Являются обязательными веществами плазматической мембраны
Плазматическая мембрана
Лекция
Плазматическая мембрана
План
1.Строение плазматической мембраны
2.Функции плазматической мембраны. Механизмы транспорта веществ через плазмолемму. Рецепторная функция плазмалеммы
.Межклеточные контакты
1.Строение плазматической мембраны
Плазматическая мембрана, или плазмалемма, представляет собой поверхностную периферическую структуру, ограничивающую клетку снаружи и обеспечивающую ее связь с другими клетками и внеклеточной средой. Она имеет толщину около 10 нм. Среди других клеточных мембран плазмалемма является самой толстой. В химическом отношении плазматическая мембрана представляет собой липопротеиновый комплекс. Основными компонентами являются липиды (около 40%), белки (более 60%) и углеводы (около 2-10%).
К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофильность). Характерными представителями липидов, встречающимися в плазматической мембране, являются фосфолипиды, сфингомиелины и холестерин. В растительных клетках холестерин замещается фитостерином. По биологической роли белки плазмалеммы можно разделить на белки-ферменты, рецепторные и структурные белки. Углеводы плазмалеммы входят в состав плазмалеммы в связанном состоянии (гликолипиды и гликопротеины).
В настоящее время общепринятой является жидкостно-мозаичная модель строения биологической мембраны. Согласно этой модели структурную основу мембраны образует двойной слой фосфолипидов, инкрустированный белками. Хвосты молекул обращены в двойном слое друг к другу, а полярные головки остаются снаружи, образуя гидрофильные поверхности. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь на разную глубину (есть периферические белки, часть белков пронизывает мембрану насквозь, часть погружена в слой липидов). Большинство белков не связаны с липидами мембраны, т.е. они как бы плавают в «липидном озере». Поэтому молекулы белков способны перемещаться вдоль мембраны, собираться в группы или, наоборот, рассеиваться на поверхности мембраны. Это говорит о том, что плазматическая мембрана не является статичным, застывшим образованием.
Снаружи от плазмолеммы располагается надмембранный слой — гликокаликс. Толщина этого слоя составляет около 3-4 нм. Гликокаликс обнаружен практически у всех животных клеток. Он представляет собой связанный с плазмолеммой гликопротеиновый комплекс. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами плазматической мембраны. В гликокаликсе могут располагаться белки-ферменты, участвующие во внеклеточном расщеплении различных веществ. Продукты ферментативной активности (аминокислоты, нуклеотиды, жирные кислоты и др.) транспортируются через плазматическую мембрану и усваиваются клетками.
Плазматическая мембрана постоянно обновляется. Это происходит путем отшнуровывания мелких пузырьков с ее поверхности внутрь клетки и встраивания в мембрану вакуолей, поступивших изнутри клетки. Таким образом, в клетке постоянно происходит поток мембранных элементов: от плазматической мембраны внутрь цитоплазмы (эндоцитоз) и поток мембранных структур из цитоплазмы к поверхности клетки (экзоцитоз). В круговороте мембран ведущая роль отводится системе мембранных вакуолей комплекса Гольджи.
2. Функции плазматической мембраны. Механизмы транспорта веществ через плазмолемму. Рецепторная функция плазмалеммы
Плазматическая мембрана выполняет ряд важнейших функций:
1)Барьерная. Барьерная функция плазматической мембраны заключается в ограничении свободной диффузии веществ из клетки в клетку, предотвращении утечки водорастворимого содержимого клетки. Но поскольку клетка должна получать необходимые питательные вещества, выделять конечные продукты метаболизма, регулировать внутриклеточные концентрации ионов, то в ней образовались специальные механизмы переноса веществ через клеточную мембрану.
2)Транспортная. К транспортной функции относится обеспечение поступления и выведения различных веществ в клетку и из клетки. Важное свойство мембраны — избирательная проницаемость, или полупроницаемость. Она легко пропускает воду и водорастворимые газы и отталкивает полярные молекулы, такие как глюкоза или аминокислоты.
Существует несколько механизмов транспорта веществ через мембрану:
пассивный транспорт;
активный транспорт;
транспорт в мембранной упаковке.
Перенос полярных молекул (сахаров, аминокислот), осуществляемый с помощью специальных мембранных транспортных белков называется облегченной диффузией.
i>Такие белки обнаружены во всех типах биологических мембран, и каждый конкретный белок предназначен для переноса молекул определенного класса. Транспортные белки являются трансмембранными, их полипептидная цепь пересекает липидный бислой несколько раз, формируя в нем сквозные проходы. Это обеспечивает перенос специфических веществ через мембрану без непосредственного контакта с ней. Существует два основных класса транспортных белков: белки-переносчики (транспортеры) и каналообразующие белки (белки-каналы). Белки-переносчики переносят молекулы через мембрану, предварительно изменяя их конфигурацию. Каналообразующие белки формируют в мембране заполненные водой поры. Когда поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них. Если молекула транспортируемого вещества не имеет заряда, то направление транспорта определяется градиентом концентрации. Если молекула заряжена, то на ее транспорт, кроме градиента концентрации, влияет и электрический заряд мембраны (мембранный потенциал). Внутренняя сторона плазмалеммы обычно заряжена отрицательно по отношению к наружной. Мембранный потенциал облегчает проникновение в клетку положительно заряженных ионов и препятствует прохождению ионов заряженных отрицательно.
Активный транспорт. Активным транспортом называется перенос веществ против электрохимического градиента.
всегда осуществляется белками-транспортерами и тесно связан с источником энергии. В белках-переносчиках имеются участки связывания с транспортируемым веществом. Чем больше таких участков связывается с веществом, тем выше скорость транспорта. Селективный перенос одного вещества называется унипортом. Перенос нескольких веществ осуществляют котранспортные системы. Если перенос идет в одном направлении — это симпорт, если в противоположных — антипорт. Так, например, глюкоза из внеклеточной жидкости в клетку переносится унипортно. Перенос же глюкозы и Na4 из полости кишечника или канальцев почек соответственно в клетки кишечника или кровь осуществляется симпортно, а перенос С1~ и НСО’ антипортно. Предполагается, что при переносе возникают обратимые конформационные изменения в транспортере, что и позволяет премещать соединенные с ним вещества.
Примером белка-переносчика, использующего для транспорта веществ энергию выделившуюся при гидролизе АТФ, является Na+-К+ насос, обнаруженный в плазматической мембране всех клеток. Na+-K насос работает по принципу антипорта, перекачивая Na" из клетки и Кт внутрь клетки против их электрохимических градиентов. Градиент Na+ создает осмотическое давление, поддерживает клеточный объем и обеспечивает транспорт сахаров и аминокислот.
работу этого насоса тратится треть всей энергии необходимой для жизнедеятельности клеток. При изучении механизма действия Na+-K+ насоса было установлено, что он является ферментом АТФазой и трансмембранным интегральным белком. В присутствии Na+ и АТФ под действием АТФа-зы от АТФ отделяется концевой фосфат и присоединяется к остатку аспарагиновой кислоты на молекуле АТФазы. Молекула АТФазы фосфорилируется, изменяет свою конфигурацию и Na+ выводится из клетки. Вслед за выведением Na из клетки всегда происходит транспорт К’ в клетку. Для этого от АТФазы в присутствии К отщепляется ранее присоединенный фосфат. Фермент дефосфорилируется, восстанавливает свою конфигурацию и К1 "закачивается" в клетку.
АТФаза образована двумя субъединицами, большой и малой. Большая субъединица состоит из тысячи аминокислотных остатков, пересекающих бислой несколько раз. Она обладает каталитической активностью и способна обратимо фосфорилироваться и дефосфорилироваться. Большая субъединица на цитоплазматической стороне имеет участки для связывания Na+ и АТФ, а на внешней стороне -участки для связывания К+ и уабаина. Малая субъединица является гликопротеином и функция его пока не известна.
