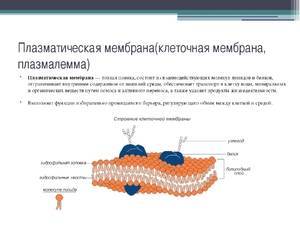
Плазматическая мембрана образована
 Строение клеток живых организмов во многом зависит от того, какие функции они выполняют. Однако существует ряд общих для всех клеток принципов архитектуры. В частности, любая клетка имеет снаружи оболочку, которая называется цитоплазматической или плазматической мембраной. Существует и еще одно название — плазмолемма.
Строение клеток живых организмов во многом зависит от того, какие функции они выполняют. Однако существует ряд общих для всех клеток принципов архитектуры. В частности, любая клетка имеет снаружи оболочку, которая называется цитоплазматической или плазматической мембраной. Существует и еще одно название — плазмолемма.
- Строение
- Функции
- Избирательная проницаемость
- Транспорт веществ
- Пассивный
- Активный
- В мембранной упаковке
- Экзоцитоз
Строение
Плазматическая мембрана состоит из молекул трех основных видов — протеинов, углеводов и липидов. У разных типов клеток соотношение этих компонентов может различаться.
В 1972 году учеными Николсоном и Сингером был предложена жидкостно-мозаичная модель строения цитоплазматической мембраны. Эта модель послужила ответом на вопрос о строении клеточной мембраны и не утратила своей актуальности и по сей день. Суть жидкостно-мозаичной модели заключается в следующем:
- Липиды располагаются в два слоя, составляя основу клеточной стенки;
- Гидрофильные концы липидных молекул расположены внутрь, а гидрофобные — наружу;
- Внутри эта структура имеет слой протеинов, которые пронизывают липиды подобно мозаике;
- Кроме белков здесь имеется небольшое количество углеводов — гексоз;
Эта биологическая система отличается большой подвижностью. Белковые молекулы могут выстраиваться, ориентируясь к одной из сторон липидного слоя, или же свободно перемещаются и меняют свое положение.
Это интересно: сколько хромосом у нормального человека?
Функции
Несмотря на некоторые различия в строении, плазмолеммы всех клеток обладают набором общих функций. Кроме того, они могут обладать характеристиками, сугубо специфичными для данного вида клеток. Рассмотрим кратко общие основные функции всех клеточных мембран:
 Барьерная функция обеспечивает клетке обмен веществ с окружающим пространством. Этот обмен является регулируемым, избирательным и может быть как пассивным, так и активным.
Барьерная функция обеспечивает клетке обмен веществ с окружающим пространством. Этот обмен является регулируемым, избирательным и может быть как пассивным, так и активным.- Транспортная функция заключается в том, что мембрана осуществляет транспорт веществ как в клетку, так и из нее. Таким образом в клетку поставляются питательные вещества, а наружу выводятся продукты метаболизма. Благодаря транспортной функции происходит поддержание в клетке определенного уровня рН, создается ионный градиент и производится секреция различных веществ, необходимых для жизнедеятельности организма.
- Матричная функция обеспечивает белкам определенную локализацию и ориентацию, благодаря чему осуществляется их оптимальное взаимодействие.
- Механическая функция обеспечивает клеткам автономность внутриклеточных образований и одновременно контакт с другими клетками. Немаловажная роль в этом взаимодействии отводится межклеточному веществу.
- Энергетическая функция заключается в переносе белками мембраны энергии в процессе фотосинтеза и клеточном митохондриальном дыхании.
- Рецепторная функция осуществляется за счет некоторых белков-рецепторов. Эти сложные молекулы помогают летке воспринимать те или иные сигналы. В качестве примера можно привести гормоны или нейромедиаторы, которые воздействуют на определенные белки-рецепторы клеток-мишеней.
- Ферментативная функция обеспечивается также за счет белков цитоплазматической мембраны. Часть этих белков могут служить ферментами. К примеру, плазмалеммы кишечного эпителия содержат пищеварительные ферменты.
- Насосная функция плазмолеммы заключается в выработке и проведении потенциалов. Благодаря мембране в клетке поддерживается постоянная концентрация ионов калия и натрия. Это позволяет поддерживать разность потенциалов и проведение нервного импульса.
- Маркерная функция осуществляется благодаря белкам-антигенам, которые позволяют распознавать «свои» и «чужие» клетки. Эти маркеры состоят из белков с присоединенными к ним олигосахаридными цепями. С помощью этих маркеров клетки могут распознавать друг друга в процессе построения тканей, а также при работе иммунной системы организма.
Избирательная проницаемость
Основным свойством плазматической мембраны является избирательная проницаемость. Через нее проходят ионы, аминокислоты, глицерол и жирные кислоты, глюкоза. При этом клеточная мембрана пропускает одни вещества и задерживает другие.
Существует несколько видов механизмов транспорта веществ через клеточную мембрану:
- Диффузия;
- Осмос;
- Экзоцитоз;
- Эндоцитоз;
- Активный транспорт;
Диффузия и осмос не требуют энергетических затрат и осуществляются пассивно, остальные виды транспорта — это активные процессы, протекающие с потреблением энергии.
Такое свойство клеточной оболочки во время пассивного транспорта обусловлено наличием специальных интегральных белков. Такие белки-каналы пронизывают плазмолемму и образуют в ней проходы. Ионы кальция, калия и лора передвигаются по таким каналам относительно градиента концентрации.
Транспорт веществ
К основным свойствам плазматической мембраны относят также ее способность транспортировать молекулы разнообразных веществ.
Описаны следующие механизмы переноса веществ через плазмолемму:
- Пассивный — диффузия и осмос;
- Активный;
- Транспорт в мембранной упаковке;
Рассмотрим эти механизмы более подробно.
Пассивный
К пассивным видам транспорта относятся осмос и диффузия. Диффузией называется движение частиц по градиенту концентрации. В этом случае клеточная оболочка выполняет функции осмотического барьера. Скорость диффузии зависит от величины молекул и их растворимости в липидах. Диффузия, в свою очередь, может быть нейтральной (с переносом незаряженных частиц) или облегченной, когда задействуются специальные транспортные белки.
Осмосом называется диффузия через клеточную стенку молекул воды.
Полярные молекулы с большой массой транспортируются с помощью специальных белков — этот процесс получил название облегченной диффузии. Транспортные белки пронизывают клеточную мембрану насквозь и образуют каналы. Все транспортные белки подразделяются на каналообразующие и транспортеры. Проникновение заряженных частиц облегчается благодаря существованию мембранного потенциала.
Активный
Перенос веществ через клеточную оболочку против электрохимического градиента называется активным транспортом. Такой транспорт всегда происходит с участием специальных белков и требует энергии. Транспортные белки имеют специальные участки, которые связываются с переносимым веществом. Чем больше таких участков, тем быстрее и интенсивнее происходит перенос. В процессе переноса белок транспортер претерпевает обратимые структурные изменения, что и позволяет ему выполнять свои функции.
В мембранной упаковке
Молекулы органически веществ с большой массой переносятся через мембрану с образованием замкнутых пузырьков — везикул, которые образует мембрана.
Отличительной чертой везикулярного транспорта является то, что переносимые макрочастицы не смешиваются с другим молекулами клетки или ее органеллами.
