Ферменты лизосом образуются в
Лизосо́ма (от греч. λύσις — разложение и σώμα — тело) — окружённая мембраной клеточная органелла, в полости которой поддерживается кислая среда и находится множество растворимых гидролитических ферментов[1]. Лизосома отвечает за внутриклеточное переваривание макромолекул, в том числе при аутофагии; лизосома способна к секреции своего содержимого в процессе лизосомного экзоцитоза; также лизосома участвует в некоторых внутриклеточных сигнальных путях, связанных с метаболизмом и ростом клетки[2].
Лизосома является одним из видов везикул и относится к эндомембранной системе клетки[3]. Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
Лизосомы были открыты в 1955 году бельгийским биохимиком Кристианом де Дювом[4].
зосомы есть во всех клетках млекопитающих, за исключением эритроцитов[5]. У растений к лизосомам по способу образования, а отчасти и по функциям близки вакуоли. Лизосомы есть также у большинства протистов (как с фаготрофным, так и с осмотрофным типом питания) и у грибов. Таким образом, наличие лизосом характерно для клеток всех эукариот. У прокариот лизосомы отсутствуют, так как у них отсутствует фагоцитоз и нет внутриклеточного пищеварения.
С нарушением функций лизосом связан ряд наследственных заболеваний у человека, называемых лизосомными болезнями накопления[6].
История открытия[править | править код]
В 1949—1952 годах биохимик Кристиан де Дюв и его студенты, изучавшие действие инсулина в клетках печени крыс, случайно обнаружили неожиданное различие в активности кислой фосфатазы в зависимости от способа выделения. Кислая фосфатаза использовалась ими в качестве стандарта, основным предметом их изучения был фермент глюкозо-6-фосфатаза, вовлечённый в метаболизм инсулина. В ходе экспериментов выяснилось, что при фракционировании клеточного содержимого на центрифуге кислая фосфатаза была ассоциирована с микросомальной фракцией, но проявляла только десятую часть активности в сравнении с простым клеточным экстрактом, причём после нескольких дней хранения микросомальной фракции в холодильнике активность кислой фосфатазы возрастала.
и обнаружении этого феномена первым объяснением было то, что произошла какая-то техническая ошибка. Однако повторение эксперимента неизменно воспроизводило первоначальную картину. Это позволило предположить существование неких окружённых мембраной клеточных частиц, которые содержат внутри себя фермент. С 1952 по 1955 год было открыто ещё несколько кислых гидролаз, связанных с микросомальной фракцией. В 1955 году, который считается годом открытия лизосом, К. де Дюв предложил название «лизосома» для клеточной органеллы, которая окружена мембраной, внутри которой поддерживается низкий pH и внутри которой находится ряд ферментов, оптимально работающих в кислой среде[7][8]. В том же 1955 году американский цитолог Алекс Новиков (англ.)русск. из Вермонтского университета США, блестяще владевший техникой микроскопии, посетил лабораторию К. де Дюве и смог получить первые электронные фотографии этих органелл, используя препарат частично очищенных лизосом. Позднее в 1961 году Алекс Новиков с помощью гистохимического выявления кислой фосфатазы и электронной микроскопии подтвердил локализацию этого фермента в лизосомах[9][10]. В 1963 году бельгийский биохимик Генри Хэрс, ранее работавший в группе К. де Дюве, обнаружил недостаточность лизосомного фермента α-глюкозидазы у пациентов с болезнью Помпе и высказал предположение о связи других генетических заболеваний с нарушением работы лизосом[11]. В настоящее время более 50 наследственных заболеваний связывают с лизосомной недостаточностью[12].
В 1974 году за свой вклад в раскрытие структурной и функциональной организации клетки К. де Дюв был удостоен Нобелевской премии по медицине[13].
Признаки лизосом[править | править код]
Лизосомы являются гетерогенными по форме, размеру, ультраструктурным и цитохимическим особенностям. В клетках животных размер лизосом составляет обычно менее 1 мкм, хотя в некоторых типах клеток, например, в макрофагах, размер лизосом может превышать несколько микрон. Лизосомы, как правило, имеют сферическую, овальную, иногда тубулярную форму[14]. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных). Лизосомы у животных обычно составляют не более 5 % внутриклеточного объёма[15].
