Центральная вакуоль растительной клетки
Клетки растений содержат те же внутриклеточные отделы, что и животные. Но вместе с тем в растительных клетках присутствуют два особых типа компартментов, окруженных мембраной – это вакуоли и пластиды. Наличие вакуолей и пластид связано с неподвижным образом жизни растительной клетки. Вакуоли растений, как высших, так и низших, выполняют важные физиологические функции.
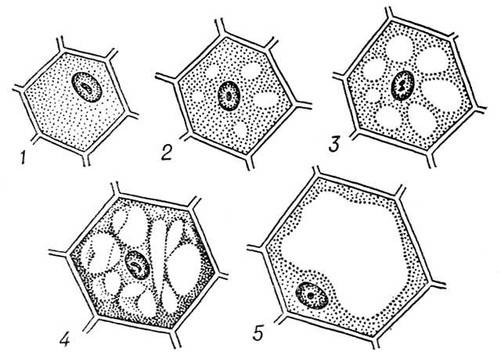
Развитие вакуолей начинается с момента образования клетки после деления. Из мелких пузырьков, отделившихся от ЭПС, возникают мелкие центральные вакуоли. Они постепенно увеличиваются в объеме, что совпадает с ростом и дифференцировкой клеток, и сливаются друг с другом, оттесняя цитоплазму вместе с органоидами и ядро клетки к периферии.
Полностью развитые растительные клетки обычно содержат одну крупную центральную вакуоль, окруженную полупроницаемой мембраной — тонопластом. Он отличается большей механической плотностью, чем плазматическая мембрана, но по своей природе сходен с ней. Он образован трехслойной липопротеидной мембраной, которая богата полярными молекулами липидов.
Полость вакуоли заполнена клеточным соком. Клеточный сок – это водный раствор веществ, которые с физиологической точки зрения принадлежат к двум разным категориям. К ним относятся продукты обмена – фенолы, флавонолы, антоцианы, алколоиды и др. Растение не способно выделять эти вещества наружу, поэтому они откладываются в вакуолях. При этом вещества недостаточно растворимые в воде, переводятся в растворимую форму. Например, антоцианидины превращаются в водорастворимые глюкозиды-антоцианы, которые придают окраску лепесткам цветков и других органов растения. Ядовитые вещества, высокотоксичные алколоиды, неприятные на вкус вещества высвобождаются из вакуолей клеток при повреждении растений травоядными животными или насекомыми и служат растению химической защитой. К другой категории относятся важные для растений ассимиляты – сахара и белки. Они сохраняются в вакуолях как запасные вещества, которые могут включаться в процессы метаболизма по мере необходимости. Сахара содержатся в вакуолях в форме растворов, а белки осаждаются, превращаются в гомогенную массу и обезвоживаются, превращаясь в алейроновые зерна, образующиеся, например, в эндосперме семян. При набухании семян происходит гидролиз белков, а аминокислоты используются для питания развивающегося зародыша.
Гидростатическое давление в вакуолях и цитоплазме примерно одинаково, они вносят совместный вклад в поддержание тургорного давления в клетке и ее осмотического равновесия.
Источник: media.ls.urfu.ru
Вакуоль в клетках растений
Вакуоль в растительной клетке окружена одной мембраной, называемой тонопластом. Она образуется, когда везикулы, высвобождаемые эндоплазматическим ретикулумом и комплексом Гольджи, сливаются вместе. Недавно развившиеся растительные клетки обычно содержат несколько небольших вакуолей. По мере созревания клетки крупная центральная вакуоль образуется из слияния меньших вакуолей. Центральная вакуоль может занимать до 90% объема клетки.
Функция вакуоли
Вакуоли в клетках растений выполняют ряд важных функций, включая:
- Тургорное давление — сила, воздействующая на клеточную стенку, так как содержимое клетки подталкивает плазматическую мембрану к стенке клетки. Вода, заполняющая центральную вакуолью, оказывает давление на клеточную стенку, чтобы помочь растительным структурам оставаться жесткими и прямыми.
- Рост — центральные вакуоли помогают в удлинении клеток, поглощая воду и оказывают давление тургора на клеточную стенку. Росту способствует высвобождение определенных белков, которые снижают жесткость клеточной стенки.