Перенос крупных молекул внутрь клетки получил название эндоцитоза. В свою очередь, эндоцитоз подразделяется на два вида — пиноцитоз и фагоцитоз. При этом часть плазматической мембраны клетки образует вокруг переносимых частиц пузырек, называемый вакуолью. Размеры вакуолей при пиноцитозе и фагоцитозе имеют существенные различия.
В процессе пиноцитоза происходит поглощение клеткой жидкостей. Фагоцитоз обеспечивает поглощение крупных частиц, обломков клеточных органелл и даже микроорганизмов.
Экзоцитоз
Экзоцитозом принято называть выведение из клетки веществ. В таком случае вакуоли перемещаются к плазмолемме. Далее стенка вакуоли и плазмолемма начинают слипаться, а затем сливаться. Вещества, которые содержатся в вакуоли, перемещаются в окружающую среду.
Клетки некоторых простейших организмов имеют строго определенные участки для обеспечения такого процесса.
Как эндоцитоз, так и экзоцитоз протекают в клетке при участии фибриллярных компонентов цитоплазмы, которые имеют тесную непосредственную связь с плазмолеммой.
Источник: obrazovanie.guru
Лекция № 4.
Количество часов: 2
Плазматическая мембрана
1. Строение плазматической мембраны
2. Функции плазматической мембраны. Механизмы транспорта веществ через плазмолемму. Рецепторная функция плазмалеммы
3. Межклеточные контакты.
1. Строение плазматической мембраны
Плазматическая мембрана, или плазмалемма, представляет собой поверхностную периферическую структуру, ограничивающую клетку снаружи и обеспечивающую ее связь с другими клетками и внеклеточной средой.
а имеет толщину около 10 нм. Среди других клеточных мембран плазмалемма является самой толстой. В химическом отношении плазматическая мембрана представляет собой липопротеиновый комплекс. Основными компонентами являются липиды (около 40%), белки (более 60%) и углеводы (около 2-10%).
К липидам относится большая группа органических веществ, обладающих плохой растворимостью в воде (гидрофобность) и хорошей растворимостью в органических растворителях и жирах (липофильность). Характерными представителями липидов, встречающимися в плазматической мембране, являются фосфолипиды, сфингомиелины и холестерин. В растительных клетках холестерин замещается фитостерином. По биологической роли белки плазмалеммы можно разделить на белки-ферменты, рецепторные и структурные белки. Углеводы плазмалеммы входят в состав плазмалеммы в связанном состоянии (гликолипиды и гликопротеины).
В настоящее время общепринятой является жидкостно-мозаичная модель строения биологической мембраны. Согласно этой модели структурную основу мембраны образует двойной слой фосфолипидов, инкрустированный белками. Хвосты молекул обращены в двойном слое друг к другу, а полярные головки остаются снаружи, образуя гидрофильные поверхности.
лекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь на разную глубину (есть периферические белки, часть белков пронизывает мембрану насквозь, часть погружена в слой липидов). Большинство белков не связаны с липидами мембраны, т.е. они как бы плавают в «липидном озере». Поэтому молекулы белков способны перемещаться вдоль мембраны, собираться в группы или, наоборот, рассеиваться на поверхности мембраны. Это говорит о том, что плазматическая мембрана не является статичным, застывшим образованием.
Снаружи от плазмолеммы располагается надмембранный слой — гликокаликс. Толщина этого слоя составляет около 3-4 нм. Гликокаликс обнаружен практически у всех животных клеток. Он представляет собой связанный с плазмолеммой гликопротеиновый комплекс. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами плазматической мембраны. В гликокаликсе могут располагаться белки-ферменты, участвующие во внеклеточном расщеплении различных веществ. Продукты ферментативной активности (аминокислоты, нуклеотиды, жирные кислоты и др.) транспортируются через плазматическую мембрану и усваиваются клетками.
Плазматическая мембрана постоянно обновляется. Это происходит путем отшнуровывания мелких пузырьков с ее поверхности внутрь клетки и встраивания в мембрану вакуолей, поступивших изнутри клетки. Таким образом, в клетке постоянно происходит поток мембранных элементов: от плазматической мембраны внутрь цитоплазмы (эндоцитоз) и поток мембранных структур из цитоплазмы к поверхности клетки (экзоцитоз). В круговороте мембран ведущая роль отводится системе мембранных вакуолей комплекса Гольджи.
4. Функции плазматической мембраны. Механизмы транспорта веществ через плазмолемму. Рецепторная функция плазмалеммы
Плазматическая мембрана выполняет ряд важнейших функций:
1) Барьерная. Барьерная функция плазматической мембраны заключается в ограничении свободной диффузии веществ из клетки в клетку, предотвращении утечки водорастворимого содержимого клетки. Но поскольку клетка должна получать необходимые питательные вещества, выделять конечные продукты метаболизма, регулировать внутриклеточные концентрации ионов, то в ней образовались специальные механизмы переноса веществ через клеточную мембрану.
2) Транспортная. К транспортной функции относится обеспечение поступления и выведения различных веществ в клетку и из клетки. Важное свойство мембраны — избирательная проницаемость, или полупроницаемость. Она легко пропускает воду и водорастворимые газы и отталкивает полярные молекулы, такие как глюкоза или аминокислоты.
Существует несколько механизмов транспорта веществ через мембрану:
пассивный транспорт;
активный транспорт;
транспорт в мембранной упаковке.
Пассивный транспорт. Диффузия — это движение частиц среды, приводящее к переносу вещества из зоны, где его концентрация высока в зону с низкой концентрацией. При диффузионном транспорте мембрана функционирует как осмотический барьер. Скорость диффузии зависит от величины молекул и их относительной растворимости в жирах. Чем меньше размеры молекул и чем более они жирорастворимы (липофильны), тем быстрее произойдет их перемещение через липидный бислой. Диффузия может быть нейтральной (перенос незаряженных молекул) и облегченной (с помощью специальных белков переносчиков). Скорость облегченной диффузии выше, чем нейтральной. Максимальной проникающей способностью обладает вода, так как ее молекулы малы и незаряжены. Диффузия воды через клеточную мембрану называется осмосом. Предполагается, что в клеточной мембране для проникновения воды и некоторых ионов существуют специальные "поры". Число их невелико, а диаметр составляет около 0,3-0,8 нм. Наиболее быстро диффундируют через мембрану легко растворимые в липидном бислое молекулы, например О, и незаряженные полярные молекулы небольшого диаметра (СО, мочевина).
Перенос полярных молекул (сахаров, аминокислот), осуществляемый с помощью специальных мембранных транспортных белков называется облегченной диффузией. Такие белки обнаружены во всех типах биологических мембран, и каждый конкретный белок предназначен для переноса молекул определенного класса. Транспортные белки являются трансмембранными, их полипептидная цепь пересекает липидный бислой несколько раз, формируя в нем сквозные проходы. Это обеспечивает перенос специфических веществ через мембрану без непосредственного контакта с ней. Существует два основных класса транспортных белков: белки-переносчики (транспортеры) и каналообразующие белки (белки-каналы). Белки-переносчики переносят молекулы через мембрану, предварительно изменяя их конфигурацию. Каналообразующие белки формируют в мембране заполненные водой поры. Когда поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них. Если молекула транспортируемого вещества не имеет заряда, то направление транспорта определяется градиентом концентрации. Если молекула заряжена, то на ее транспорт, кроме градиента концентрации, влияет и электрический заряд мембраны (мембранный потенциал). Внутренняя сторона плазмалеммы обычно заряжена отрицательно по отношению к наружной. Мембранный потенциал облегчает проникновение в клетку положительно заряженных ионов и препятствует прохождению ионов заряженных отрицательно.