Один из признаков лизосом — наличие в них ряда ферментов (кислых гидролаз), способных расщеплять белки, углеводы, липиды и нуклеиновые кислоты. К числу ферментов лизосом относятся катепсины (тканевые протеазы), кислая рибонуклеаза, фосфолипаза и др. Кроме того, в лизосомах присутствуют ферменты, которые способны отщеплять от органических молекул сульфатные (сульфатазы) или фосфатные (кислая фосфатаза) группы. Всего полость лизосомы содержит около 60 растворимых кислых гидролитических ферментов[2].
Для лизосом характерна кислая реакция внутренней среды, которая обеспечивает оптимум работы лизосомных гидролаз[14]. Обычно pH в лизосомах составляет около 4,5-5, то есть концентрация протонов в них на два порядка выше, чем в цитоплазме. Это обеспечивается активным транспортом протонов, который осуществляет встроенный в мембраны лизосом белок-насос протонная АТФаза[15]. Помимо протонного насоса в мембрану лизосом встроены белки-переносчики для транспорта в цитоплазму продуктов гидролиза макромолекул: аминокислот, сахаров, нуклеотидов, липидов[16].
Высокая активность кислой фосфатазы ранее использовалась как один из маркеров лизосом. В настоящее время более надежным маркером считается присутствие специфических мембранных гликопротеидов — LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома.
Образование лизосом и их типы[править | править код]
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе[17]. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.
Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путём фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
- Ранняя эндосома — в неё поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
- Поздняя эндосома — в неё из ранней эндосомы поступают пузырьки с материалом, поглощённом при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.
- Лизосома — в неё из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.
- Фагосома — в неё попадают более крупные частицы (бактерии и т. п.), поглощённые путём фагоцитоза. Фагосомы обычно сливаются с лизосомой.
- Аутофагосома — окружённый двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.
- Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окружённые одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путём отпочковывания от ядерной оболочки.
- Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал (в частности, липофусцин). В нормальных клетках сливаются с наружной мембраной и путём экзоцитоза покидают клетку. При старении или патологии накапливаются.
Функции лизосом[править | править код]
Функциями лизосом являются:
- переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток)
- аутофагия — уничтожение ненужных клетке структур, к примеру, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки
- автолиз — самопереваривание клетки, приводящее к её гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
- растворение внешних структур (см, например, остеокласты)
Внутриклеточное пищеварение и участие в обмене веществ[править | править код]
У многих протистов и у животных, имеющих внутриклеточное пищеварение, лизосомы участвуют в переваривании пищи, захваченной путём эндоцитоза. При этом лизосомы сливаются с пищеварительными вакуолями. У протистов непереваренные остатки пищи обычно удаляются из клетки при слиянии пищеварительной вакуоли с наружной мембраной.
Многие клетки животных, у которых преобладает полостное пищеварение (например, хордовые) получают питательные вещества из межклеточной жидкости или плазмы крови с помощью пиноцитоза. Эти вещества также вовлекаются в обмен веществ клетки после их переваривания в лизосомах. Хорошо изученный пример такого участия лизосом в обмене веществ — получение клетками холестерина. Холестерин, приносимый кровью в виде ЛПНП, поступает внутрь пиноцитозных везикул после соединения ЛПНП с рецепторами ЛПНП на мембране. Рецепторы возвращаются к мембране из ранней эндосомы, а ЛПНП поступают в лизосомы. После этого ЛПНП перевариваются, а высвободившийся холестерин через мембрану лизосом поступает в цитоплазму.
Косвенно лизосомы участвуют в обмене, обеспечивая десенсибилизацию клеток к воздействию гормонов. При длительном действии гормона на клетку часть рецепторов, связавших гормон, поступают в эндосомы и затем деградируют внутри лизосом. Снижение числа рецепторов понижает чувствительность клетки к гормону.