- Хранение — вакуоли хранят важные минералы, воду, питательные вещества, ионы, отходы, небольшие молекулы, ферменты и растительные пигменты.
- Деградация молекул — внутренняя кислая среда вакуолей способствует деградации более крупных молекул, направляемых в вакуоль для разрушения. Тонопласт помогает создать эту кислую среду путем переноса ионов водорода из цитоплазмы в вакуоль. Среда с низким рН активирует ферменты, которые разрушают биологические полимеры.
- Детоксикация — вакуоли удаляют потенциально токсичные вещества из цитозоля, такие как избыточные тяжелые металлы и гербициды.
- Защита — некоторые вакуоли хранят и выделяют химические вещества, которые являются ядовитыми или неприятными для защиты растений от животных.
- Прорастание семян — вакуоли являются источником питательных веществ для семян во время прорастания. Они хранят важные углеводы, белки и жиры, необходимые для роста.
Вакуоли клеток растений функционируют аналогично лизосомам в клетках животных. Лизосомы являются мембранными мешочками ферментов, которые переваривают клеточные макромолекулы. Вакуоли и лизосомы также участвуют в запрограммированной гибели клеток, которая в растениях происходит посредством процесса, называемого автолизом. Автолиз растений — это естественный процесс, при котором растительная клетка разрушается своими ферментами. В упорядоченной серии событий вакуумирующий тонопласт разрывается, высвобождая свое содержимого в цитоплазму клетки. Пищеварительные ферменты из вакуоли затем разрушают всю клетку.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info
Центральная вакуоль
Центральная вакуоль занимает более половины объема зрелых клеток, особенно в паренхиме и колленхиме. Основные функции – запас воды, накопление ионов, поддержание тургора.
Мембрана вакуоли называется тонопластом, а внутреннее содержимое — клеточным соком. Он представляет собой концентрированный раствор. Состав клеточного сока: вода, минеральные соли, сахара, танины, органические кислоты, кислород, диоксид углерода, пигменты антоцианы, продукты клеточного метаболизма и др.
Тонопласт избирательно проницаем. Через него в вакуоль поступает вода. Возникает тургорное давление, и цитоплазма прижимается к клеточной стенке. За счет такого осмотического поглощения воды клетка растягивается во время роста.
Ряд содержащихся в клеточном соке антоцианов придают разноцветную окраску цветкам и плодам. Так вакуоль выполняет функцию привлечения насекомых-опылителей и животных-распространителей плодов и семян. Совместно с каратиноидами антоцианы придают окраску осенней листве, когда разрушается хлорофилл.
Центральная вакуоль может содержать гидролитические ферменты, что позволяет ей выполнять функцию лизосом. После гибели клетки ферменты попадают в цитоплазму, и происходит автолиз.
В вакуолях накапливаются такие отходы жизнедеятельности как кристаллы оксалата кальция. Среди вторичных продуктов метаболизма — алкалоиды, которые предположительно выполняют защитную функцию наряду с танинами, препятствуя поеданию животными.
У некоторых растений в клеточном соке накапливается млечный сок, представляющий собой беловатую эмульсию. У ряда растений есть клетки, которые его экскретируют.
В центральных вакуолях также запасаются питательные вещества (сахароза, инулин), которые используются при необходимости, как и содержащиеся здесь минеральные соли.
Источник: biology.su
ЦЕНТРАЛЬНАЯ ВАКУОЛЬ РАСТИТЕЛЬНОЙ КЛЕТКИ.
I. ВВЕДЕНИЕ
Современный этап развития биологии характеризуется исследованиями в ряде приоритетных научных направлений, одним из которых является изучение механизмов мембранного транспорта. Менее всего в настоящее время изучены эти механизмы применительно к специфичной для растений мембране — вакуолярной.
Наличие вакуоли является отличительной особенностью растительной клетки. Её роль в жизнедеятельности растений достаточно велика. В настоящее время показано, что это полифункциональный компартмент, который занимает особое место в структурной и функциональной организации растительного организма. Трудно переоценить роль вакуоли в регуляции клеточного объёма, тургора, а также pH и ионного гомеостаза цитозоля, что обеспечивает работу ферментов цитоплазмы, в изоляции вторичных продуктов метаболизма и метаболических ядов, в трансдукции сигналов различной природы, в процессах апоптоза, в ответных защитных реакциях растения на стрессовое воздействие и т.д. Одна из важнейших функций вакуоли состоит в запасании метаболитов.