Активный транспорт. Активным транспортом называется перенос веществ против электрохимического градиента. Он всегда осуществляется белками-транспортерами и тесно связан с источником энергии. В белках-переносчиках имеются участки связывания с транспортируемым веществом. Чем больше таких участков связывается с веществом, тем выше скорость транспорта. Селективный перенос одного вещества называется унипортом. Перенос нескольких веществ осуществляют котранспортные системы. Если перенос идет в одном направлении — это симпорт, если в противоположных – антипорт. Так, например, глюкоза из внеклеточной жидкости в клетку переносится унипортно. Перенос же глюкозы и Na4 из полости кишечника или канальцев почек соответственно в клетки кишечника или кровь осуществляется симпортно, а перенос С1~ и НСО’ антипортно. Предполагается, что при переносе возникают обратимые конформационные изменения в транспортере, что и позволяет премещать соединенные с ним вещества.
Примером белка-переносчика, использующего для транспорта веществ энергию выделившуюся при гидролизе АТФ, является Na+-К+ насос, обнаруженный в плазматической мембране всех клеток. Na+-K насос работает по принципу антипорта, перекачивая Na" из клетки и Кт внутрь клетки против их электрохимических градиентов. Градиент Na+ создает осмотическое давление, поддерживает клеточный объем и обеспечивает транспорт сахаров и аминокислот. На работу этого насоса тратится треть всей энергии необходимой для жизнедеятельности клеток. При изучении механизма действия Na+-K+ насоса было установлено, что он является ферментом АТФазой и трансмембранным интегральным белком. В присутствии Na+ и АТФ под действием АТФа-зы от АТФ отделяется концевой фосфат и присоединяется к остатку аспарагиновой кислоты на молекуле АТФазы. Молекула АТФазы фосфорилируется, изменяет свою конфигурацию и Na+ выводится из клетки. Вслед за выведением Na из клетки всегда происходит транспорт К’ в клетку. Для этого от АТФазы в присутствии К отщепляется ранее присоединенный фосфат. Фермент дефосфорилируется, восстанавливает свою конфигурацию и К1 "закачивается" в клетку.
АТФаза образована двумя субъединицами, большой и малой. Большая субъединица состоит из тысячи аминокислотных остатков, пересекающих бислой несколько раз. Она обладает каталитической активностью и способна обратимо фосфорилироваться и дефосфорилироваться. Большая субъединица на цитоплазматической стороне имеет участки для связывания Na+ и АТФ, а на внешней стороне -участки для связывания К+ и уабаина. Малая субъединица является гликопротеином и функция его пока не известна.
Na+-K насос обладает электрогенным эффектом. Он удаляет три положительно заряженных иона Naf из клетки и вносит в нее два иона К В результате через мембрану течет ток, образующий электрический потенциал с отрицательным значением во внутренней части клетки по отношению к ее наружной поверхности. Na"-K+ насос регулирует клеточный объем, контролирует концентрацию веществ внутри клетки, поддерживает осмотическое давление, участвует в создании мембранного потенциала.
Транспорт в мембранной упаковке. Перенос через мембрану макромолекул (белков, нуклеиновых кислот, полисахаридов, липопротеидов) и других частиц осуществляется посредством последовательного образования и слияния окруженных мембраной пузырьков (везикул). Процесс везикулярного транспорта проходит в две стадии. Вначале мембрана пузырька и плазмалемма слипаются, а затем сливаются. Для протекания 2 стадии необходимо чтобы молекулы воды были вытеснены взаимодействующими липидными бислоями, которые сближаются до расстояния 1-5 нм. Считается, что данный процесс активизируют специальные белки слияния (они выделены пока только у вирусов). Везикулярный транспорт имеет важную особенность — поглощенные или секретируемые макромолекулы, находящиеся в пузырьках, обычно не смешиваются с другими макромолекулами или органеллами клетки. Пузырьки могут сливаться со специфическими мембранами, что и обеспечивает обмен макромолекулами между внеклеточным пространством и содержимым клетки. Аналогично происходит перенос макромолекул из одного компартмента клетки в другой.
Транспорт макромолекул и частиц в клетку называется эндоцитозом. При этом транспортируемые вещества обволакиваются частью плазматической мембраны, образуется пузырек (вакуоль), который перемещается внутрь клетки. В зависимости от размера образующихся пузырьков различают два вида эндоцитоза — пиноцитоз и фагоцитоз.
Пиноцитоз обеспечивает поглощение жидкости и растворенных веществ в виде небольших пузырьков (d=150 нм). Фагоцитоз — это поглощение больших частиц, микроорганизов или обломков органелл, клеток. При этом образуются крупные пузырьки, фагосомы или вакуоли (d-250 нм и более). У простейших фагоцитарная функция — форма питания. У млекопитающих фагоцитарная функция осуществляется макрофагами и нейтрофилами, защищающими организм от инфекции путем поглощения вторгшихся микробов. Макрофаги участвуют также в утилизации старых или поврежденных клеток и их обломков (в организме человека макрофаги ежедневно поглощают более 100 старых эритроцитов). Фагоцитоз начинается только тогда, когда поглощаемая частица свяжется с поверхностью фагоцита и активирует специализированные рецепторные клетки. Связывание частиц со специфическими рецепторами мембраны вызывает образование псевдоподии, которые обволакивают частицу и, сливаясь краями, образуют пузырек —фагосому. Образование фагосомы и собственно фагоцитоз происходит лишь в том случае, если в процессе обволакивания частица постоянно контактирует с рецепторами плазмалеммы, как бы "застегивая молнию".
Значительная часть материала, поглощенного клеткой путем эндоцитоза, заканчивает свой путь в лизосомах. Большие частицы включаются в фагосомы, которые затем сливаются с лизосомами и образуют фаголизосомы. Жидкость и макромолекулы, поглощенные при пиноцитозе, первоначально переносятся в эндосомы, которые также сливаются с лизосомами, образуя эндолизосомы. Присутствующие в лизосомах разнообразные гидролитические ферменты быстро разрушают макромолекулы. Продукты гидролиза (аминокислоты, сахара, нуклеотиды) транспортируются из лизосом в цитозоль, где используются клеткой. Большинство мембранных компонентов эндоцитозных пузырьков из фагосом и эндосом возвращаются с помощью экзоцитоза к плазматической мембране и там повторно утилизируются. Основным биологическим значением эндоцитоза является получение строительных блоков за счет внутриклеточного переваривания макромолекул в лизосомах.