Na+-K насос обладает электрогенным эффектом. Он удаляет три положительно заряженных иона Naf из клетки и вносит в нее два иона К В результате через мембрану течет ток, образующий электрический потенциал с отрицательным значением во внутренней части клетки по отношению к ее наружной поверхности. Na"-K+ насос регулирует клеточный объем, контролирует концентрацию веществ внутри клетки, поддерживает осмотическое давление, участвует в создании мембранного потенциала.
Транспорт в мембранной упаковке. Перенос через мембрану макромолекул (белков, нуклеиновых кислот, полисахаридов, липопротеидов) и других частиц осуществляется посредством последовательного образования и слияния окруженных мембраной пузырьков (везикул). Процесс везикулярного транспорта проходит в две стадии. Вначале мембрана пузырька и плазмалемма слипаются, а затем сливаются. Для протекания 2 стадии необходимо чтобы молекулы воды были вытеснены взаимодействующими липидными бислоями, которые сближаются до расстояния 1-5 нм. Считается, что данный процесс активизируют специальные белки слияния (они выделены пока только у вирусов). Везикулярный транспорт имеет важную особенность — поглощенные или секретируемые макромолекулы, находящиеся в пузырьках, обычно не смешиваются с другими макромолекулами или органеллами клетки. Пузырьки могут сливаться со специфическими мембранами, что и обеспечивает обмен макромолекулами между внеклеточным пространством и содержимым клетки. Аналогично происходит перенос макромолекул из одного компартмента клетки в другой.
Транспорт макромолекул и частиц в клетку называется эндоцитозом. При этом транспортируемые вещества обволакиваются частью плазматической мембраны, образуется пузырек (вакуоль), который перемещается внутрь клетки. В зависимости от размера образующихся пузырьков различают два вида эндоцитоза — пиноцитоз и фагоцитоз.
Пиноцитоз обеспечивает поглощение жидкости и растворенных веществ в виде небольших пузырьков (d=150 нм). Фагоцитоз — это поглощение больших частиц, микроорганизов или обломков органелл, клеток. При этом образуются крупные пузырьки, фагосомы или вакуоли (d-250 нм и более). У простейших фагоцитарная функция — форма питания. У млекопитающих фагоцитарная функция осуществляется макрофагами и нейтрофилами, защищающими организм от инфекции путем поглощения вторгшихся микробов. Макрофаги участвуют также в утилизации старых или поврежденных клеток и их обломков (в организме человека макрофаги ежедневно поглощают более 100 старых эритроцитов). Фагоцитоз начинается только тогда, когда поглощаемая частица свяжется с поверхностью фагоцита и активирует специализированные рецепторные клетки. Связывание частиц со специфическими рецепторами мембраны вызывает образование псевдоподии, которые обволакивают частицу и, сливаясь краями, образуют пузырек —фагосому. Образование фагосомы и собственно фагоцитоз происходит лишь в том случае, если в процессе обволакивания частица постоянно контактирует с рецепторами плазмалеммы, как бы "застегивая молнию".
Значительная часть материала, поглощенного клеткой путем эндоцитоза, заканчивает свой путь в лизосомах. Большие частицы включаются в фагосомы, которые затем сливаются с лизосомами и образуют фаголизосомы. Жидкость и макромолекулы, поглощенные при пиноцитозе, первоначально переносятся в эндосомы, которые также сливаются с лизосомами, образуя эндолизосомы. Присутствующие в лизосомах разнообразные гидролитические ферменты быстро разрушают макромолекулы. Продукты гидролиза (аминокислоты, сахара, нуклеотиды) транспортируются из лизосом в цитозоль, где используются клеткой. Большинство мембранных компонентов эндоцитозных пузырьков из фагосом и эндосом возвращаются с помощью экзоцитоза к плазматической мембране и там повторно утилизируются. Основным биологическим значением эндоцитоза является получение строительных блоков за счет внутриклеточного переваривания макромолекул в лизосомах.
Поглощение веществ в эукариотических клетках начинается в специализированных областях плазматической мембраны, так называемых окаймленных ямках. На электронных микрофотографиях ямки выглядят как впячивания плазматической мембраны, цитоплазматическая сторона которых покрыта волокнистым слоем. Слой как бы окаймляет небольшие ямки плазмалеммы. Ямки занимают около 2% общей поверхности клеточной мебраны эукариотов. В течении минуты ямки растут, все глубже впячиваются, втягиваются в клетку и затем, сужаясь у основания, отщепляются, образуя окаймленные пузырьки. Установлено, что из плазматической мембраны фибробластов в течении одной минуты отщепляется примерно четвертая часть мембраны в виде окаймленных пузырьков. Пузырьки быстро теряют свою кайму и приобретают способность сливаться с лизосомой.
Эндоцитоз может быть неспецифическим (конститутивным) и специфическим (рецепторным). При неспецифическом эндоцитозе клетка захватывает и поглощает совершенно чуждые ей вещества, например, частицы сажи, красители. Вначале происходит осаждение частиц на гликокаликсе плазмалеммы. Особенно хорошо осаждаются (адсорбируются) положительно заряженные группы белков, так как гликокаликс несет отрицательный заряд. Затем изменяется морфология клеточной мембраны. Она может либо погружаться, образуя впячивания (инвагинации), либо, наоборот, формировать выросты, которые как бы складываются, отделяя небольшие объемы жидкой среды. Образование инвагинаций более характерно для клеток кишечного эпителия, амеб, а выростов — для фагоцитов и фибробластов. Заблокировать эти процессы можно ингибиторами дыхания. Образовавшиеся пузырьки — первичные эндосомы, могут сливаться между собой, увеличиваясь в размере. В дальнейшем они соединяются с лизосомами, превращаясь в эндолизосому — пищеварительную вакуоль. Интенсивность жидкофазного неспецифического пиноцитоза довольно высока. Макрофаги образуют до 125, а клетки эпителия тонкого кишечника до тысячи пиносом в минуту. Обилие пиносом приводит к тому, что плазмалемма быстро тратится на образование множества мелких вакуолей. Восстановление мембраны идет довольно быстро при рециклизации в процессе экзоцитоза за счет возвращения вакуолей и их встраивания в плазмалемму. У макрофагов вся плазматическая мембрана замещается за 30 минут, а у фибробластов за 2 часа.
Более эффективным способом поглощения из внеклеточной жидкости специфических макромолекул является специфический эндоцитоз (опосредуемый рецепторами). Макромолекулы при этом связываются с комплементарными рецепторами на поверхности клетки, накапливаются в окаймленной ямке, и затем, образуя эндосому, погружаются в цитозоль. Рецепторный эндоцитоз обеспечивает накопление специфических макромолекул у своего рецептора. Молекулы, которые связываются на поверхности плазмалеммы с рецептором, называются лигандами. При помощи рецепторного эндоцитоза во многих животных клетках идет поглощение холестерина из внеклеточной среды.
Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае вакуоли подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются и содержимое вакуоли поступает в окружающую среду. У некоторых простейших места на клеточной мембране для экзоцитоза заранее предопределены. Так, в плазматической мембране некоторых ресничных инфузорий есть определенные участки с правильным расположением крупных глобул интегральных белков. У мукоцист и трихоцист инфузорий полностью готовых к секреции, на верхней части плазмалеммы имеется венчик из глобул интегральных белков. Этими участками мембраны мукоцист и трихоцист соприкасаются с поверхностью клетки. Своеобразный экзоцитоз наблюдается в нейтрофилах. Они способны при определенных условиях выбрасывать в окружающую среду свои лизосомы. При этом в одних случаях образуются небольшие выросты плазмалеммы, содержащие лизосомы, которые затем отрываются и переходят в среду. В других случаях наблюдается инвагинация плазмалеммы вглубь клетки и захват ею лизосом, распложенных далеко от поверхности клетки.
Процессы эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы.
Рецепторная функция плазмалеммы. Это одна из главных, универсальных для всех клеток, является рецепторная функция плазмалеммы. Она определяет взаимодействие клеток друг с другом и с внешней средой..