Основным метаболитом, запасаемым в растении (в отличие от глюкозы в животной клетке), является сахароза. Процессы синтеза и транспортировки сахарозы по растению в настоящее время изучаются.
каким бы способом ни транспортировалась сахароза (по симпласту или по апопласту), конечный этап дальнего транспорта — вакуолярная мембрана, которая содержит переносчик сахарозы и протонные помпы, обеспечивающие процесс транспорта. Путь к тонопласту идёт по донорно-акцепторным градиентам, а скорость закачивания сахарозы в вакуоль, которая зависит от активности протонных помп тонопласта, может являться в этом процессе одним из определяющих звеньев.
Различают пищеварительные и сократительные (пульсирующие) вакуоли, регулирующие
осмотическое давление и служащие для выведения из организма продуктов распада.
В клетках животных – пиноцитозная вакуоль.
Внутренняя мембрана клетки – тонопласт — окружает вакуоль. Царство одноклеточные, тип простейшие. – группа животных большинство из которой питаются органической пищей (бактерии, одноклеточные водоросли). Захваченные ими оформленные частицы пищи перевариваются в пищеварительных вакуолях – органоидах пищеварения. После чего растворенная часть переваренной пищи поступает в цитоплазму. У простейших, поглощающих
из окружающей среды жидкие органические вещества, пищеварительные вакуоли,
как правило, отсутствуют.
Так же для простейших характерны сократительные вакуоли – органоиды, основная функция которых заключается в регуляции осмотического давления в клетке. Эти вакуоли свойственны преимущественно пресноводным простейшим, так как в их теле осмотическое давление благодаря образованию солей, всегда выше, чем в окружающей среде, в результате чего через покровы внутрь тела непрерывно поступает вола. Удаление избытка воды с помощью сократительных вакуолей предохраняет простейших от гибели. Вместе с водой через сократительные вакуоли из тела простейших частично выводятся продукты обмена веществ.
Для клеток растений характерна хорошо развитая система вакуолей, в значительной степени обеспечивающая их соматические свойства.
Почти во всех клетках, особенно в старых, хорошо заметны полости – вакуоли. Они заполнены клеточным соком.
Клеточный сок – это вода с растворенными в ней сахарами и другими органическими веществами.
клеточном соке могут содержаться красящие вещества, придающие синюю, фиолетовую, малиновую окраску лепесткам и другим органам растений. Осенняя окраска листьев также обусловлена окрашенным клеточным соком. Разрезая спелый плод или другую сочную часть растения, мы повреждаем клетки, и из их вакуолей вытекает сок. Цитоплазма в каждой из клеток оттеснена вакуолью к оболочке. Молодые клетки, в отличии от старых, неспособных делиться, содержат много вакуолей. В старой клетке обычно имеется одна большая вакуоль.
II. ОСНОВНОЕ СОДЕРЖАНИЕ.
Р астительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал.
астительные клетки отличаются наличием толстой целлюлозной клеточной стенки, пластид, крупной центральной вакуоли, смещающей ядро к периферии. Клеточный центр высших растений не содержит центриоли. Запасным углеводом является крахмал.
Вакуоль (франц. vacuole, от лат. vacuus — пустой) – наполненный жидкостью мембранный мешочек. В животных клетках могут наблюдаться небольшие вакуоли, выполняющие фагоцитарную, пищеварительную, сократительную и другие функции. Растительные клетки имеют одну большую центральную вакуоль. Жидкость, заполняющая её, называется клеточным соком. Это концентрированный раствор сахаров, минеральных солей, органических кислот, пигментов и других веществ. Вакуоли накапливают воду, могут содержать красящие пигменты, защитные вещества (например, таннины), гидролитические ферменты, вызывающие автолиз клетки, отходы жизнедеятельности, запасные питательные вещества. У растений вакуоли — производные эндоплазматической сети, окружены полупроницаемой мембраной — тонопластом. Вся система вакуолей растительной клетки называется вакуомом, который в молодой клетке представлен системой канальцев и пузырьков; по мере роста и дифференцировки клетки они увеличиваются и сливаются в одну большую центральную вакуолю, занимающую 70- 95% объема зрелой клетки.