Поглощение веществ в эукариотических клетках начинается в специализированных областях плазматической мембраны, так называемых окаймленных ямках. На электронных микрофотографиях ямки выглядят как впячивания плазматической мембраны, цитоплазматическая сторона которых покрыта волокнистым слоем. Слой как бы окаймляет небольшие ямки плазмалеммы. Ямки занимают около 2% общей поверхности клеточной мебраны эукариотов. В течении минуты ямки растут, все глубже впячиваются, втягиваются в клетку и затем, сужаясь у основания, отщепляются, образуя окаймленные пузырьки. Установлено, что из плазматической мембраны фибробластов в течении одной минуты отщепляется примерно четвертая часть мембраны в виде окаймленных пузырьков. Пузырьки быстро теряют свою кайму и приобретают способность сливаться с лизосомой.
Эндоцитоз может быть неспецифическим (конститутивным) и специфическим (рецепторным). При неспецифическом эндоцитозе клетка захватывает и поглощает совершенно чуждые ей вещества, например, частицы сажи, красители. Вначале происходит осаждение частиц на гликокаликсе плазмалеммы. Особенно хорошо осаждаются (адсорбируются) положительно заряженные группы белков, так как гликокаликс несет отрицательный заряд. Затем изменяется морфология клеточной мембраны. Она может либо погружаться, образуя впячивания (инвагинации), либо, наоборот, формировать выросты, которые как бы складываются, отделяя небольшие объемы жидкой среды. Образование инвагинаций более характерно для клеток кишечного эпителия, амеб, а выростов — для фагоцитов и фибробластов. Заблокировать эти процессы можно ингибиторами дыхания. Образовавшиеся пузырьки — первичные эндосомы, могут сливаться между собой, увеличиваясь в размере. В дальнейшем они соединяются с лизосомами, превращаясь в эндолизосому — пищеварительную вакуоль. Интенсивность жидкофазного неспецифического пиноцитоза довольно высока. Макрофаги образуют до 125, а клетки эпителия тонкого кишечника до тысячи пиносом в минуту. Обилие пиносом приводит к тому, что плазмалемма быстро тратится на образование множества мелких вакуолей. Восстановление мембраны идет довольно быстро при рециклизации в процессе экзоцитоза за счет возвращения вакуолей и их встраивания в плазмалемму. У макрофагов вся плазматическая мембрана замещается за 30 минут, а у фибробластов за 2 часа.
Более эффективным способом поглощения из внеклеточной жидкости специфических макромолекул является специфический эндоцитоз (опосредуемый рецепторами). Макромолекулы при этом связываются с комплементарными рецепторами на поверхности клетки, накапливаются в окаймленной ямке, и затем, образуя эндосому, погружаются в цитозоль. Рецепторный эндоцитоз обеспечивает накопление специфических макромолекул у своего рецептора. Молекулы, которые связываются на поверхности плазмалеммы с рецептором, называются лигандами. При помощи рецепторного эндоцитоза во многих животных клетках идет поглощение холестерина из внеклеточной среды.
Плазмолемма принимает участие в выведении веществ из клетки (экзоцитоз). В этом случае вакуоли подходят к плазмолемме. В местах контактов плазмолемма и мембрана вакуоли сливаются и содержимое вакуоли поступает в окружающую среду. У некоторых простейших места на клеточной мембране для экзоцитоза заранее предопределены. Так, в плазматической мембране некоторых ресничных инфузорий есть определенные участки с правильным расположением крупных глобул интегральных белков. У мукоцист и трихоцист инфузорий полностью готовых к секреции, на верхней части плазмалеммы имеется венчик из глобул интегральных белков. Этими участками мембраны мукоцист и трихоцист соприкасаются с поверхностью клетки. Своеобразный экзоцитоз наблюдается в нейтрофилах. Они способны при определенных условиях выбрасывать в окружающую среду свои лизосомы. При этом в одних случаях образуются небольшие выросты плазмалеммы, содержащие лизосомы, которые затем отрываются и переходят в среду. В других случаях наблюдается инвагинация плазмалеммы вглубь клетки и захват ею лизосом, распложенных далеко от поверхности клетки.
Процессы эндоцитоза и экзоцитоза осуществляется при участии связанной с плазмолеммой системы фибриллярных компонентов цитоплазмы.
Рецепторная функция плазмалеммы. Это одна из главных, универсальных для всех клеток, является рецепторная функция плазмалеммы. Она определяет взаимодействие клеток друг с другом и с внешней средой..
Все многообразие информационных межклеточных взаимодействий схематически можно представить как цепь последовательных реакций сигнал-рецептор-вторичный посредник-ответ (концепция сигнал-ответ). Передачу информации от клетки к клетке осуществляют сигнальные молекулы, которые вырабатываются в одних клетках и специфически влияют на другие, чувствительные к сигналу (клетки-мишени). Сигнальная молекула — первичный посредник связывается с находящимися на клетках-мишенях рецепторами, реагирующими только на определенные сигналы. Сигнальные молекулы —лиганды- подходят к своему рецептору как ключ к замку. Лиганда-ми для мембранных рецепторов (рецепторов плазмалеммы) являются гидрофильные молекулы, пептидные гормоны, нейромедиа-торы, цитокины, антитела, а для ядерных рецепторов — жирорастворимые молекулы, стероидные и тиреоидные гормоны, витамин Д В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокалик-са — полисахариды и гликопротеиды. Считается, что чувствительные к отдельным веществам участки, разбросаны по поверхности клетки или собраны в небольшие зоны. Так, на поверхности прокариотических клеток и клеток животных имеется ограниченное число мест с которыми могут связываться вирусные частицы. Мембранные белки (переносчики и каналы) узнают, взаимодействуют и переносят лишь определенные вещества. Клеточные рецепторы участвуют в передаче сигналов с поверхности клетки внутрь ее. Разнообразие и специфичность наборов рецепторов на поверхности клеток ведет к созданию очень сложной системы маркеров, позволяющих отличать свои клетки от чужих. Сходные клетки взаимодействуют друг с другом, поверхности их могут слипаться (конъюгация у простейших, образование тканей у многоклеточных). Клетки не воспринимающие маркеры, а также отличающиеся набором детерминантных маркеров уничтожаются или отторгаются. При образовании комплекса рецептор-лиганд активируются трансмембранные белки: белок преобразователь, белок усилитель. В результате рецептор изменяет свою конформацию и взаимодействует с находящимся в клетке предшественником вторичного посредника — мессенджером. Мессенджерами могут быть ионизированный кальций, фосфолипаза С, аденилатциклаза, гуанилатциклаза. Под влиянием мессенджера происходит активация ферментов, участвующих в синтезе циклических монофосфатов — АМФ или ГМФ. Последние изменяют активность двух типов ферментов протеинкиназ в цитоплазме клетки, ведущих к фосфорилированию многочисленных внутриклеточных белков.
Наиболее распространено образование цАМФ, под действием которого усиливается секреция ряда гормонов — тироксина, кортизона, прогестерона, увеличивается распад гликогена в печени и мышцах, частота и сила сердечных сокращений, остеодеструкция, обратное всасывание воды в канальцах нефрона.
Активность аденилатциклазной системы очень велика — синтез цАМФ приводит к десяти тысячному усилению сигнала.
Под действием цГМФ увеличивается секреция инсулина поджелудочной железой, гистамина тучными клетками, серотонина тромбоцитами, сокращается гладкомышечная ткань.
Во многих случаях при образовании комплекса рецептор-лиганд происходит изменение мембранного потенциала, что в свою очередь приводит к изменению проницаемости плазмалеммы и метаболических процессов в клетке.