Аутофагия[править | править код]
Обычно различают два типа аутофагии — микроаутофагия и макроаутофагия. При микроаутофагии, как при образовании мультивезикулярных телец, образуются впячивания мембраны эндосомы или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путём клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды; так, у дрожжей описана микроаутофагия пероксисом и частичная микроаутофагия ядер, при которой клетка сохраняет жизнеспособность.
При макроаутофагии участок цитоплазмы (часто содержащий какие-либо органоиды) окружается мембранным компартментом, похожим на цистерну эндоплазматической сети. В результате этот участок оказывается отгорожен от остальной цитоплазмы двумя мембранами. Затем такая аутофагосома сливается с лизосомой, и её содержимое переваривается. Видимо, макроаутофагия также неизбирательна, хотя часто подчеркивается, что с помощью неё клетка может избавляться от «отслуживших свой срок» органоидов (митохондрий, рибосом и др.).
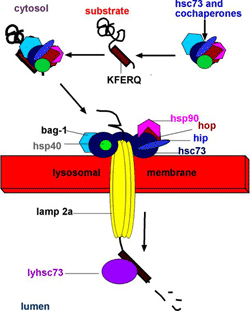
Третий тип аутофагии — шаперон-зависимая. При этом способе происходит направленный транспорт частично денатурировавших белков из цитоплазмы сквозь мембрану лизосомы в её полость.
Автолиз[править | править код]
Ферменты лизосом нередко высвобождаются при разрушении мембраны лизосомы. Обычно при этом они инактивируются в нейтральной среде цитоплазмы. Однако при одновременном разрушении всех лизосом клетки может произойти её саморазрушение — автолиз. Различают патологический и обычный автолиз. Распространенный вариант патологического автолиза — посмертный автолиз тканей.
В норме процессы автолиза сопровождают многие явления, связанные с развитием организма и дифференцировкой клеток. Так, автолиз клеток описывается как механизм разрушения тканей у личинок насекомых при полном превращении, а также при рассасывании хвоста у головастика. Правда, эти описания относятся к периоду, когда различия между апоптозом и некрозом ещё не были установлены, и в каждом случае требуется выяснять, не лежит ли на самом деле в основе деградации органа или ткани апоптоз, не связанный с автолизом.
У растений автолизом сопровождается дифференциация клеток, которые функционируют после смерти (например, трахеид или члеников сосудов). Частичный автолиз происходит и при созревании клеток флоэмы- члеников ситовидных трубок.
Клиническое значение[править | править код]
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером лизосомных болезней накопления может служить болезнь Гоше, болезнь Помпе, Болезнь Тея — Сакса. Всего известно более 50 наследственных заболеваний, связанных с нарушением функции лизосомы[12].
Повреждение лизосом некротизированных клеток, в том числе гранулоцитов, даёт начало воспалительному процессу[18].
См. также[править | править код]
- Вакуоль
- Фагоцитоз
- Эндоцитоз
- Лизосомные болезни накопления
- Восковидные липофусцинозы нейронов
Источник: ru.wikipedia.org
Лизосомы
— клеточные органеллы образованные одним бислоем мембраны. В лизосомах происходит разрушение комплексов лиганд-рецептор
метаболизм холистерола
гидролазы разрушают белки, липиды, углеводы, нуклеиновые кислоты
Внутри лизосом поддерживается постоянный pH = 5, обеспечиваемый ATP-зависимой помпой, которая посредством антипорта Na
+
и H
+
закачивает H
+
внутрь лизосомы. pH поддерживается так же Cl-ионными каналами
Ферменты лизосом: рибонуклеаза, дезоксирибонуклеаза, фосфатаза, гликозидазы, арилсульфатазы (органические эфиры серной кислоты), коллагеназа, катепсины
Структура лизосомного фермента D катепсина с присоединенным сахаром включающим маннозу: PDB = 1LYA.
lgpA lgpB группа интегральных белков 100-120 кДа, сильно гликозилированы. Гликозилирование собственных мембранных белков предотвращает самопереваривание.