Вакуоли растений, строение и функции.
В
очень молодых клетках вакуолей нет или они почти незаметны по мере роста и дифференцировки клетки вакуоли появляются в разных её участках, а затем, постепенно увеличиваясь, сливаются друг с другом и образуют одну или несколько крупных вакуолей, занимающих до 80% объема всей клетки.
Центральные вакуоли отделены от цитоплазмы одинарной липоидно-белковой полупроницаемой мембраной, сходной по толщине с плазмалеммой. Мембрана, ограничивающая центральные вакуоли, носит название тонопласта. Возникают центральные вакуоли из мелких пузырьков, отщепившихся от эндоплазматической сети. Такие первичные провакуоли растут в объеме, сливаются друг с другом и, в конце концов, образуют одну или несколько крупных вакуолей, оттесняющих цитоплазму с ядром и органоидами к периферии клетки.
Полость вакуоли заполнена так называемым клеточным соком, представляющим собой водный раствор, в который входят различные неорганические соли, сахара, органические кислоты и их соли и другие низкомолекулярные соединения, а также некоторые высокомолекулярные вещества (например, белки).
Одной из главных ее функций является поддержание тургорного давления клеток. Растворенные в соке вакуолей молекулы определяют его осмотическую концентрацию. Соответствующая молярная концентрация сока вакуолей и полупроницаемые свойства как ее мембраны, тонопласта, так и плазмалеммы способствуют тому, что вакуоль функционирует в
качестве осмометра и придает клетке необходимую прочность и тургисцентность (напряженность).
Другая функция определяется тем, что вакуоль представляет собой большую полость, отделенную от метаболизирующей гиалоплазмы мембраной, тонопластом, обладающим свойствами полупроницаемости и через который может происходить, как и через плазматическую мембрану, активный транспорт различных молекул. Поэтому вакуоли могут использоваться клетками как накопительные резервуары не только для отложения запасных веществ, но и для выброса метаболитов, для экскреции. Так выводятся, секретируются из клетки все водорастворимые метаболиты. Нерастворимые в воде органические компоненты могут превращаться в растворимые глюкозиды, соединяясь с молекулами сахаров. В вакуолях происходит отложение многих глюкозидов, к которым относятся различные пигменты, например антоцианы. Из неорганических веществ в вакуолярном соке накапливаются фосфаты калия, натрия, кальция, могут накапливаться соли органических кислот (оксалаты, цитраты и др.). Это придает вакуолярному соку отчетливую кислую реакцию (рН от 2 до 5).
Другой обширный ряд функций вакуолей связан с накоплением запасных веществ, таких, как сахара и белки. Сахара в вакуолях содержатся в виде растворов, встречаются и резервные полисахариды типа инулина. В вакуолях происходит запасание белков, что характерно для семян. Поступление белков в вакуоли связано со способностью вакуолей ЭР и АГ сливаться с тонопластом. Запасание белков семян происходит в так называемых алейроновых вакуолях, которые заполняются альбуминами и глобулинами, после чего вакуоли обезвоживаются, превращаясь в твердые алейроновые зерна. При прорастании семян эти зерна обводняются и снова превращаются в вакуоли. В таких новообразованных вакуолях выявляется активность некоторых ферментов, протеазы и РНКазы. Следовательно, алейроновые вакуоли отчасти напоминают лизосомы, где происходит переваривание запасных белков при прорастании семян. Стенки вакуолей могут образовывать впячивания внутрь, затем они отщепляются от тонопласта и растворяются внутри вакуоли.
Вакуолярная мембрана и роль вакуоли в растительной клетке.
Отличительной особенностью растительной клетки является наличие в ней центральной вакуоли, которая в ряде случаев может занимать до 90% объема зрелой растительной клетки и играет особую роль в структурной и функциональной организации растительного организма. Вакуоли активно участвуют во внутриклеточном перераспределении веществ, а также выполняют функции лизосом. Вакуоли аккумулируют большие количества запасных веществ и
других метаболитов, составляющих пищевую и лекарственную ценность растений (сахара, аминокислоты, органические кислоты, витамины, белки и др.).