На плазматической мембране находятся специфические рецепторы, реагирующие на физические факторы. Так, у фотосинтезирующих бактерий на поверхности клетки располагаются хлорофиллы, реагирующие на свет. У светочувствительных животных в плазматической мембране находится целая система фогорецепторных белков-родопсинов, с помощью которых световой раздражитель трансформируется в химический сигнал, а затем электрический импульс.
3. Межклеточные контакты
У многоклеточных животных организмов плазмолемма принимает участие в образовании межклеточных соединений, обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур.
§ Простой котакт. Простой контакт встречается среди большинства прилежащих друг к другу клеток различного происхождения. Представляет собой сближение плазмолемм соседних клеток на расстояние 15-20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток.
§ Плотный (замыкающий) контакт. При таком соединении внешние слои двух плазмолемм максимально сближены. Сближение настолько плотное, что происходит как бы слияние участков плазмолемм двух соседних клеток. Слияние мембран происходит не по всей площади плотного контакта, а представляет собой ряд точечных сближений мембран. Роль плотного контакта заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды.
§ Пятно сцепления, или десмосома. Десмосома представляет собой небольшую площадку диаметром до 0,5 мкм. В зоне десмосомы со стороны цитоплазмы находится область тонких фибрилл. Функциональная роль десмосом в основном заключается в механической связи между клетками.
§ Щелевой контакт, или нексус. При таком типе контакта плазмолеммы соседних клеток на протяжении 0,5-3 мкм разделены промежутком в 2-3 нм. В структуре плазмолемм располагаются специальные белковые комплексы (коннексоны). Одному коннексону на плазматической мембране клетки точно противостоит коннексон на плазматической мембране соседней клетки. В результате образуется канал из одной клетки в другую. Коннексоны могут сокращаться, изменяя диаметр внутреннего канала, и тем самым участвовать в регуляции транспорта молекул между клетками. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевого контакта заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус.
§ Синаптический контакт,или синапс. Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. Этот тип соединений характерен для нервной ткани и встречается как между двумя нейронами, так и между нейроном и каким-либо иным элементом. Мембраны этих клеток разделены межклеточным пространством – синаптической щелью шириной около 20-30 нм. Мембрана в области синаптического контакта одной клетки называется пресинаптической, другой – постсинаптической. Около пресинаптической мембраны выявляется огромное количество мелких вакуолей (синаптических пузырьков), содержащих медиатор. В момент прохождения нервного импульса синаптические пузырьки выбрасывают медиатор в синаптичекую щель. Медиатор взаимодействует с рецепторными участками постсинаптической мембраны, что в конечном итоге приводит к передаче нервного импульса. Кроме передачи нервного импульса синапсы обеспечивают жесткое соединение поверхностей двух взаимодействующих клеток.
§ Плазмодесмы. Этот тип межклеточных связей встречается у растений. Плазмодесмы представляют собой тонкие трубчатые каналы, соединяющие две соседние клетки. Диаметр этих каналов составляет обычно 40-50 нм. Плазмодесмы проходят сквозь клеточную стенку, разделяющую клетки. В молодых клетках число плазмодесм может быть очень велико (до 1000 на клетку). При старении клеток их число падает за счет разрывов при увеличении толщины клеточной стенки. Функциональная роль плазмодесм заключается в обеспечении межклеточной циркуляции растворов, содержащих питательные вещества, ионы и другие соединения. Через плазмодесмы происходит заражение клеток растительными вирусами.
Специализированные структуры плазматической мембраны
Плазмолемма многих клеток животных образует выросты различной структуры (микроворсинки, реснички, жгутики). Наиболее часто на поверхности многих животных клеток встречаются микроворсинки. Эти выросты цитоплазмы, ограниченные плазмолеммой, имеющие форму цилиндра с закругленной вершиной. Микроворсинки характерны для клеток эпителиев, но обнаруживаются и у клеток других тканей. Диаметр микроворсинок составляет около 100 нм. Число и длина их различны у разных типов клеток. Значение микроворсинок заключается в значительном увеличении площади клеточной поверхности. Это особенно важно для клеток, участвующих во всасывании. Так, в кишечном эпителии на 1 мм2 поверхности насчитывается до 2х108 микроворсинок.
Источник: studizba.com
Плазматическая мембрана, или плазмолемма, среди клеточных мембран занимает особое место. Это поверхностная периферическая структура, ограничивающая клетку снаружи, что определяет ее непосредственную связь с внеклеточной средой, поэтому она является барьером между внутриклеточным содержимым и внешней средой.
Плазматическая мембрана осуществляет функции, связанные с регулируемым избирательным трансмембранным транспортом веществ и играет роль первичного клеточного анализатора. В этом отношении её можно считать клеточным органоидом, входящим в вакуолярную систему клетки.
Окружая клетку со всех сторон, плазматическая мембрана исполняет роль механического барьера. Механическая устойчивость плазматической мембраны определяется такими дополнительными образованиями как гликокаликс и кортикальный слой цитоплазмы (рис. 127).
Гликокаликс – это внешний по отношению к липопротеидной мембране слой, содержащий полисахаридные цепочки мембранных интегральных белков — гликопротеидов. В состав гликопротеидов входят такие углеводы как манноза, глюкоза, N-ацетилглюкозамин, сиаловая кислота и др.
Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что снижает в слое скорость диффузии различных веществ. В гликокаликсе находятся выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
В электронном микроскопе гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3-4 нм, покрывающего всю поверхность клетки. Гликокаликс обнаружен практически у всех животных клеток, но особенно хорошо он выражен в щеточной каемке всасывающего эпителия кишечника.
Кроме гликокаликса механическую устойчивость плазматической мембране обеспечивают кортикальный слой цитоплазмы и внутриклеточне фибриллярные структуры. Кортикальный (от слова — cortex -кора, кожица) слой цитоплазмы, лежащий в тесном контакте с наружной мембраной, имеет ряд особенностей. В нем в толщине 0,1-0,5 мкм отсутствуют рибосомы и мембранные пузырьки и в большом количестве сосредоточены микрофиламенты и микротрубочки. Основным компонентом кортикального слоя является сеть актиновых микрофибрилл. Здесь же располагается ряд вспомогательных белков, необходимых для движения участков цитоплазмы.
У простейших, особенно у инфузорий, плазматическая мембрана принимает участие в образовании пелликулы, жесткого слоя, определяющего форму клетки.
Выполнение барьерной роли плазмолеммой заключается также в ограничении свободной диффузии веществ. Она проницаема для воды, газов, малых неполярных молекул жирорастворимых веществ, но совершенно не проницаема для заряженных молекул (ионы) и для крупных незаряженных (сахара) (рис. 130).
Естественные мембраны ограничивают скорость проникновения низкомолекулярных соединений в клетку.
Трансмембранныый перенос ионов и низкомоекулярных соединений.Плазматическая мембрана, как и другие липопротеидные мембраны клетки, является полупроницаемой. Чем больше размер молекул, тем меньше скорость прохождения их через мембрану. В этом отношении она является осмотическим барьером. Максимальной проникающей способностью обладает вода и растворенные в ней газы, медленнее проникают сквозь мембрану ионы (примерно в 104 раз медленнее). Если клетку поместить в среду с концентрацией солей ниже, чем в клетке (гипотония), то вода снаружи устремляется внутрь клетки, что приводит к увеличению объема клетки и разрыву плазматической мембраны. И наоборот, при помещении клетки в растворы солей с более высокой концентрацией, чем в клетке, происходит выход воды из клетки во внешнюю среду. Клетка при этом сморщивается и уменьшается в объеме.