Белки лизосом синтезируются в ЭР, проходя транс-сеть АГ, образуют эндосомы которые сливаются
d=0,2-2мкм. деградация клеточных компонентов, ~40 гидролаз (нуклеазы, протеиназы, гликозидазы, липазы, фосфатазы, сульфатазы, фосфолипазы) с оптимумом pH~4,5-5 (в цитоплазме ~7-7,3) — протонные насосы — защита от деградации клетки
образуемые в ШЭР белки гликозилируются в АГ, терминальные маннозные остатки (Man) фосфорилируются по С-6, с образованием концевого остатка —
маннозо-6-фосфат
(Man-6-P). Рецепторы АГ узнают Man-6-P, происходит локальное накопление белков в АГ –
клатрин
– вырезает и транспортируют подходящие мембранные фрагменты в составе транспортных везикул к эндолизосомам. В эндолизосомах происходит понижение pH протонными насосами (H
+
-ATP-аза). Происходит диссоциация белков от рецепторов, отщепление фосфатной группы от Man-6-P. Man-6-P рецепторы используются вторично после рецикла – переносятся в АГ.
транс-Гольджи содержит
манноз 6-фосфатный рецептор связывающий фосфарилированную маннозу лизосомных ферментов,
направляя ферменты к транспортной везикуле.
Первичные лизосомы.
Аутофагия
– захват органеллы =
вторичные лизосомы
– процесс гидролитического расщепления |
остаточные тельца.
Гетероцитоз
– слияние лизосомы с эндосомами эндо- и фагоцитоза.
Мембранные элементы лизосом защищены от действия кислых гидролаз олигосахаридными участками, которые не узнаются ферментами, или мешают гидролазам взаимодействовать с ними.
оксидаза D-аминокислот
– окисляет D-аминокислоты до кетокислот.
Разрушение белков в лизосомах.

Цитоплазматические белки могут разрушаться в протеасомах (см. обзор
Протеасомы
) или в лизосомах. Разрушаемые белки имеют специфический участок узнаваемый шаперонами, которые связываясь с белком транспортируют его к рецепторам на мембране лизосомы. Белок расплетается шаперонами и проникает в канал, ведущий внутрь лизосомы, на другом конце которого протеаза разрезает белок на мелкие фрагменты. Активность такого пути заметно снижается в фибробластах и клетках печени старых крыс. Такое снижение способствует накоплению ненужных белков, нарушая различные клеточные процессы.
Автофагия
При длительном голодании клетка берет энергию и необходимые компаненты для своего выживания разрушая некоторые органеллы. В разрушении органелл участвуют лизосомы.
Органеллы образуемые из лизосом
В некоторых дифференцированных клетках лизосомы могут выполнять специфические функции, образуя дополнительные органеллы. Все
дополнительные функции связаны с секрецией веществ.
| органеллы | клетки | функции |
| меланосомы | меланоциты, ретинальный
пигментный эпителий |
образование, хранение и транспорт меланина |
| тромбоцитные гранулы | тромбоциты, мегакариоциты | освобождение АТФ, АДФ, серотонина и кальция необходимых для свертывания крови |
| ламеллярные тельца | эпителий легких типа II | хранение и секреция сурфактанта необходимого для работы легких |
| лизирующие гранулы | цитотоксические Т
лимфоциты, NK клетки |
разрушение клеток инфицированных вирусом или опухолевы |
| ГКГ класс II | антиген представленные клетки (дендритные клетки, В лимфоциты, макрофаги и др.) |
Изменение и представление антигенов для CD4+ T лимфоцитов для имунной регуляции |
| базофильные гранулы | базофилы, mast клетки | запускают высвобождение гистаминов и других воспалительных стимулов |
| азурофильные гранулы | нейтрофилы, эозинофилы | высвобождают микробицидные и воспалительные агенты |
| остеокластные гранулы | остеокласты | разрушение костей |
| тельца Вейбеля-Палладе | эндотелиальные клетки | созревание и регулируемый выброс фактора Виллебрандта в кровь |
| а-гранулы тромбоцитов | Тромбоциты, мегакариоциты | выброс фибриногена и фактора Виллебрандта для адгезии тромбоцитов и свертывания крови |
Заболевания связанные с лизосомами
Tay-Sachs disease
Pompe’s disease
chediak-Higashi синдром
Hermansky-Pudlak синдром
Griscelli синдром
Некоторые паразиты способны проникать в клетки через лизосомы.