При отложении веществ в запас вакуолярная мембрана выступает в качестве конечного барьера, регулирующего процессы накопления и удержания питательных веществ в запасающих органах растений. Механизмы формирования вакуолей при дифференциации клеток подробно рассматривались Матилем, который показал, что центральную вакуоль нужно рассматривать как скомбинированную органеллу, образованную путём объединения и слияния мембран различных клеточных органелл. Она является конечным продуктом дифференциации эндомембранной системы растений.
Образование центральной вакуоли стало решающем условием и основой выживания при выходе растений из воды на сушу. Формирование центральной вакуоли со специализированной мембраной — тонопластом, имеет фундаментальное значение в эволюции сосудистых растений и, видимо, является генетически закреплённым способом поддержания гомеостаза целого растения. То, что центральные вакуоли имеются практически во всех растительных организмах, свидетельствует о фундаментальной значимости этих органелл, поскольку в противном случае они бы элиминировали в процессе эволюции, по крайней мере, у некоторых видов или в некоторых тканях. Возникновение вакуоли способствовало ускорению эволюции растений и, в свою очередь, привело к изменению функциональной нагрузки и дифференциации вакуолей и их мембран в связи со специализацией клеток в системе целого растения.
В выполнении указанных функций ключевая роль принадлежит уникальной клеточной мембране — тонопласту. На сегодня известно, что вакуолярная мембрана содержит ряд специализированных систем пассивного и активного переноса веществ: каналы, переносчики, протонные помпы. Основной вклад в транспорт на тонопласте вносят две протонные помпы: Н^-АТФаза и ЕГ-пирофосфатаза. Оба фермента способны преобразовывать освобождённую при гидролизе АТФ и неорганического пирофосфата энергию в перенос протонов через тонопласт. Генерируемая разность электрохимического потенциала расходуется на вторичный транспорт ионов, Сахаров, аминокислот, который осуществляется через каналы и переносчики. Таким образом, Н^-АТФаза и К^-пирофосфтаза являются ключевыми ферментами в системе переноса углеводов и других соединений в антипорте с протоном.
Функции вакуоли в клетках высших растений.
В настоящее время убедительно доказано, что вакуоль является полифункциональным компартментом растительных клеток. Она играет основную роль в запасании метаболитов, регуляции клеточного объёма, тургора, а также pH и ионного гомеостаза цитозоля, что обеспечивает работу ферментов цитоплазмы. Вакуоль изолирует вторичные продукты метаболизма и метаболические яды, в ней происходят процессы детоксикации и катаболической деградации целого ряда соединений эндогенного и экзогенного происхождения, переваривание «стареющих» или повреждённых органелл.
В последние годы выявлена важная роль вакуоли в трансдукции сигналов различной природы, в процессах апоптоза и в ответных защитных реакциях растений на стрессовое воздействие. Это перечисление наиболее значимых функций вакуоли скорее всего не является исчерпывающим.
Трудно сказать, какая из этих функций является наиболее важной для растения. Одной из первых была сформулирована концепция о функции растительных вакуолей как о литическом компартменте, который, по аналогии с лизосомами животных клеток, переваривает «стареющие» или повреждённые органеллы. Кислые гидролазы были обнаружены в вакуолях, изолированных из всех изученных растительных объектов. Многие клеточные белки, имеющие дефект в своей структуре, способны накапливаться внутри вакуоли и там претерпевать протеолитическую деградацию под действием вакуолярных гидролаз. Интенсивная деградация белков внутри вакуоли свойственна также процессу вакуолярной аутофагии растительных клеток, который происходит в условиях сахарозного или углеводного голодания, когда содержание Сахаров и крахмала в клетках, а также их метаболическая активность заметно снижаются. Установлено, что вакуолярная аутофагия свойственна также стареющим клеткам растений. Из-за наличия внутриклеточных пулов углеводов и способности контролировать процесс аутофагии растительные клетки в условиях голодания могут некоторое время сохранять свою жизнеспособность. Литическая функция вакуолей тесно связана с апоптозом, который, как было показано в последние годы, является важным фактором развития и дифференциации многоклеточного растительного организма. В частности, он имеет место в клетках листьев как ответная, защитная реакция на действие патогена или во время старения таких клеток, способствуя транслокации тех или иных компонентов к другим, более молодым, растущим растительным клеткам.