Такой пассивный транспорт воды из клетки и в клетку все же идет с низкой скоростью. Скорость проникновения воды через мембрану составляет около 10-4 см/с, что в 100 000 раз меньше скорости диффузии молекул воды через водный слой толщиной 7,5 нм. Оказывается, для проникновения воды и ионов в клеточной мембране существуют специальные “поры”. Число пор не велико и суммарная их площадь составляет лишь 0,06% всей клеточной поверхности.
Плазматическая мембрана с разной скоростью способна транспортировать ионы и многие мономеры, такие как сахара, аминокислоты и др. Скорость прохождения катионов (K+, Na+) более высока, в сравнении со скоростью прохождения анионов (Cl—).
В транспорт ионов через плазмалемму принимают участие мембранные транспортные белки — пермеазы. Они могут вести транспорт в одном направлении одного вещества (унипорт) или нескольких веществ одновременно (симпорт), или же вместе с импортом одного вещества выводить из клетки другое (антипорт). Например, глюкоза входит в клетки симпортно с ионом Na+.
Транспорт ионов может происходить по градиенту концентрации — пассивно без дополнительной затраты энергии. Так, в клетку проникает ион Na+ из внешней среды, где его концентрация выше, чем в цитоплазме. При пассивном транспорте мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые растворенные молекулы проходят через мембрану по градиенту концентрации. Часть каналов открыта постоянно, а другая часть закрывается или открывается в ответ на сигнальные молекулы, либо на изменение внутриклеточной концентрации ионов. В других случаях специальные мембранные белки — переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану (облегченная диффузия) (рис. 131).
В организме животных в цитоплазме клеток концентрация ионов резко отличается от плазмы крови, омывающей клетки. Если суммарные концентрации одновалентных катионов как внутри клеток, так и снаружи практически одинаковы (150 мМ), изотоничны,то в цитоплазме концентрация K+ почти в 50 раз выше, а Na+ ниже, чем в плазме крови.
Это связано с тем, что в клетках существуют мембранные белковые переносчики, которые работают против градиента концентрации, затрачивая при этом энергию за счет гидролиза АТФ. Такой тип переноса носит название активного транспорта, и он осуществляется с помощью белковых ионных насосов. В плазматической мембране находится двухсубъединичная молекула (K+ + Na+)-насоса, которая одновременно является и АТФазой. Этот насос при работе откачивает за один цикл 3 иона Na+ и закачивает в клетку 2 иона K+ против градиента концентрации. При этом затрачивается одна молекула АТФ, идущая на фосфорилирование АТФазы, в результате чего Na+ переносится через мембрану из клетки, а K+ получает возможность связаться с белковой молекулой и затем переносится в клетку (рис. 132). С помощью мембранных насосов и с затратой АТФ происходит также регуляция в клетке концентрации двухвалентных катионов Mg2+ и Ca2+
Работа пермеаз и насосов создает в клетке постоянство концентраций осмотических активных веществ или гомеостаз. Примерно 80% всей АТФ клетки тратится на поддержание гомеостаза.
Совместно с активным транспортом ионов через плазматическую мембрану происходит транспорт различных сахаров, нуклеотидов и аминокислот.
Активный транспорт сахаров и аминокислот в бактериальных клетках связан с градиентом ионов водорода.
Участие специальных мембранных белков в пассивном или активном транспорте низкомолекулярных соединений свидетельствует о выcокой специфичности данных процессов, они меняют при этом свою конформацию и функционируют. Таким образом мембраны выступают как анализаторы, как рецепторы.
Везикулярный перенос: эндоцитоз и экзоцитоз. Любые клеточные мембраны не способны к трансмембранному переносу макромолекул, биополимеров, за исключением мембран, имеющих особые белковые комплексные переносчики — порины (мембраны митохондрий, пластид, пероксисом). В клетку макромолекулы попадают заключенными внутри вакуолей или везикул. Везикулярный перенос разделяют на два вида: экзоцитоз — вынос из клетки макромолекулярных продуктов, и эндоцитоз — поглощение клеткой макромолекул (рис. 133).
При эндоцитозе участок плазмалеммы обволакивает внеклеточный материал и заключает его в мембранную вакуоль, возникшую за счет впячивания плазматической мембраны. В такую первичную вакуоль, или эндосому, могут попадать биополимеры, макромолекулярные комплексы, части клеток или даже целые клетки, в которых они распадаются до мономеров и затем путем трансмембранного переноса попадают в гиалоплазму. Биологическое значение эндоцитоза заключается в получении питательных веществ за счет внутриклеточного переваривания, которое осуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой, содержащей набор гидролитических ферментов (см. ниже).
Эндоцитоз формально разделяют на пиноцитоз и фагоцитоз (рис. 134). Фагоцитоз — захват и поглощение клеткой крупных частиц — был впервые описан И,И, Мечниковым. Фагоцитоз встречается как среди одноклеточных (например, амебы, некоторые хищные инфузории), так и у специализированных клеток многоклеточных животных. Сейчас известно, что фагоцитоз и пиноцитоз протекают очень сходно и различия заключаются лишь в массе поглощенных веществ.
В настоящее время эндоцитоз подразделяют на неспецифический или конститутивный, постоянный и специфический, опосредованный рецепторами (рецепторный). Неспецифический эндоцитоз (пиноцитоз и фагоцитоз) протекает автоматически и приводит к захвату и поглощению совершенно чуждых или безразличных для клетки веществ, например, частичек сажи или красителей.
Неспецифический эндоцитоз сопровождается первоначальной сорбцией захватывающего материала гликокаликсом плазмолеммы. Жидкофазный пиноцитоз приводит к поглощению вместе с жидкой средой растворимых молекул, которые не связываются с плазмолеммой.
На следующем этапе происходят впячивания плазматической мембраны, инвагинации, появляются на поверхности клетки выросты, складки, которые как бы захлестываются, складываются, отделяя небольшие объемы жидкой среды (рис. 135, 136). Первый тип возникновения пиноцитозного пузырька, пиносомы, характерен для клеток кишечного эпителия, эндотелия, для амеб, второй — для фагоцитов и фибробластов. Эти процессы зависят от поступления энергии.
Вслед перестройкой поверхности следуют слипание и слияние контактирующих мембран, которые приводят к образованию пеноцитозного пузырька (пиносома). Она отрывается от клеточной поверхности и уходит вглубь цитоплазмы.