Все многообразие информационных межклеточных взаимодействий схематически можно представить как цепь последовательных реакций сигнал-рецептор-вторичный посредник-ответ (концепция сигнал-ответ). Передачу информации от клетки к клетке осуществляют сигнальные молекулы, которые вырабатываются в одних клетках и специфически влияют на другие, чувствительные к сигналу (клетки-мишени). Сигнальная молекула — первичный посредник связывается с находящимися на клетках-мишенях рецепторами, реагирующими только на определенные сигналы. Сигнальные молекулы — лиганды — подходят к своему рецептору как ключ к замку. Лигандами для мембранных рецепторов (рецепторов плазмалеммы) являются гидрофильные молекулы, пептидные гормоны, нейромедиаторы, цитокины, антитела, а для ядерных рецепторов — жирорастворимые молекулы, стероидные и тиреоидные гормоны, витамин Д В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса — полисахариды и гликопротеиды. Считается, что чувствительные к отдельным веществам участки, разбросаны по поверхности клетки или собраны в небольшие зоны. Так, на поверхности прокариотических клеток и клеток животных имеется ограниченное число мест с которыми могут связываться вирусные частицы. Мембранные белки (переносчики и каналы) узнают, взаимодействуют и переносят лишь определенные вещества. Клеточные рецепторы участвуют в передаче сигналов с поверхности клетки внутрь ее. Разнообразие и специфичность наборов рецепторов на поверхности клеток ведет к созданию очень сложной системы маркеров, позволяющих отличать свои клетки от чужих. Сходные клетки взаимодействуют друг с другом, поверхности их могут слипаться (конъюгация у простейших, образование тканей у многоклеточных). Клетки не воспринимающие маркеры, а также отличающиеся набором детерминантных маркеров уничтожаются или отторгаются. При образовании комплекса рецептор-лиганд активируются трансмембранные белки: белок преобразователь, белок усилитель. В результате рецептор изменяет свою конформацию и взаимодействует с находящимся в клетке предшественником вторичного посредника — мессенджером. Мессенджерами могут быть ионизированный кальций, фосфолипаза С, аденилатциклаза, гуанилатциклаза. Под влиянием мессенджера происходит активация ферментов, участвующих в синтезе циклических монофосфатов — АМФ или ГМФ. Последние изменяют активность двух типов ферментов протеинкиназ в цитоплазме клетки, ведущих к фосфорилированию многочисленных внутриклеточных белков.
Наиболее распространено образование цАМФ, под действием которого усиливается секреция ряда гормонов — тироксина, кортизона, прогестерона, увеличивается распад гликогена в печени и мышцах, частота и сила сердечных сокращений, остеодеструкция, обратное всасывание воды в канальцах нефрона.
Активность аденилатциклазной системы очень велика — синтез цАМФ приводит к десяти тысячному усилению сигнала.
Под действием цГМФ увеличивается секреция инсулина поджелудочной железой, гистамина тучными клетками, серотонина тромбоцитами, сокращается гладкомышечная ткань.
Во многих случаях при образовании комплекса рецептор-лиганд происходит изменение мембранного потенциала, что в свою очередь приводит к изменению проницаемости плазмалеммы и метаболических процессов в клетке.
3.Межклеточные контакты
плазматическая мембрана липопротеиновый рецепторный
У многоклеточных животных организмов плазмолемма принимает участие в образовании межклеточных соединений, обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур.
§Простой контакт. Простой контакт встречается среди большинства прилежащих друг к другу клеток различного происхождения. Представляет собой сближение плазмолемм соседних клеток на расстояние 15-20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток.
§Плотный (замыкающий) контакт. При таком соединении внешние слои двух плазмолемм максимально сближены. Сближение настолько плотное, что происходит как бы слияние участков плазмолемм двух соседних клеток. Слияние мембран происходит не по всей площади плотного контакта, а представляет собой ряд точечных сближений мембран. Роль плотного контакта заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды.
§Пятно сцепления, или десмосома. Десмосома представляет собой небольшую площадку диаметром до 0,5 мкм. В зоне десмосомы со стороны цитоплазмы находится область тонких фибрилл. Функциональная роль десмосом в основном заключается в механической связи между клетками.
§Щелевой контакт, или нексус. При таком типе контакта плазмолеммы соседних клеток на протяжении 0,5-3 мкм разделены промежутком в 2-3 нм. В структуре плазмолемм располагаются специальные белковые комплексы (коннексоны). Одному коннексону на плазматической мембране клетки точно противостоит коннексон на плазматической мембране соседней клетки. В результате образуется канал из одной клетки в другую. Коннексоны могут сокращаться, изменяя диаметр внутреннего канала, и тем самым участвовать в регуляции транспорта молекул между клетками. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевого контакта заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус.
§Синаптический контакт,или синапс. Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. Этот тип соединений характерен для нервной ткани и встречается как между двумя нейронами, так и между нейроном и каким-либо иным элементом. Мембраны этих клеток разделены межклеточным пространством — синаптической щелью шириной около 20-30 нм. Мембрана в области синаптического контакта одной клетки называется пресинаптической, другой — постсинаптической. Около пресинаптической мембраны выявляется огромное количество мелких вакуолей (синаптических пузырьков), содержащих медиатор. В момент прохождения нервного импульса синаптические пузырьки выбрасывают медиатор в синаптичекую щель. Медиатор взаимодействует с рецепторными участками постсинаптической мембраны, что в конечном итоге приводит к передаче нервного импульса. Кроме передачи нервного импульса синапсы обеспечивают жесткое соединение поверхностей двух взаимодействующих клеток.
§Плазмодесмы. Этот тип межклеточных связей встречается у растений. Плазмодесмы представляют собой тонкие трубчатые каналы, соединяющие две соседние клетки. Диаметр этих каналов составляет обычно 40-50 нм. Плазмодесмы проходят сквозь клеточную стенку, разделяющую клетки. В молодых клетках число плазмодесм может быть очень велико (до 1000 на клетку). При старении клеток их число падает за счет разрывов при увеличении толщины клеточной стенки. Функциональная роль плазмодесм заключается в обеспечении межклеточной циркуляции растворов, содержащих питательные вещества, ионы и другие соединения. Через плазмодесмы происходит заражение клеток растительными вирусами.
Специализированные структуры плазматической мембраны
Плазмолемма многих клеток животных образует выросты различной структуры (микроворсинки, реснички, жгутики). Наиболее часто на поверхности многих животных клеток встречаются микроворсинки. Эти выросты цитоплазмы, ограниченные плазмолеммой, имеющие форму цилиндра с закругленной вершиной. Микроворсинки характерны для клеток эпителиев, но обнаруживаются и у клеток других тканей. Диаметр микроворсинок составляет около 100 нм. Число и длина их различны у разных типов клеток. Значение микроворсинок заключается в значительном увеличении площади клеточной поверхности. Это особенно важно для клеток, участвующих во всасывании. Так, в кишечном эпителии на 1 мм2 поверхности насчитывается до 2х108 микроворсинок.