Доминирующая роль центральной вакуоли в регуляции внутриклеточного осмотического давления и клеточного тургора доказана результатами многочисленных исследований. Она
находится в соответствии с интенсивным накоплением внутри вакуоли неорганических ионов, а также целого ряда других соединений, в том числе органических. Осмотический баланс в клетке может достигаться благодаря аккумуляции внутри вакуолярного и цитозольного компартментов осмолитов различной природы, что особенно важно в условиях осмотического стресса, когда в цитоплазме накапливаются осмолиты, способные защитить ферменты и другие биополимеры от инактивации в результате обезвоживания или сопутствующего ему окислительного стресса. Осморегуляторная функция вакуоли не ограничивается участием в регуляции клеточного тургора и, по-видимому, связана с включением в регуляцию объёма или оводнённости цитозоля растительных клеток. Это предположение подтверждается обнаружением на вакуолярной мембране аквапоринов -интегральных белков, облегчающих пассивный трансмембранный перенос молекул воды.
Одной из основных функций вакуоли является гомеостатическая регуляция ионного состава цитозоля растительных клеток, связанная с поддержанием оптимальных условий для активной работы ферментов цитоплазмы. Её потенциальная способность к выполнению этой важной функции в клетке обусловлена тем, что она может накапливать внутри себя значительные концентрации как основных физиологически важных ионов, таких как Н4″, К+, Na+, Ca2+, Mg2+, СГ, N03″, так и ионов, обладающих токсическим действием (Cd , Zn ). Недавно была убедительно продемонстрирована гомеостатическая регуляция уровня К+ в цитозоле растительных клеток за счёт его вакуолярной компартментации. Включение вакуоли в регуляцию уровня свободного Са2+ также является хорошо известным примером участия этой органеллы во внутриклеточном ионном гомеостазе растительных клеток. Регуляция уровня Са2+ в цитозоле с участием вакуоли включает в себя не только аккумуляцию Са2+ внутри этой органеллы, но и мобилизацию его из вакуоли в цитозоль. Экспорт Са из вакуоли выполняет также очень важную роль в трансдукции сигналов в растительной клетке. В этом случае вакуоль выступает в роли донора вторичных посредников, обусловливающих передачу сигналов к метаболическим системам клетки.
Одна из важных функций вакуоли состоит в изоляции вторичных продуктов метаболизма и метаболических ядов. К настоящему времени достигнут значительный прогресс в изучении обусловленных активностью вакуоли механизмов детоксикации экзогенных токсинов растительными клетками. Решающую роль в этом сыграло недавно обнаруженная способность изолированных вакуолей к Mg-АТФ-зависимой аккумуляции разнообразных глютатион-S-коньюгатов, в том числе и с гербицидами. Экспорт из цитозоля в вакуоль соответствующих GS-коньюгатов предотвращает токсическое действие на клетку тех или иных ксенобиотиков. В настоящее время найдено, что через GS-коньюгат-транслоцирующую систему могут
переноситься GS-коньюгаты антоцианина, некоторые фенольные соединения с аллелопатическими свойствами, а также некоторые фитоалексины. В животных клетках через эту транспортную систему могут изолироваться соединения, образуемые в ходе перекисного окисления липидов клеточных мембран или окислительного повреждения ДНК под действием активных форм кислорода. Поэтому есть основание предполагать, что эти же функции будут обнаружены и в растительных клетках.
Одной из наиболее важных функций вакуоли является запасающая. В некоторых случаях в вакуолях накапливаются значительные количества аминокислот и водорастворимых углеводов, причём их содержание нередко достигает 70-80% от суммы всех аминокислот или углеводов клетки. Нередко в вакуолях обнаруживают значительный пул органических кислот. Так, на листьях Bryophylum показано, что всего 2% всей внутриклеточной изолимонной кислоты участвует в метаболизме, остальные 98% изолированы в вакуолях. Состав углеводов, накапливаемых в большом количестве в вакуолях, значительно различается. Если вакуоли из лепестков и листьев тюльпана содержат в основном глюкозу и фруктозу, то корнеплоды столовой свёклы накапливают сахарозу. В вакуолях накапливаются большие количества фенолов, нитратов, витаминов и других вторичных соединений, которые нередко являются антиоксидантами или относятся к лекарственным соединениям.