Неспецифический и рецепторный эндоцитоз, приводящие к отщеплению мембранных пузырьков, происходят в окаймленных ямках, специализированных участках плазматической мембраны. В окаймленных ямках со стороны цитоплазмы плазматическая мембрана покрыта тонким (около 20 нм) волокнистым слоем, который на ультратонких срезах как бы окаймляет, покрывает небольшие впячивания, ямки (рис. 137). Эти ямки есть почти у всех клеток животных, они занимают около 2% клеточной поверхности. Окаймляющий слой состоит в основном из белка клатрина, ассоциированного с рядом дополнительных белков. Три молекулы клатрина вместе с тремя молекулами низкомолекулярного белка образуют структуру трискелиона, напоминающего трехлучевую свастику (рис. 138). Клатриновый трискелионы на внутренней поверхности ямок плазматической мембраны образуют рыхлую сеть, состоящую из пяти- и шестиугольников, в целом напоминающую корзинку. Клатриновый слой одевает весь периметр отделяющихся первичных эндоцитозных вакуолей, окаймленных пузырьков.
Клатрин относится к одному из видов т.н. “одевающих” белков (COP — coated proteins). Эти белки связываются с интегральными белками-рецепторами со стороны цитоплазмы и образуют одевающий слой по периметру возникающей пиносомы, первичного эндосомного пузырька — “окаймленного” пузырька. в отделении первичной эндосомы участвуют также белки — динамины, которые полимеризуются вокруг шейки отделяющегося пузырька (рис. 139).
После того как окаймленный пузырек отделится от плазмолеммы и начнет переноситься вглубь цитоплазмы клатриновый слой распадается. После потери клатринового слоя эндосомы начинают сливаться друг с другом.
Интенсивность жидкофазного неспецифического пиноцитоза может быть очень высокой. Так клетка эпителия тонкого кишечника образует до 1000 пиносом в секунду, а макрофаги образуют около 125 пиносом в минуту. Размер пиносом невелик, их нижний предел составляет 60-130 нм, но обилие их приводит к тому, что при эндоцитозе плазмолемма быстро замещается, как бы расходуется на образование множества мелких вакуолей. Так у макрофагов вся плазматическая мембрана заменяется за 30 минут, у фибробластов — за два часа.
Дальнейшая судьба эндосом может быть различной, часть из них может возвращаться к поверхности клетки и сливаться с ней, но большая часть вступает в процесс внутриклеточного пищеварения.
В ходе фагоцитоза и пиноцитоза клетки теряют большую площадь плазмолеммы (см. макрофаги), которая довольно быстро восстанавливается при рециклизации мембран, за счет возвращения вакуолей и их встраивания в плазмолемму. Это происходит вследствие того, что от эндосом или вакуолей, так же как и от лизосом могут отделяться небольшие пузырьки, которые вновь сливаются с плазмолеммой.
Специфический или опосредуемый рецепторами эндоцитоз отличается от неспецифического тем, что поглощаются молекулы, для которых на плазматической мембране имеются специфические рецепторы, ассоциирующиеся только с данным типом молекул. Такие молекулы, связывающиеся с белками-рецепторами на поверхности клеток, называют лигандами.
Примером избирательного эндоцитоза является транспорт в клетку холестерина. Этот липид синтезируется в печени и в комплексе с другими фосфолипидами и белковой молекулой образует т.н. липопротеид низкой плотности (ЛНП), который секретируется клетками печени и кровеносной системой разносится по всему телу (рис. 140). Специальные рецепторы плазматической мембраны, диффузно расположенные на поверхности различных клеток, узнают белковый компонент ЛНП, и образуют специфический комплекс рецептор-лиганд. Затем комплекс перемещается в зону окаймленных ямок, окружается мембраной и погружается вглубь цитоплазмы. В ней поглощенные частицы ЛНП подвергается распаду в составе вторичной лизосомы.
Эндосомы характеризуются более низким значением рН (рН 4-5), более кислой средой, чем другие клеточные вакуоли. Это связано с наличием в их мембранах белков протонного насоса, закачивающих ионы водорода с одновременной затратой АТФ (Н+-зависимая АТФаза). Кислая среда внутри эндосом играет решающую роль в диссоциации рецепторов и лигандов. Кроме того, кислая среда является оптимальной для активации гидролитических ферментов в составе лизосом, которые активируются при слиянии лизосом с эндосомами и приводят к образованию эндолизосомы, в которой и происходит расщепление поглощенных биополимерв.
В некоторых случаях не всегда судьба диссоциированных лигандов связана с лизосомным гидролизом. Так в некоторых клетках после связывания рецепторов плазмолеммы с определенными белками, покрытые клатрином вакуоли погружаются в цитоплазму и переносятся к другой области клетки, где сливаются снова с плазматической мембраной, а связанные белки диссоциируют от рецепторов. Так осуществляется перенос, трансцитозис, некоторых белков через стенку эндотелиальной клетки из плазмы крови во межклеточную среду (рис. 141). Другой пример трансцитоза — перенос антител. Так у млекопитающих антитела матери, могут передаваться детенышу через молоко. В этом случае комплекс рецептор-антитело остается в эндосоме без изменений.
Фагоцитоз Фагоцитоз является вариантом эндоцитоза и связан с поглощением клеткой крупных агрегатов макромолекул вплоть до живых или мертвых клеток. Так же как и пиноцитоз, фагоцитоз может быть неспецифическим и специфическим, опосредуемым рецепторами на поверхности плазматической мембраны фагоцитирующих клеток. При фагоцитозе происходит образование больших эндоцитозных вакуолей — фагосом, которые сливаясь с лизосомами образуют фаголизосомы.
На поверхности клеток, способных к фагоцитозу (у млекопитающих это нейтрофилы и макрофаги) существует набор рецепторов, взаимодействующих с белками-лигандами. Так при бактериальных инфекциях антитела к белкам бактерий связываются с поверхностью бактериальных клеток, образуют слой, который узнается специфическими рецепторами на поверхности макрофагов и нейтрофилов, и в местах их связывания начинается поглощение бактерии путем обволакивания ее плазматической мембраной клетки (рис. 142).
Экзоцитоз. Плазматическая мембрана принимает участие в выведении веществ из клетки с помощью экзоцитоза — процесса, обратного эндоцитозу (см. рис. 133).
В случае экзоцитоза, внутриклеточные вакуоли или пузырьки подходят к плазматической мембране. В местах контактов плазматическая и вакуолярная мембраны сливаются, и пузырек опустошается в окружающую среду.
С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Экзоцитоз или секреция в большинстве случаев происходит в ответ на внешний сигнал (нервный импульс, гормоны, медиаторы и др.). В ряде случаев экзоцитоз происходит постоянно (секреция фибронектина и коллагена фибробластами). Сходным образом из цитоплазмы растительных клеток выводятся некоторые полисахариды (гемицеллюлозы), участвующие в образовании клеточных стенок.
Но большинство секретируемых веществ используется другими клетками многоклеточных организмов (секреция молока, пищеварительных соков, гормонов и др.). Часть секретирующих веществ клетки используют для собственных нужд. Например, рост плазматической мембраны осуществляется за счет встраивания участков мембраны в составе экзоцитозных вакуолей, отдельные элементы гликокаликса выделяются клеткой в виде гликопротеидных молекул и т.д.
Выделенные из клеток путем экзоцитоза гидролитические ферменты могут сорбироваться в слое гликокаликса и обеспечивать примембранное внеклеточное расщепление различных биополимеров и органических молекул. Огромное значение примембранное неклеточное пищеварение имеет для животных. Было обнаружено, что в кишечном эпителии млекопитающих в зоне так называемой щеточной каемки всасывающего эпителия, особенно богатой гликокаликсом, обнаруживается огромное количество разнообразных ферментов. Часть этих же ферментов имеет панкреатическое происхождение (амилаза, липазы, различные протеиназы и др.), а часть выделяется собственно клетками эпителия (экзогидролазы, расщепляющие преимущественно олигомеры и димеры с образованием транспортируемых продуктов).