Источник: www.BiblioFond.ru
Основу структурной организации клетки составляют биологические мембраны. Плазматическая мембрана (плазмалемма) — это мембрана, окружающая цитоплазму живой клетки. Мембраны состоят из липидов и белков. Липиды (в основном фосфолипиды) образуют двойной слой, в котором гидрофобные «хвосты» молекул обращены внутрь мембраны, а гидрофильные — к её поверхностям. Молекулы белков могут располагаться на внешней и внутренней поверхности мембраны, могут частично погружаться в слой липидов или пронизывать её насквозь. Большая часть погруженных белков мембран — ферменты. Это жидкостно-мозаичная модель строения плазматической мембраны. Молекулы белка и липидов подвижны, что обеспечивает динамичность мембраны. В состав мембран входят также углеводы в виде гликолипидов и гликопротеинов (гликокаликс) , располагающихся на внешней поверхности мембраны. Набор белков и углеводов на поверхности мембраны каждой клетки специфичен и является своеобразным указателем типа клеток. Функции мембраны: Разделительная. Она заключается в образовании барьера между внутренним содержимым клетки и внешней средой. Обеспечение обмена веществ между цитоплазмой и внешней средой. В клетку поступают вода, ионы, неорганические и органические молекулы (транспортная функция) . Во внешнюю среду выводятся продукты, образованные в клетке (секреторная функция) . Транспортная. Транспорт через мембрану может проходить разными путями. Пассивный транспорт осуществляется без затрат энергии, путем простой диффузии, осмоса или облегченной диффузии с помощью белков- переносчиков. Активный транспорт — с помощью белков- переносчиков, и он требует затрат энергии (например, натрий-калиевый насос) . Крупные молекулы биополимеров попадают внутрь клетки в результате эндоцитоза. Его разделяют на фагоцитоз и пиноцитоз. Фагоцитоз — захват и поглощение клеткой крупных частиц. Явление впервые было описано И. И. Мечниковым. Сначала вещества прилипают к плазматической мембране, к специфическим белкам-рецепторам, затем мембрана прогибается, образуя углубление. Образуется пищеварительная вакуоль. В ней перевариваются поступившие в клетку вещества. У человека и животных к фагоцитозу способны лейкоциты. Лейкоциты поглощают бактерии и другие твердые частицы. Пиноцитоз — процесс захвата и поглощения капель жидкости с растворенными в ней веществами. Вещества прилипают к белкам мембраны (рецепторам) , и капля раствора окружается мембраной, формируя вакуоль. Пиноцитоз и фагоцитоз происходят с затратой энергии АТФ. Секреторная. Секреция — выделение клеткой веществ, синтезированных в клетке, во внешнюю среду. Гормоны, полисахариды, белки, жировые капли, заключаются в пузырьки, ограниченные мембраной, и подходят к плазмалемме. Мембраны сливаются, и содержимое пузырька выводится в среду, окружающую клетку. Соединение клеток в ткани (за счет складчатых выростов) . Рецепторная. В мембранах имеется большое число рецепторов — специальных белков, роль которых заключается в передаче сигналов извне внутрь клетки.
Источник: otvet.mail.ru
СТРОЕНИЕ И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН. ЛИПИДЫ БИОМЕМБРАН. МЕХАНИЗМЫ ТРАНСПОРТА ВЕЩЕСТВ ЧЕРЕЗ БИОМЕМБРАНЫ
1. Многообразие мембранных структур и выполняемых ими функций.
2. Строение липидов, входящих в состав клеточных мембран: формулы фосфолипидов, гликолипидов, холестерола. Амфипатические свойства липидов мембран.
3. Белки мембран (интегральные, периферические): особенности структуры, свойства, функции. Взаимодействия белков и липидов в биологических мембранах.
4. Асимметрия мембран (примеры). Способность белков и липидов мембран к латеральной диффузии. Ограниченная возможность поперечной диффузии в мембранах.
5. Транспорт веществ через мембраны: простая и облегчённая диффузия, активный транспорт, экзо- и эндоцитоз, их особенности.
6. Na+, K+-АТФ-аза, механизм действия, роль в поддержании трансмембранного потенциала и возбудимости мембраны.
Ссылки по теме:
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. Глава 9. Биомембраны и биоэнергетика. Основные принципы организации биомембран.
| Раздел 9.1 |
Классификация липидов и их роль в организме. |
|
|
|
|
9.1.1. Из курса биоорганической химии известно, что липиды – это большая группа соединений, которые существенно различаются по своему химическому строению и биологической роли. Общими признаками липидов являются:
Основные классы липидов представлены на рисунке 9.1.
Рисунок 9.1. Основные классы липидов. 9.1.2. Липиды выполняют в организме ряд функций. Главными из них являются следующие:
|
|

| Раздел 9.2 |
Классификация и функции биологических мембран. |
|
|
|
|
9.2.1. Биологические мембраны – сложные надмолекулярные структуры, окружающие все живые клетки и образующие в них замкнутые, специализированные компартменты – органеллы. Основные разновидности биологических мембран представлены на рисунке 9.2. Мембрану, ограничивающую цитоплазму клетки снаружи, называют цитоплазматической или плазматической мембраной (1). Название внутриклеточных мембран обычно происходит от названия ограничиваемых или образуемых ими субклеточных структур. Различают ядерную (2), митохондриальную (3), лизосомальную (4) мембраны, мембраны комплекса Гольджи (5), эндоплазматический ретикулум (6) и другие.
Рисунок 9.2. Схематическое изображение органелл животной клетки (объяснения в тексте). 9.2.2. Некоторые примеры функций биологических мембран. Плазматическая мембрана – ограничивает содержимое клетки от внешней среды; осуществляет контакт с другими клетками, получение, обработку и передачу информации внутрь клетки, поддержание постоянства внутренней среды. Ядерные мембраны (внешняя и внутренняя) – образуют ядерную оболочку, которая отделяет хромосомный материал от цитоплазматических органелл; через поры ядерной оболочки происходит транспорт белков и нуклеиновых кислот в ядро и из ядра. Митохондриальные мембраны – осуществляют преобразование энергии в ходе окислительного фосфорилирования, синтез АТФ. Лизосомальные мембраны – ограничивают гидролитические ферменты от цитоплазмы клетки, препятствуют самоперевариванию (аутолизу) клеток, способствуют поддержанию постоянства рН среды в лизосомах. Мембраны эндоплазматического ретикулума – принимают участие в образовании новых мембран, осуществляют синтез белков, липидов, полисахаридов, окисление гидрофобных метаболитов и ксенобиотиков. |
|
| Раздел 9.3 |
Состав и строение мембран. |
|
|
|
|
9.3.1. Все мембраны по своей организации и составу обнаруживают ряд общих свойств. Они:
9.3.2. Схема строения биологической мембраны, представлена на рисунке 9.3. Основу мембраны составляет липидный бислой – двойной слой молекул липидов, которые обладают свойствомамфифильности (содержат как гидрофильные, так и гидрофобные функциональные группы). В липидном бислое гидрофобные участки молекул взаимодействуют между собой, а гидрофильные участки обращены в окружающую водную среду.
Рисунок 9.3. Схема строения биологической мембраны (по Сингеру и Николсону). Мембранные липиды выполняют роль растворителя мембранных белков, создавая жидкую среду, в которой они могут функционировать. По степени влияния на структуру бислоя и по силе взаимодействия с ним мембранные белки делят на интегральные и периферические. Важнейшие особенности интегральных и периферических белков представлены в таблице 9.1. Таблица 9.1 Характеристика мембранных белков По выполняемым функциям белки в составе мембран делятся на
Количество белков в мембранах могут существенно отличаться. Например, в миелиновой мембране, предназначенной для изоляции нервных волокон, белки составляют только 25% массы мембраны, а в мембранах митохондрий, связанных с процессами окислительного фосфорилирования, на долю белков приходится около 75% массы. В плазматической мембране доля белков и липидов примерно одинаковы. Углеводы в составе мембран не представлены самостоятельными соединениями, а обнаруживаются только в соединении с белками (гликопротеины) или липидами (гликолипиды). Длина углеводных цепей колеблется от двух до восемнадцати остатков моносахаридов. Большая часть углеводов расположена на наружной поверхности плазматической мембраны. Функции углеводов в биомембранах – контроль за межклеточными взаимодействиями, поддержание иммунного статуса, рецепция, обеспечение стабильности белковых молекул в мембране. |
|
| Раздел 9.4 |
Липидный состав мембран |
|
|
|
|
9.4.1. Как уже упоминалось (9.1), компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран, представлены на рисунке 9.4. Выучите эти формулы.
Рисунок 9.4. Наиболее часто встречающиеся природные жирные кислоты. Основные особенности строения жирных кислот, входящих в состав природных жиров:
9.4.2. Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины. Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида представлена на рисунке 9.5. Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой).
Рисунок 9.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 9.6). Соединение сфингозина и жирной кислоты получило название церамид.
Рисунок 9.6. Структурные формулы сфингозина и его производных. В сфингомиелинах водород гидроксильной группы у первого углеродного атома в церамиде замещён на фосфохолин. Пример сфингомиелина, содержащего остаток олеиновой кислоты:
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 9.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани.
Рисунок 9.7. Структурные формулы холестерола и его эфира. 9.4.3. Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 9.4.4. Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 9.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 9.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
Рисунок 9.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 9.9,а).