В выполнении всех этих функций важная роль принадлежит уникальной клеточной мембране — тонопласту, которая разделяет содержимое вакуоли и цитоплазму клетки.
1.2. Тонопласт: особенности химического состава и физиологическая роль белков
Изолированная вакуолярная мембрана обладает типичным трёхслойным строением при толщине 9,5-10 нм, присущим для пограничных мембран in suti, и характеризуется достаточно сложной надмолекулярной структурой. При этом тонопласт имеет чёткую асимметрию, которая выражается в различной электронной плотности периферических слоев мембраны на поперечных срезах и в неодинаковой концентрации внутримембранных глобулярных частиц на вакуолярных и цитоплазматических сколах. Коэффициент распределения глобулярных частиц между вакуолярной и цитоплазматической сторонами составлял 725/1006 = 0.361. Известно, что высокая насыщенность глобулярными частицами (до 1000 — 3500 частиц на 1мкм2) характерна для мембран, проявляющих особенно высокую функциональную активность.
Внутримембранные глобулы, как правило, представляют собой глобулы трансмембранных (интегральных) белков, которые могут являться ферментами, ионными каналами и переносчиками. Суммарная площадь, занятая частицами на продольных сколах, составляла около 23,5% от общей площади поверхности тонопласта, а объём, занимаемый глобулярными частицами, составлял около 22% от объёма мембраны. Это удовлетворительно
коррелирует с результатами прямых биохимических анализов тонопласта, согласно которым во фракции вакуолярных мембран, тщательно очищенных от периферических белков, около 20% приходится на долю интегральных белков, извлекаемых из мембран только с помощью детергентов. Белок / липидное соотношение в тонопласте столовой свёклы составляло 0,69. 6-8% белков тонопласта были отнесены к периферическим, остальные 92% -к интегральным, пронизывающим мембранный матрикс. При электрофоретическом разделении в ПААГ с ДС в спектре белков тонопласта выявлялось около 20 полипетидов, большинство из которых имели молекулярные массы менее 70 кД. Спектр белков тонопласта, солюбилизированных Тритоном Х-100, состоял из 15 белковых полос, 8 из которых были гликопротеинами.
Такие биохимические характеристики, как белок/липидное соотношение, количественный и качественный спектр белков изменялись при использовании других объектов для получения вакуолярных мембран. Так, например, во фракции тонопласта, выделенной из корнеплодов столовой свёклы и из лепестков амариллиса обнаружены лишь следы углеводов, тогда как в тонопласте, выделенном из дрожжей, на 100 мг белка приходилось 79 мг углеводов. Но все исследователи, независимо от объекта исследования, отмечают высокую текучесть и эластичность вакуолярной мембраны, которая обусловлена большим содержанием липидов, составляющих до 80% от её общего веса.
III. ЛИТЕРАТУРА.
1. К.М.Суханова «Цитология» Москва,1970г.
2. Л.Я.Кулинич «Справочник по биологии» Москва,1986г.
3. Б.М.Медников «Аксиомы биологии» Москва,1985г.
4. В. Азерников. Тайнопись жизни. Москва, 1973г.
5. Н. Н. Воронцов, Л. Н. Сухорукова. Эволюция органического мира. Москва,1991г.
6. Э. Рис, М. Стернберг. От клеток к атомам. Москва, 1988г.
7. А. С. Трошин, А. Д. Браун, Ю. Б. Вахтин, Л. Н. Жинкин, С. Штрбанова. Кто мы? Книга о жизни, клетках и ученых. Москва, 1984г
8. Руководство по цитологии, т. 1, М.—Л., 1965, гл. 2;
9. Робертис Э. де, Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967;
10. Робертсон Дж., Мембрана живой клетки, в сборнике: Структура и функция клетки, пер. с англ., М., 1964; Finean J. В., The molecular organization of cell membranes, «Progress in Biophysics and Molecular Biology», 1966, v. 16, p. 143—70.
11. http://www.zubreshka.ru/?id=27090&from=350
12. http://www.college.ru/biology/course/content/chapter9/section1/paragraph5/theory.html
13. http://www.medkurs.ru/lecture1k/med_biology/qm24/2480.html
14. http://all-referats.ru/alls/10/6497.html
15. http://fundamed.narod.ru/Shpori/Cytology_3.doc
Источник: infourok.ru