Рецепторная роль плазмалеммы. В качестве рецепторов на поверхности клетки выступают белки мембраны или элементы гликокаликса — гликопротеиды. Чувствительные участки к отдельным веществам могут быть разбросаны по поверхности клетки или собраны в небольшие зоны.
Клетки животных организмов обладают разными наборами рецепторов или же разной чувствительностью одного и того же рецептора.
Многие клеточные рецепторы способны передавать межклеточные сигналы с поверхности внутрь клетки. В настоящее время хорошо изучена система передачи сигнала клеткам с помощью некоторых гормонов, в состав которых входят пептидные цепочки. Они связываются со специфическими рецепторами на поверхности плазматической мембраны клетки. Рецепторы, после связи с гормоном активируют другой белок, лежащий уже в цитоплазматической части плазматической мембраны, — аденилатциклазу. Этот фермент синтезирует молекулу циклического АМФ из АТФ. Циклического АМФ (цАМФ) является вторичным мессенджером — активатором ферментов — киназ, вызывающих модификации других белков-ферментов. Так, при действии на печеночную клетку гормона поджелудочной железы глюкагона, вырабатываемого А-клетками островков Лангерганса стимулируется активация аденилатциклазы. Синтезированный цАМФ активирует протеинкиназу А, которая активирует каскад ферментов, в конечном счете расщепляющих гликоген (запасной полисахарид животных) до глюкозы. Действие инсулина заключается в обратном — он стимулирует вхождение глюкозы в печеночные клетки и отложение ее в виде гликогена.
Эффективность этой аденилатциклазной системы очень высока. Взаимодействие одной или нескольких молекул гормона стимулирует синтез множества молекул цАМФ, что приводит к усилению сигнала в тысячи раз. В данном случае аденилатциклазная система служит преобразователем внешних сигналов.
Другим примером рецепторной активности являются рецепторы ацетилхолина. Ацетилхолин, освобождаясь из нервного окончания, связывается с рецептором на мышечном волокне, вызывает импульсное поступление Na+ в клетку (деполяризация мембраны), открывает сразу около 2000 ионных каналов в зоне нервно-мышечного окончания.
Разнообразие и специфичность наборов рецепторов на поверхности клеток создаёт сложную систему маркеров, позволяющих отличать свои клетки (той же особи или того же вида) от чужих. Сходные клетки вступают друг с другом во взаимодействия, приводящие к слипанию поверхностей (конъюгация у простейших и бактерий, образование тканевых клеточных комплексов). При этом клетки, отличающиеся набором детерминантных маркеров или не воспринимающие их, либо исключаются из такого взаимодействия, либо у высших животных уничтожаются в результате иммунологических реакций (см. ниже).
В плазматической мембране находятся специфические рецепторы, реагирующие на физические факторы. Так, в плазматической мембране фотосинтетических бактерий и синезеленых водорослей расположены белки-рецепторы (хлорофиллы), взаимодействующими с квантами света. В плазматической мембране светочувствительных клеток животных локализована фоторецепторные белки (родопсин), с помощью которых световой сигнал превращается в химический, а затем в электрический.
Межклеточное узнавание. В многоклеточных организмах клетки остаются в связи друг с другом за счет способности их поверхностей слипаться. Это свойство адгезии (соединения, сцепления) клеток определяется свойствами их поверхности и обеспечивается взаимодействием между гликопротеидами плазматических мембран. При таком межклеточном взаимодействии клеток между плазматическими мембранами всегда остается щель шириной около 20 нм, заполненная гликокаликсом.
Было установлено, что за взаимодействие однородных клеток отвечают трансмембранные гликопротеиды. Непосредственно за соединение, адгезию, клеток отвечают молекулы т.н. CAM-белков (cell adhesion molecules). Некоторые из них связывают клетки друг с другом за счет межмолекулярных взаимодействий, другие образуют специальные межклеточные соединения или контакты.
Когда соседние клетки связываются друг с другом с помощью однородных молекул адгезивных белков взаимодействия называется гомофильным, и когда в адгезии участвуют разного рода CAM на соседних клетках- гетерофильным. Встречается межклеточное связывание через дополнительные линкерные молекулы.
Из CAM-белков выделяют несколько классов. Это кадгерины, молекулы адгезии нервных клеток (иммуноглобулино-подобные N-CAM), селектины, интегрины.
Кадгерины представляют собой интегральные фибриллярные мембранные белки, которые образуют параллельные гомодимеры. Отдельные домены этих белков связаны с ионами Ca2+, что придает им определенную жесткость. Кадгеринов насчитывают более 40 видов. Так Е-кадгерин характерен для клеток преимплантированных эмбрионов и для эпителиальных клеток взрослых организмов. P-кадгерин характерен для клеток трофобласта, плаценты и эпидермиса.
Молекулы адгезии нервных клеток (N-CAM) принадлежат к суперсемейству иммуноглобулинов, они образуют связи между нервными клетками. Некоторые из N-CAM участвуют в соединении синапсов, а также при адгезии клеток иммунной системы.
Селектины также интегральные белки плазматической мембраны участвуют в адгезии эндотелиальных клеток, в связывании кровяных пластинок, лейкоцитов.
Интегрины представляют собой гетеродимеры, с a и b-цепями. Интегрины в первую очередь осуществляют связь клеток с внеклеточными субстратами, но могут участвовать и в адгезии клеток друг с другом.
Узнавание чужеродных белков. На попавшие в организм чужеродные макромолекулы (антигены), развивается иммунная реакция. Суть ее заключается в том, что часть лимфоцитов вырабатывает специальные белки — антитела, которые специфически связываются с антигенами. Так, например, макрофаги своими поверхностными рецепторами узнают комплексы антиген-антитело и поглощают их (например, поглощение бактерий при фагоцитозе).
В организме всех позвоночных также существует система рецепции чужеродных клеток или же своих, но с измененными белками плазматической мембраны, например при вирусных инфекциях или при мутациях, часто связанных с опухолевым перерождением клеток.
На поверхности всех клеток позвоночных располагаются белки, т.н. главного комплекса гистосовместимости (major histocompatibility complex — MHC). Это интегральные белки гликопротеины, гетеродимеры. Каждый индивидуум имеет свой набор таких белков MHC. Это приводит к тому, что каждая клетка данного организма отличается от клеток индивидуума этого же вида. Специальная форма лимфоцитов, Т-лимфоциты, узнают MHC своего организма и малейшие изменения в его структуре (например, связь с вирусом, или результат мутации в отдельных клетках), приводит к тому, что Т-лимфоциты узнают такие изменившиеся клетки и их уничтожают. Но уничтожают не путем фагоцитоза а выделяют из секреторных вакуолей белки-перфорины, которые встраиваются в цитоплазматическую мембрану измененной клетки, образуют в ней трансмембранные каналы, тем самым делают плазматическую мембрану проницаемой, что и приводит к гибели измененной клетки (рис. 143, 144).
Источник: studopedia.ru