Рисунок 9.9. Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 9.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул. |
|
| Раздел 9.5 |
Трансмембранный перенос веществ. |
|
|
|
|
9.5.1. Одна из главных функций мембран – участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
Рисунок 9.10. Механизмы транспорта молекул через мембрану Простая диффузия — перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы – Н2О, СО2, О2, мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации. Облегчённая диффузия — перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения – при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины. Активный транспорт – также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+, K+, Ca2+, Mg2+, через митохондриальную – протоны. Для активного транспорта веществ характерна кинетика насыщения. 9.5.2. Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+,K+-аденозинтрифосфатаза (Na+,K+-АТФаза или Na+,K+-насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+,K+-АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.
Рисунок 9.11. Схема Na+, K+-насоса. 9.5.3. Перенос через мембраны частиц и высокомолекулярных соединений Наряду с транспортом органических веществ и ионов, осуществляемым переносчиками, в клетке существует совершенно особый механизм, предназначенный для поглощения клеткой и выведения из неё высокомолекулярных соединений при помощи изменения формы биомембраны. Такой механизм называют везикулярным транспортом.
Рисунок 9.12. Типы везикулярного транспорта: 1 — эндоцитоз; 2 — экзоцитоз. При переносе макромолекул происходит последовательное образование и слияние окружённых мембраной пузырьков (везикул). По направлению транспорта и характеру переносимых веществ различают следующие типы везикулярного транспорта: Эндоцитоз (рисунок 9.12, 1) — перенос веществ в клетку. В зависимости от размера образующихся везикул различают: а) пиноцитоз — поглощение жидкости и растворённых макромолекул (белков, полисахаридов, нуклеиновых кислот) с помощью небольших пузырьков (150 нм в диаметре); б) фагоцитоз — поглощение крупных частиц, таких, как микроорганизмы или обломки клеток. В этом случае образуются крупные пузырьки, называемые фагосомами диаметром более 250 нм. Пиноцитоз характерен для большинства эукариотических клеток, в то время как крупные частицы поглощаются специализированными клетками — лейкоцитами и макрофагами. На первой стадии эндоцитоза вещества или частицы адсорбируются на поверхности мембраны, этот процесс происходит без затраты энергии. На следующей стадии мембрана с адсорбированным веществом углубляется в цитоплазму; образовавшиеся локальные впячивания плазматической мембраны отшнуровываются от поверхности клетки, образуя пузырьки, которые затем мигрируют внутрь клетки. Этот процесс связан системой микрофиламентов и является энергозависимым. Поступившие в клетку пузырьки и фагосомы могут сливаться с лизосомами. Содержащиеся в лизосомах ферменты расщепляют вещества, содержащиеся в пузырьках и фагосомах до низкомолекулярных продуктов (аминокислот, моносахаридов, нуклеотидов), которые транспортируются в цитозоль, где они могут быть использованы клеткой. Экзоцитоз (рисунок 9.12, 2) — перенос частиц и крупных соединений из клетки. Этот процесс, как и эндоцитоз, протекает с поглощением энергии. Основными разновидностями экзоцитоза являются: а) секреция — выведение из клетки водорастворимых соединений, которые используются или воздействуют на другие клетки организма. Может осуществляться как неспециализированными клетками, так и клетками эндокринных желёз, слизистой желудочно-кишечного тракта, приспособленными для секреции производимых ими веществ (гормонов, нейромедиаторов, проферментов) в зависимости от определённых потребностей организма. Секретируемые белки синтезируются на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Затем эти белки транспортируются к аппарату Гольджи, где они модифицируются, концентрируются, сортируются, и затем упаковываются в пузырьки, которые отщепляются в цитозоль и в дальнейшем сливаются с плазматической мембраной, так что содержимое пузырьков оказывается вне клетки. В отличие от макромолекул, секретируемые частицы малых размеров, например, протоны, транспортируются из клетки при помощи механизмов облегчённой диффузии и активного транспорта. б) экскреция — удаление из клетки веществ, которые не могут быть использованы (например, удаление в ходе эритропоэза из ретикулоцитов сетчатой субстанции, представляющей собой агрегированные остатки органелл). Механизм экскреции, по-видимому, состоит в том, что вначале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазматической мембраной. |
|
Источник: dendrit.ru
Плазматическая мембрана
Строение и свойства элементарной мембраны. Каждая клетка, независимо от происхождения, имеет структуру, которая служит ей границей, – плазматическую мембрану. Этот термин имеет много синонимов. Плазматическую мембрану называют также цитоплазматической мембраной, клеточной мембраной или плазмалеммой. Толщина плазматической мембраны 8–10 нм, и под электронным микроскопом она выявляется в виде тонкой линии (рис. 5.1–4). Под световым микроскопом границы клеток обычно представлены более объемной структурой: плазматической мембраной и клеточной оболочкой, расположенной поверх плазматической мембраны.
В цитоплазме эукариотической клетки большинство органоидов также окружены мембраной. Среди них можно отметить цистерны эндоплазматического ретикулума (ЭПР), аппарат Гольджи, лизосомы, пероксисомы, вакуоли. Некоторые органоиды имеют две мембраны, это митохондрии, хлоропласты и другие пластиды. Принцип организации всех мембран клетки одинаков, однако каждый тип мембран имеет свои особенности. Описывая организацию мембран, их химический состав, удобнее рассматривать прежде всего общие черты строения для всех мембран. Такую обобщенную мембрану будем называть «элементарная мембрана».
Если смотреть на мембрану сверху, то она имеет мозаичное строение: в липидные поля включены молекулы белка. На поперечном срезе мембраны выявляется двойной слой очень правильно ориентированных липидных молекул, в который погружены молекулы белка. Большая часть белков погружена в липидный бислой перпендикулярно этому слою. В среднем на одну молекулу белка приходится 50 молекул липидов. В весовом соотношении на липиды приходится чуть более 50 %, на белки несколько меньше. Двойной слой липидов обеспечивает барьерные функции мембраны.
К липидам относится большая группа органических соединений, нерастворимых в воде и хорошо растворимых в органических растворителях и жирах. Наиболее типичными липидами мембран являются фосфолипиды, которые образуются на основе трехатомного спирта – глицерина. Фосфолипиды представляют собой сложные эфиры глицерина с двумя жирными кислотами и фосфорной кислотой. Характерной особенностью молекул фосфолипидов и других липидов является наличие полярно заряженной головки, которая соответствует остатку фосфорной кислоты, и двух длинных неполярных хвостов, образованных остатками жирных кислот. Бо́льшая часть полярных липидов имеют сходное строение с небольшими нюансами. Полярная головка является гидрофильной, что означает «люблю воду». Длинные неполярные хвосты – гидрофобные, что означает «боюсь воды». Спонтанные взаимодействия липидных молекул на границе раздела водной среды при высокой их концентрации приводят к образованию двуслойной мембраны, в которой хвосты липидов обращены внутрь, избегая контакта с водой, а головки ориентированы наружу, стремясь к контакту с молекулами воды. Двойные липидные слои могут замыкаться на себя, образуя отсеки, что иногда наблюдается при восстановлении поврежденной мембраны.
Липиды – подвижные молекулы, активно перемещающиеся в своем слое со скоростью до 2 мкм в секунду. Кроме того, они могут вращаться вокруг своей оси. Подвижность липидов обеспечивается очень важным свойством мембраны – текучестью. Благодаря текучести мембрана подвижна, легко образует выросты и впячивания, что имеет большое значение при поглощении высокомолекулярных соединений с поверхности мембраны и при выведении секретируемых веществ во внешнюю среду.
Молекулы липидов в составе мембраны плотно прилегают друг к другу своими головками, создавая барьер для многих молекул. Через двойной слой липидов свободно проходят только небольшие молекулы воды и газы O2, CO2, N2. Для положительно и отрицательно заряженных ионов натрия, калия, кальция, хлора (Na+, K+, Ca2+, Cl – ) и др. двойной липидный слой служит барьером. Свободно проходят через двойной слой липидов молекулы, растворимые в жирах, например этиловый спирт, жирорастворимые витамины. Такое свойство мембраны, когда одни соединения проходят через нее, а другие – нет, называется полупроницаемостью. Этим свойством обладают все биомембраны.
