собственный геном и белоксинтезирующую систему;
5) пероксисомы — здесь происходят многие окислительные процессы;
6) лизосомы (в растительных клетках — литические вакуоли) — место компартментации литических ферментов.
Растительная клетка наряду с этими органеллами обязательно содержит:
1) пластиды — органеллы, выполняющие различные функции в зависимости от типа клетки, в которой находятся; основная функция — фотосинтез; имеют собственный геном и белоксинтезирующую систему;
2) вакуоли — органеллы, выполняющие несколько функций, в том числе литические (литические вакуоли) и запасающие (запасающие вакуоли).
Классификацию органелл эукариотической клетки целесообразно построить на предполагаемой схеме ее эволюции. Практически доказано, что митохондрии и пластиды имеют симбиотическое происхождение, т. е. являются потомками микроорганизмов, «захваченных» предшественником эукариотической клетки. При этом внешняя мембрана пластид и митохондрий соответствует плазматической мембране «хозяйской» клетки, внутренняя — плазматической мембране захваченной бактерии, а строма пластид и матрикс митохондрий — бактериальной цитоплазме (рис. 1).
Происхождение ядра менее ясно. Скорее всего, двухслойная ядерная оболочка образовалась из глубокого впячивания плазматической мембраны. В результате часть цитоплазмы оказалась изолированной от остального пространства клетки. Следовательно, внутреннее пространство ядра происходит из цитозоля и при определенных условиях эти пространства могут объединиться (иногда для обозначения подобных пространств применяют математический термин «топологически эквивалентные пространства»). Известно, что во время митоза у высших эукариот ядерная оболочка разрушается и содержимое ядра полностью смешивается с цитозолем. Этого не происходит ни с одной другой мембранной органеллой.
Эндоплазматический ретикулум, аппарат Гольджи и вакуоли образовались, по-видимому, также за счет впячивания плазматической мембраны с последующим «отшнуровыванием». Если это верно, то внутренняя поверхность мембран этих органелл должна быть топологически эквивалентна внешней поверхности плазмалеммы. Это справедливо для ЭР, АГ, вакуолей и многих промежуточных везикул, участвующих в эндоцитозе, экзоцитозе и транспортных процессах между этими органеллами.
Рис. 1. Схема предполагаемого эволюционного происхождения ядра и хлоропластов (пластид):
А — происхождение ядра и ЭР; Б — происхождение пластид
Пероксисомы представляются «потомками» древних органелл, возникших для защиты клеток от кислорода, накопившегося в атмосфере в результате оксигенного фотосинтеза.
Исходя из сказанного основные внутриклеточные компартменты эукариотической клетки можно разделить на пять групп:
1) ядро и цитозоль — разделены мембраной, выполняют разные функции, но топологически эквивалентны;
2) митохондрии;
3) пластиды (только в растительной клетке;
4) пероксисомы;
5) эндомембранная система клетки — остальные мембранные органеллы: ЭР, аппарат Гольджи, вакуоли (в растительных клетках), лизосомы (в животных клетках), транспортные везикулы.
Целесообразность объединения последней группы органелл в одну систему вытекает не только из эволюционной схемы, но и по функциональным признакам, о которых будет сказано позже.
Транспорт и сортировка белков в эукариотической клетке
С возникновением компартментов эукариотическая клетка получает не только очевидные преимущества, но и ряд проблем. Одна из них связана с необходимостью сортировки и доставки нужных соединений в определенные органеллы. Прежде всего это касается белков. Судьба синтезированных белков различна и зависит от мест их последующего функционирования. Существуют два магистральных пути транспорта белков, которые начинаются в разных местах цитоплазмы (рис. 2).
Рис. 2. Схема двух путей транспорта белков в растительной клетке.
Цифры — сигналы сортировки белков, описанные в табл. 1;— путь, не требующий специального сигнала сортировки
Первая транспортная ветвь обеспечивает белками пластиды, митохондрии, ядро и пероксисомы, т. е. все компартменты клетки, кроме органелл эндомембранной системы. Синтез этих белков происходит на свободных рибосомах цитозоля. Белки, предназначенные для транспорта, содержат сигналы сортировки, направляющие их в соответствующие органеллы. Подобными сигналами обычно служат один или несколько участков белка, которые называют сигнальными или лидерными пептидами. В мембране органеллы находится специальный белок-транслокатор, который «узнает» сигнальный пептид. Молекула транспортируемого белка должна развернуться, чтобы подобно нитке развернувшегося клубка «продеться» через «игольное ушко» белка-транслокатора. В табл. 1 представлены некоторые характеристики различных сигналов сортировки в растительной клетке. Этот путь транспорта белков иногда называют цитозольным. Следует отметить, что большинство белков, синтезируемых на свободных рибосомах цитозоля, не имеют сигналов сортировки и остаются в цитозоле в качестве постоянных компонентов.