Сокращения.
ЭР — эндоплазматический ретикулум
АГ — аппарат Гольджи
Источник: cellbiol.ru
Форма простейших
В своей форме лизосомы визуализируют в виде плотных шаровидных вакуоли, но они могут показывать значительные отличия размера, в зависимости от материалов, которые были приняты для пищеварения (см.рисунок).
Из таблицы строения и функций лизосом видно, что они обычно сферической формы, с диаметром от 0,05 до 0,5 мкм. На электронных микрофотографиях они имеют форму равномерно-зернистых, электронно-плотных структур (см. схему). Таким образом, особенности строения лизосом в том, что они представляют собой морфологически разнообразные органеллы, проводят определение общей функции деградации внутриклеточного материала.
Это цитоплазматические внутриорганические органеллы, принадлежащие к эндомембранной системе, содержащей ферменты (кислотные гидролазы), которые разрушают многие биологические молекулы. Они встречаются во всех клетках, но чаще — в ответственных за защиту организма: макрофагах, нейтрофильных полинуклеарных или очень специализированных клетках, таких как остеокласты. Первичный видимый в оптической и электронной микроскопии гетерогенный вид лизосом обнаруживается гистоэнзимологической окраской кислой фосфатазы.
Ферменты вида
Лизосомы содержат около 50 различных разлагающих комплексов ферментов, которые могут гидролизовать белки, ДНК, РНК, полисахариды и липиды. Мутации в генах, что кодируют эти ферменты, ответственны за более чем 30 различных генетических заболеваний человека, которые называются лизосомальными заболеваниями хранения, потому что необработанный материал накапливается. Большинство из этих остаточных заболеваний является результатом дефицита отдельных химических лизосомальных ферментов. Например:
- Болезнь под названием Гоше (наиболее распространённое из этих расстройств) является результатом мутации в гене, который кодирует лизосомальный фермент, необходимый для расщепления гликолипидов.
- Клеточное заболевание, которое вызвано дефицитом фермента, катализирующего первый этап мечения лизосомальных ферментов маннозой-6-фосфатом в аппарате Гольджи.
Это всё результат общей неспособности лизосомальных ферментов, которые должны быть включены в органеллах.
Все лизосомальные ферменты являются гидролазами, что активны при рН (около 5), который поддерживается в лизосомах, но не при нейтральном показателе (около 7,2), характерном для остальной цитоплазмы. Требование этих лизосомальных гидролаз для кислотного рН обеспечивает двойную защиту против неконтролируемого пищеварения содержания цитозоля. Даже если лизосомальная мембрана опущена, выпущенные кисловочные гидролазы были бы неактивны на нейтральном рН цитозоля.
Для поддержания кислотного внутреннего рН лизосомы должны активно концентрировать ионы H+ (протоны). Это достигается протонным насосом в лизосомальной мембране, который активно транспортирует протоны в лизосому из цитозоля. Эта перекачка требует затрат энергии в виде гидролиза АТФ, так как она поддерживает примерно в сто раз более высокую концентрацию H+ внутри лизосомы. Органоиды содержат различные кислотные гидролазы, которые активны при кислотном рН, поддерживаемом в органелле, но не при нейтральном рН цитозоля.
Образование органелл
Одной из основных функций лизосом является переваривание материала, взятого извне клетки эндоцитозом. Однако роль органелл в переваривании материала, принимаемого эндоцитозом, связана не только с функцией органелл, но и с их образованием. В частности, лизосомы образуются путём слияния транспортных везикул, почковавшихся из транс-Гольджи сети с эндосомами, которые содержат молекулы, принятые эндоцитозом на плазматической мембране.