Липидам присущи свойства диэлектрика, благодаря чему на поверхности мембраны всегда существует разность потенциалов. Наружная поверхность клеточной мембраны всегда несет положительный заряд, а внутренняя – отрицательный. Разность потенциалов имеет химическую природу, и определяется концентрацией положительных и отрицательных ионов вблизи мембраны с одной и другой стороны. Наличие разности потенциалов на поверхности клеток имеет большое значение для их функционирования. В частности, проведение нервного импульса связано с деполяризацией мембраны нейронов. В мембранах органелл цитоплазмы присутствуют ферменты, активность которых изменяется при достижении определенной величины разности потенциалов. Примером таких ферментов являются АТФ-синтетазные комплексы, расположенные в мембранах крист митохондрий и тилакоидных мембранах хлоропластов. Некоторые виды пластиножаберных рыб – скаты – имеют по бокам тела электрические органы, генерирующие достаточно мощные электрические заряды.
В составе мембран, помимо липидов, присутствуют белки, которые могут занимать трансмембранное положение, пронизывая два слоя липидов, или располагаться в одном из слоев. Кроме того, молекулы белка могут прилегать к липидной мембране изнутри клетки, со стороны цитоплазмы, соединяясь с другими компонентами мембраны. Такие периферические белковые структуры обычно служат для контакта с нитями цитоскелета. Наличие взаимосвязи мембраны с цитоскелетом делает ее менее текучей, более стабильной.
Белки в составе мембран выполняют множество функций. Прежде всего, это транспортная функция. Заряженные ионы и небольшие молекулы могут проходить через мембрану с помощью транспортных белков. Например, транспортные белки могут иметь каналы определенного диаметра. Транспорт может осуществляться двумя способами: по градиенту концентрации, из места с высокой концентрацией в локальное место с низкой концентрацией, и наоборот, против градиента концентрации. Например, концентрация ионов К+ внутри эритроцита почти в 30 раз выше, чем в плазме крови. Соответственно, по градиенту концентрации ионы калия могли бы выходить в плазму крови, и постепенно концентрация этих ионов внутри и снаружи клетки стала бы одинаковой. Однако этого в живой клетке не происходит, поскольку транспортные белки осуществляют транспорт К+ против градиента концентрации. Это касается не только ионов калия, но и других ионов, а также небольших органических молекул. На транспорт молекул против градиента концентрации требуются большие энергетические затраты. Более 80 % молекул АТФ расходуется на выполнение транспортных функций в клетке. Благодаря работе транспортных белков, осуществляющих транспорт против градиента концентрации, поддерживается гомеостаз клетки и внутренней среды отдельных органоидов.
Многие белки, интегрированные в мембрану, входят в состав рецепторов. Рецепторы – это структуры мембран, которые могут воспринимать сигнал из внешней или внутренней среды клетки и реагировать на этот сигнал изменением своей пространственной структуры, что приводит к локальным изменениям функции мембраны. Рецепторы присутствуют не только в плазматической мембране, где они воспринимают сигнал из среды, окружающей клетку, но и в мембране органоидов. Например, в мембране ЭПР есть рецепторы, обеспечивающие взаимосвязь рибосомы с мембраной.
Таким образом, строение элементарной мембраны описывается в виде белково-липидной мозаики, где липиды образуют двухслойную структуру. Именно липиды обеспечивают барьерную функцию всех мембран.
Вопросы
1. Что такое элементарная мембрана?
2. Назовите основные компоненты элементарной мембраны и их функции.
3. Что такое рецепторы мембран? В чем их значение?
4. Какие свойства мембраны обеспечивают распространение нервного импульса вдоль нейрона?
5. С чем связана текучесть мембраны?
Свойства плазматической мембраны. Плазматической, или клеточной, мембране свойственны некоторые особенности строения и химического состава. В состав двойного слоя фосфолипидов цитоплазматической мембраны входит еще один компонент – холестерол в животной клетке и фитостерол в клетке растений. Холестерол – это стероид, который тоже обладает свойствами липидов. Его полярная ОН-группа ориентирована на поверхности, а гидрофобная замкнутая структура колец внедряется между гидрофобными участками других липидов. Холестерол способствует более плотной упаковке липидов в мембране, что повышает ее барьерную функцию. Молекулы холестерола могут перемещаться из одного слоя липидов в другой, но обычно они скапливаются в наружном слое липидов, делая его толще.
Мембраны некоторых клеток животного происхождения содержат большое количество сфинголипидов – производных аминоспиртов, в которых остаток жирной кислоты содержит 18 атомов углерода. Многие нервные волокна имеют защитную оболочку из вспомогательных шванновских клеток, которые обладают свойствами изолятора, что предотвращает рассеивание электрического потенциала. Плазматическая мембрана шванновских клеток, многократно накрученная на аксон, состоит преимущественно из сфинголипидов, что значительно улучшает свойства электрического изолятора такой мембраны. Нарушение метаболизма сфинголипидов приводит к развитию тяжелых заболеваний человека, сопровождающихся умственной отсталостью, дефектом развития скелета, почечной и печеночной недостаточностью.
Другой обязательной особенностью плазматической мембраны является наличие сложных углеводных компонентов в составе наружной поверхности клеточных мембран. Эти полисахаридные компоненты, расположенные над поверхностью мембран, могут входить в состав клеточной оболочки и служить рецепторами (специфическими или неспецифическими) для различных молекул. Углеводный компонент мембран представлен в основном гликопротеинами – молекулами белков, ковалентно связанных с цепочками углеводов. Углеводы могут быть соединены не только с белками, но и с липидами, образуя гликолипиды, которые расположены в наружном слое плазматической мембраны. Углеводные компоненты гликолипидов и гликопротеинов представляют собой разветвленные или линейные цепочки, в состав которых входят галактоза, манноза, фруктоза, сиаловая кислота, N-ацетилглюкозамин и другие компоненты. Углеводные цепочки могут иметь в своем составе от 1 до 15 мономеров.
Насколько важен углеводный компонент в составе плазматической мембраны, свидетельствует следующий факт: особенностями расположения углеводов плазматической мембраны эритроцитов определяются различия между хорошо известными поверхностными антигенами групп крови человека в системе АВ0 (I, II, III, IV группы крови).
Таким образом, в состав плазматической мембраны, помимо фосфолипидов, могут входить холестерол и сфинголипиды, которые придают ей некоторые дополнительные свойства. Наружная поверхность плазматической мембраны имеет в своем составе углеводные компоненты. Схема строения плазматической мембраны представлена на рисунке 5.2.
Рис. 5.2. Схема молекулярного строения плазматической мембраны (по Хэм, Кормак, 1982).
Вопросы
1. Какую роль играет в плазматической мембране холестерол?
2. В чем особенности мембран шванновских клеток? С чем это связано?
3. Что вы знаете об углеводных компонентах плазматической мембраны? Расположение? Состав? Значение?
Барьерная и транспортная функции плазматической мембраны. Барьерная функция плазматической мембраны обеспечивается ее липидной составляющей, так же как это происходит в элементарной мембране. Холестерол и другие сложные липиды, входящие в состав плазматической мембраны, улучшают барьерные свойства двойного липидного слоя мембран.
Транспорт заряженных неорганических ионов и небольших органических молекул типа глюкозы и аминокислот осуществляют транспортные белки. Известно несколько категорий транспортных белков плазматической мембраны, осуществляющих транспорт по градиенту концентраций. Некоторые транспортные белки внутри молекул имеют транспортные каналы, диаметр которых соответствует определенным ионам. Показана строгая специфичность всех транспортных белков. Транспорт через каналообразующие транспортные белки представляет собой облегченную диффузию и не требует затрат энергии. Перенос осуществляется из среды с высокой концентрацией в среду с низкой концентрацией. Наиболее просто представить себе, что диаметр канала не изменяется, он всегда открыт, и диффузия происходит непрерывно. На самом деле работа каналообразующих белков регулируется. Размер транспортного канала зависит от концентрации ионов и транспортируемых молекул. Например, отмечается более высокая скорость прохождения для ионов натрия, калия (Na+, K+) и значительно ниже скорость транспорта для ионов хлора (Cl – ). Некоторые каналы открываются в ответ на связывание с сигнальной молекулой, так происходит при передаче нервного импульса от одного нейрона к другому. Кроме того, на диффузию влияют факторы окружающей среды, такие как изменение температуры, воздействие химических соединений. Антибиотики, например, увеличивают проницаемость плазматической мембраны.