Другая транспортная ветвь используется для секретируемых белков, а также для белков, предназначенных для органелл эндомембранной системы и плазматической мембраны. Синтез этих белков также начинается на рибосомах цитозоля, но после инициации трансляции рибосомы прикрепляются к мембране ЭР, при этом формируется шероховатый ЭР. Образующиеся белки переносятся в ЭР по мере их синтеза, т. е. котрансляционно. Это означает, что очередной участок полипептидной цепи сразу после синтеза пересекает мембрану ЭР. Некоторые из белков попадают в просвет ЭР, другие закрепляются в мембране и становятся трансмембранными белками ЭР. Эту транспортную ветвь часто называют секреторным путем клетки.
Таблица 1. Сигнальные последовательности для транспорта белков в растительной клетке (цифры в скобках — пути транспорта, показанные на рис. 2)
|
Целевая органелла |
Сигнальная последовательность |
Характеристика |
|
Хлоропласта: строма (9) люмен и мембраны тилакоидов (9 ) Митохондрии: матрикс (8) внутренняя мембрана, межмембранное пространство (8) Пероксисомы(6) Ядро (7) Сигнальный пептид секреторного пути (1) Эндоплазматический ретикулум (2) Вакуоль (3,4) |
N-концевой лидерный пептид («стромальный») Два последовательных N-концевых лидерных пептида N-концевой пресиквенс Два последовательных N-концевых пресиквенса Сигналы пероксисомальной локализации PTS1 и PTS2 Сигналы ядерной локализации NLS. Не отщепляются после переноса белка в ядро N-концевой лидерный пептид Сигнал локализации в ЭР Сигналы локализации в вакуолях: NTPP, СТРР, внутрибелковый сигнал |
Последовательность из 40—50 аминокислот Первый пептид — «стромальный», второй — «люменальный» Формирует положительно заряженную амфипатическую а -петлю Первый пресиквенс — как для белков матрикса, второй состоит из остатков гидрофобных аминокислот PTS1 — С-концевой трипептид — Ser-Lys-Leu PTS2 локализован на N-конце NLS типа 1: Pro-Lys-Lys-Lys-Arg- Lys NLS типа 2: две последовательности, разделенные спенсером NLS типа 3: Lys-Ile-Pro-Ile-Lys 10—15 остатков гидрофобных аминокислот, формирующих а-спираль С-концевой тетрапептид KDEL (Lys-Asp-Glu-Leu) NTPP — N-концевой сигнал: Asn-Pro-lle-Arg СТРР — С-концевой сигнал |
Две ветви транспорта различаются схемой транспортировки. Пути цитозольного транспорта белков параллельны, т. е. из цитозоля белки сразу направляются в нужную органеллу. С момента высвобождения белка в цитозоль до поступления его в органеллу проходит не больше 1 — 2 мин.
Транспорт белков по секреторному пути происходит последовательно — от органеллы к органелле. До достижения конечного пункта белок может побывать в нескольких органеллах (ЭР, разные отделы АГ). Путь от мембраны ЭР до места назначения может занять десятки и даже сотни минут. В процессе переноса белки могут претерпевать значительные модификации (прежде всего в АГ). На заключительных этапах транспорт может происходить параллельно: в вакуоль, периплазматическое пространство или в плазмалемму.
И наконец, два пути транспорта белков различаются по механизму переноса молекул. Для цитозольного пути возможен только мономолекулярный механизм транспорта белков, при котором каждая молекула белка индивидуально пересекает мембрану через соответствующий транслокатор. Для секреторного пути характерен везикулярный механизм транспорта белковых молекул, который опосредован транспортными пузырьками (везикулами). Везикулы отшнуровываются от одного компартмента, при этом происходит захват определенных молекул из его полости. Затем везикулы сливаются с другим компартментом, доставляя в него свое содержимое. При везикулярном транспорте белки не пересекают никаких мембран, транспорт возможен только между топологически эквивалентными компартментами. Везикулярный механизм транспорта избирательно контролируется с помощью специальных белков, выполняющих функции сигналов сортировки. В транспортный пузырек белок попадает, если его сигнал сортировки связывается с рецептором на мембране везикулы. В настоящее время в составе белков установлены лишь некоторые сигналы сортировки, большинство комплементарных им мембранных рецепторов остаются пока неизвестными.
Используемая литература: Физиология растений: Учебник для стул, вузов / Н.Д. Алехина,
Ф504 10. В. Балнокин. В.Ф. Гаврилснко и Др.; Под ред. И. П. Ермакова. —
М.: Издательский центр «Академия», 2005. — 640 с.
Скачать реферат: У вас нет доступа к скачиванию файлов с нашего сервера. КАК ТУТ СКАЧИВАТЬ
Пароль на архив: privetstudent.com
Источник: privetstudent.com
Источник: www.chem21.info