Помимо каналообразующих белков, существуют белки-переносчики, которые тоже осуществляют транспорт по градиенту концентрации, но работа таких транспортеров требует затрат АТФ. Белки-переносчики избирательно связываются с ионом, но для открытия транспортного канала необходима активация с помощью молекул АТФ. Например, большинство животных клеток поглощает глюкозу из внеклеточной жидкости, где ее концентрация достаточно высока, с помощью специфических белков-переносчиков для глюкозы. Эти транспортеры имеют один канал, открытие которого требует энергетических затрат. Белки-переносчики с одним каналом называются унипортом.
Другие белки-переносчики в своей структуре имеют два канала, транспорт через которые регулируется с помощью АТФ. Например, глюкоза во многие клетки может входить одновременно с Na+. Такой двойной совместный транспорт называется симпортом. Существует еще один вид транспорта с помощью белков-переносчиков – антипорт. Белки антипорта функционируют таким образом, что одно соединение входит в клетку, а одновременно с этим другое выходит. По такому принципу происходит обмен Cl – и HCO3 – .
Таким образом, транспорт заряженных ионов и небольших молекул через плазматическую мембрану по грандиенту концентрации происходит посредством облегченной диффузии с помощью строго специфических каналообразующих белков. Кроме того, такой транспорт осуществляется с помощью специфических белков-переносчиков, открытие транспортных каналов которых требует затрат энергии. Этот транспорт по градиенту концентрации с помощью белков-переносчиков называется пассивным транспортом (рис. 5.3).
Рис. 5.3. Схема двух вариантов транспорта ионов и небольших молекул через плазматическую мембрану по градиенту концентраций: а) облегченная диффузия; б) пассивный транспорт.
Как указывалось ранее, концентрация К+ внутри клетки в 10–30 раз выше, чем снаружи. Для Na+ картина противоположная, его концентрация выше во внешней среде. Такая разница концентраций ионов внутри клетки и в окружающей среде обеспечивается за счет транспорта этих ионов против градиента концентраций через плазматическую мембрану. Транспорт против градиента концентраций осуществляет своеобразная транспортная система – Na,К-АТФаза. Этот транспортный комплекс часто называют насосом, или помпой. Он обнаружен в плазматической мембране всех животных клеток.
Na,К-АТФаза состоит из двух больших субъединиц, обладающих каталитической активностью. В его состав входит третий компонент – небольшой гликопротеин. За один цикл работы этот транспортный комплекс откачивает из клетки 3 иона (Na+) и переносит в клетку 2 иона (К+) против градиента концентрации. При этом затрачивается одна молекула АТФ. Связь Na+ с ферментом происходит со стороны цитоплазмы с затратой молекулы АТФ на фосфорилирование транспортера. Это приводит к изменениям конформации белкового комплекса, в результате Na+ переносится наружу и освобождается во внеклеточное пространство. На внешней стороне связывается К+, что вызывает дефосфорилирование фермента, изменяется его пространственная структура, что приводит к освобождению К+ со стороны цитоплазмы. Таким образом, полезная работа насоса связана с конформационными изменениями белковой молекулы в результате процессов фосфорилирования и дефосфорилирования.
В результате активного транспорта с помощью аналогичных мембранных насосов, работающих против градиента концентрации, в клетке осуществляется регуляция концентраций двухвалентных катионов (Mg2+) и (Ca2+), о которых известно, что в клетке их концентрация велика, а снаружи во много раз меньше.
Против градиента концентраций с помощью специальных транспортных белков через плазматическую мембрану могут проходить моносахара и аминокислоты. До 30 % всей энергии клетки расходуется на транспорт против градиента концентраций. В нервных клетках работа систем транспорта против градиента концентрации требует до 70 % энергии клетки. Транспорт через плазматическую мембрану против градиента концентрации называется активным транспортом.
Подводя итог, можно сказать, что в процессе транспорта ионов и небольших молекул через плазматическую мембрану можно выделить три типа транспорта: облегченная диффузия с помощью каналообразующих белков без затраты АТФ; пассивный транспорт с помощью белков-переносчиков, работа которых активируется посредством АТФ, и активный транспорт с помощью мембранных насосов против градиента концентрации, требующий энергетических затрат.
Макромолекулы (белки, сложные полисахариды, липопротеиды, нуклеиновые кислоты, а также бактерии и фрагменты погибающих клеток) попадают в клетку в результате эндоцитоза. В процессе эндоцитоза вещества или структуры сорбируются на поверхности клеточной мембраны, затем мембрана изгибается в сторону цитоплазмы, образуя сначала ямку (впячивание), а затем маленькую вакуоль, внутри которой находятся поглощенные вещества. Небольшие вакуоли, образованные в процессе эндоцитоза, называются «эндоцитозными вакуолями», или «эндоцитозными везикулами». Их содержимое не смешивается с гиалоплазмой, оно ограничено мембраной, обладающей свойством полупроницаемости. Образование эндоцитозной вакуоли невозможно без вспомогательных белков – клатринов, находящихся в гиалоплазме. Впячивание плазматической мембраны с сорбированными веществами происходит только тогда, когда на внутренней поверхности мембраны образуется специфическая структура из белка клатрина и вспомогательных белков. При этом плазматическая мембрана со стороны цитоплазмы выглядит на электронно-микроскопических фотографиях утолщенной и покрыта волокнистой структурой. Ямки, образованные таким участком мембраны, называются «окаймленными». Клатриновый слой на поверхности мембраны сохраняется и тогда, когда вакуоль отделяется от мембраны и перемещается внутрь клетки. Наличие клатрина на поверхности эндоцитозной везикулы предотвращает слипание отдельных везикул друг с другом и обеспечивает взаимодействие вакуоли с элементами цитоскелета, которые способствуют ее движению в цитоплазме (рис. 5.4). В более глубоких слоях цитоплазмы клатрин отделяется от эндоцитозных вакуолей и возвращается к внутренней поверхности плазматической мембраны. Обычно клатрин покрывает не более 2 % внутренней поверхности мембраны.
Различают несколько типов эндоцитоза в клетках: пиноцитоз, фагоцитоз и эндоцитоз, опосредованный рецепторами. Пиноцитоз – это поглощение с поверхности клетки растворенных в водной среде веществ. Фагоцитоз, впервые описанный И. И. Мечниковым в начале XX в., – это поглощение крупных структур, явление часто встречающееся у одноклеточных животных – амебы, инфузории. У многоклеточных животных способностью к фагоцитозу обладают определенные типы клеток, например, нейтрофилы и макрофаги, принимающие участие в клеточном иммунитете. Принципиальных отличий в протекании перечисленных процессов нет. В современной литературе для описания процессов пиноцитоза и фагоцитоза используется один термин – эндоцитоз. Информация об особенностях эндоцитоза, опосредованного рецепторами, будет изложена при изучении функции рецепторов плазматической мембраны.
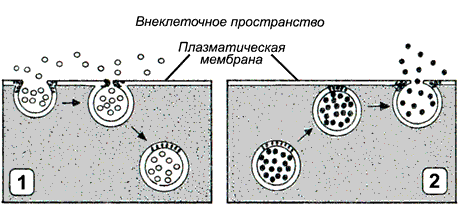
Рис. 5.4. Схема эндоцитоза. 1 – плазматическая мембрана; 2 – вещества на поверхности плазматической мембраны; 3 – белки-клатрины; 4 – эндоцитозная вакуоль.
Попадая во внутренние слои цитоплазмы, эндоцитозные вакуоли могут сливаться с лизосомами, содержащими гидролитические ферменты. Их содержимое переваривается до простых мономеров и неорганических ионов, которые из лизосом поступают в гиалоплазму и используются клеткой в ее метаболизме. Другой путь эндоцитозной вакуоли связан с попаданием в аппарат Гольджи, после чего мембраны эндоцитозных вакуолей в дальнейшем могут использоваться для восстановления плазматической мембраны клетки взамен участка, превратившегося в вакуоль.
Размеры эндосом невелики, составляют десятые или сотые доли микрометра. Но в некоторых клетках эндоцитоз происходит очень интенсивно. Например, клетки эпителия, выстилающие тонкий кишечник, поглощают с поверхности до 1000 эндосом в секунду. При таком активном эндоцитозе плазматическая мембрана будет быстро истощаться. Для ее восстановления клетка использует противоположный процесс – экзоцитоз.
Выведение высокомолекулярных веществ из клетки называется экзоцитозом. В результате экзоцитоза из клетки выводятся секретируемые вещества: молоко из грудных желез, пищеварительные ферменты из клеток поджелудочной железы, гормоны из желез внутренней секреции. Эти вещества длительно накапливаются и выделяются секретирующими клетками при получении определенного сигнала – нервного импульса, воздействия гормона или медиатора. Большинство клеток постоянно выделяют во внешнюю среду химические соединения, воспринимаемые окружающими клетками, или необходимые для нормальной работы ткани и органа. Например, многие клетки секретируют факторы роста, белки-сыворотки крови, антитела, белки, составляющие микроокружение клеток и т. д. В этом случае выделение синтезируемых веществ происходит постоянно, без получения дополнительных сигналов. В любом случае эндоцитоз осуществляется достаточно однотипно. Экзоцитозные вакуоли, содержащие секрет, отграничены от гиалоплазмы мембраной. Они приближаются к мембране, их мембрана сливается с плазматической, в точке контакта образуется выход для выделяемых веществ, и секретируемые вещества попадают в окружающую среду (рис. 5.5). Интенсивный процесс экзоцитоза сопровождается увеличением площади плазматической мембраны.
Таким образом, процессы экзоцитоза и эндоцитоза противоположны друг другу. Активный эндоцитоз, приводящий к уменьшению размеров клетки, должен сопровождаться интенсивным экзоцитозом для восполнения потерь мембраны, и наоборот. В клетке обновление мембраны происходит постоянно.
Рис. 5.5. Схема экзоцитоза. 1 – плазматическая мембрана; 2 – секреторные экзоцитозные вакуоли; 3 – слияние мембран экзоцитозных вакуолей с плазматической мембраной; 4 – выход содержимого экзоцитозных вакуолей на поверхность клетки; 5 – восстановление нормальной структуры плазматической мембраны.
Вопросы
1. Как вы понимаете барьерную функцию плазматической мембраны?
2. Какие виды транспорта осуществляются через плазматическую мембрану?
3. Что такое облегченная диффузия?
4. Как происходит пассивный транспорт?
5. Что такое активный транспорт?
6. Приведите пример мембранного насоса. Как он работает?
7. Что такое эндоцитоз? Как он происходит?
8. Какова роль белка клатрина в эндоцитозе?
9. Что такое экзоцитоз? Объясните сущность процесса.
10. В чем состоит взаимосвязь процессов эндоцитоза и экзоцитоза?
11. Как осуществляется рост и обновление плазматической мембраны?
Рецепторная функция плазматической мембраны. Выполнение клетками своих функций невозможно без способности к межклеточным взаимодействиям – способности посылать сигналы друг другу, уметь распознавать и реагировать на них. Например, специализированные эндокринные клетки секретируют гормоны, которые поступают в кровь и в очень низких концентрациях (обычно 10 – 8 – 10 – 9 М) разносятся по всему организму. Гормональный сигнал воспринимается клетками-мишенями. Способность клетки реагировать на специфические сигналы зависит от наличия у нее на поверхности особых структур – рецепторов, которые узнают и связывают сигнальную молекулу.
Сигнальные молекулы могут быть водорастворимыми и жирорастворимыми. Жирорастворимые – стероидные и тиреоидные гормоны. Водорастворимые сигнальные молекулы связываются с рецепторами на поверхности плазматической мембраны. Это внешнее событие порождает изменение метаболизма клетки. Жирорастворимые сигнальные молекулы способны проникать внутрь клетки и взаимодействовать с рецепторами на клеточных органоидах и на хроматине в ядре. Взаимодействие сигнальной молекулы и ее рецептора всегда сопровождается изменением в работе клетки на разных уровнях регуляции внутриклеточных процессов в зависимости от особенностей рецептора.
Большинство рецепторов плазматической мембраны по своей химической природе являются сложными гликопротеидами, которые пронизывают мембрану насквозь и занимают интегральное положение. В составе рецептора объединены две структуры: наружная, отвечающая за взаимодействие с сигнальной молекулой, и внутренняя часть, являющаяся ферментом, ответственным за синтез вторичного мессенджера – молекулы-посланника, которая синтезируется со стороны цитоплазмы и способствует изменению активности ферментов метаболизма в цитоплазме клетки. Наиболее часто молекулой-посланником является циклический аденозинмонофосфат (цАМФ). Эта молекула образуется из молекулы АТФ при помощи фермента, входящего в состав рецептора и расположенного в мембране со стороны цитоплазмы. Активность этого фермента проявляется только тогда, когда сигнальная молекула из внешней среды вступает во взаимодействие с наружной частью рецептора, подойдя по конфигурации, как ключ к замку. Очередность событий при работе рецептора следующая: сигнальная молекула взаимодействует с наружной частью рецептора, что приводит к изменению его пространственной структуры и вызывает активацию фермента, расположенного со стороны цитоплазмы в составе рецептора. В результате работы фермента из АТФ образуется цАМФ, что способствует изменению активности ферментов цитоплазмы и приводит к изменению метаболизма клетки. Например, глюкагон – гормон, вырабатываемый эндокринной частью поджелудочной железы, действует на рецепторы клеток печени. В этих клетках много гликогена. Под действием глюкагона активируются ферменты, расщепляющие гликоген до глюкозы. Глюкоза поступает в кровь, ее уровень в крови повышается.
Известны разные типы рецепторов, работе которых присуща специфика, при этом путь передачи сигнала внутрь клетки лежит через восприятие сигнала и конформационные изменения части рецептора, расположенной со стороны цитоплазмы. Рецепторы могут воспринимать не только гормоны, но и медиаторы, факторы роста, токсины, вирусы, антитела. Так, например, действие нейромедиатора ацетилхолина в зоне контакта двух нейронов приводит к открытию сразу 2000 ионных каналов для Na+ в рецепторной зоне клетки, воспринимающей сигнал, что вызывает снижение разности потенциалов на мембране воспринимающего нейрона. Рецепторы способны активироваться под действиями факторов различной природы. Например, рецепторы светочувствительных клеток реагируют на свет (фактор физической природы). Результатом такого воздействия является генерация электрического импульса, воспринимаемого нейроном.
После взаимодействия рецептора с сигнальной молекулой его дальнейшая работа прекращается (инактивируется). Инактивированный рецептор выводится из мембраны с образованием эндоцитозной вакуоли, которая сливается с лизосомой, где структура рецептора восстанавливается, и он опять встраивается в мембрану. В ряде случаев восстановление функциональной способности рецептора происходит более сложным путем с включением в работу аппарата Гольджи.
Таким образом, способность клетки воспринимать какой-либо сигнал из внешней среды зависит, прежде всего, от наличия рецептора для этого сигнала. Многообразие сигналов должно соответствовать многообразию рецепторов.
Вопросы
1. Что такое рецептор? Какова его функция в плазматической мембране?
2. Что происходит с рецептором после восприятия сигнала?
3. Что такое вторичный мессенджер? Приведите пример.
4. Приведите пример изменений, которые могут происходить в клетке после воздействия сигнальной молекулы через рецептор.
5. Что такое инактивированный рецептор? Что с ним происходит?
Источник: iknigi.net