Нужно выяснить, где формируются лизосомы и в чём их отличие в биологии. Появление органелл представляет собой пересечение секреторных путей (через них обрабатываются лизосомальные белки) и эндоцитозного пути, через которые молекулы внеклеточного занимают место на поверхности клетки. Материал извне клетки берётся в покрытых клатрином эндоцитарных везикулах, которые распускаются из плазматической мембраны и затем сливаются с ранними эндосомами.
Компоненты затем рециркулируются в плазматическую мембрану и ранние эндосомы постепенно созревают в поздние, которые являются предшественниками лизосом. Одним из важных изменений во время созревания эндосомы является снижение внутреннего рН примерно до 5,5, что играет ключевую роль в доставке лизосомальных кислотных гидролаз из сети транс-гольджи.
Эндоцитоз и образование
Молекулы берутся извне клетки в эндоцитарных везикулах, которые сливаются с ранними эндосомами. Мембранные компоненты перерабатываются по мере созревания ранних эндосом в поздние.
Кислотные гидролазы нацелены на лизосомы остатками манноза-6-фосфата, которые распознаются рецепторами манноза-6-фосфата в сети транс-Гольджи и упаковываются в клатрин-покрытые везикулы. После удаления слоя клатрина эти транспортные везикулы сливаются с поздними эндосомами, а кислотный внутренний рН приводит к диссоциации гидролаз с рецептором манноза-6-фосфата.
Таким образом, гидролазы высвобождаются в просвет эндосомы, в то время как рецепторы остаются в мембране и в итоге рециркулируются в Гольджи. Поздние эндосомы затем созревают в лизосомы, поскольку они приобретают полный набор кислотных гидролаз, которые переваривают молекулы, первоначально принятые эндоцитозом.
Фагоцитоз и аутофагия
Помимо деградирующих молекул, поглощаемых эндоцитозом, лизосомы переваривают материал, полученный двумя другими путями — фагоцитозом и аутофагией. При фагоцитозе специализированные клетки, такие как макрофаги, поглощают и разрушают крупные частицы, включая бактерии, клеточный мусор и старые клетки, которые необходимо удалить из организма.
Такие крупные частицы попадают в фагоцитарные вакуоли (фагосомы), которые затем сливаются с лизосомами, в результате чего происходит переваривание их содержимого. Образующиеся таким образом лизосомы (фаголизосомы) могут быть довольно крупными и неоднородными, так как их размер и форма определяются содержанием перевариваемого материала.
При фагоцитозе крупные частицы, например, бактерии, попадают в фагоцитарные вакуоли или фагосомы. В аутофагии внутренние органеллы, например, митохондрии, заключены в мембранные фрагменты из ER.
Лизосомы также отвечают за аутофагию — постепенный оборот собственных компонентов клетки. Первым шагом аутофагии, по-видимому, является включение органеллы (например, митохондрии) в мембрану, полученную из ER. Образовавшийся пузырёк затем сливается с лизосомой, его содержимое переваривается.
Патофизиологическая роль
У многих примитивных одноклеточных и беспозвоночных микроорганизмов роль лизосом, в первую очередь, питательная. Гетерофагия используется для захвата и переваривания экзогенных продуктов, аутофагия — для контролируемого использования чистых клеточных компонентов в случае нехватки пищи. Эта функция потеряла значение у высших животных, где она действует только для некоторых конкретных питательных веществ, таких как холестерин. С другой стороны, лизосомальное пищеварение подходит для многих специализированных функций.
Таким образом, в зависимости от клеточного типа, гетерофагический механизм служит:
- для защиты от захватчиков;
- очистки альвеол лёгких;
- реабсорбции почек;
- перестановки костей;
- катаболизма белков плазмы;
- регуляции гормональных эффектов;
- синтеза гормонов щитовидной железы и многих других физиологических процессов.
Аутофагия играет очень общую роль в многочисленных аутолитических явлениях, которые происходят в обновлении клеточных составляющих, а также в адаптации, дифференциации и развитии растительных компонентов.
Лизосомы часто могут находиться и участвовать в формировании генеза патологических состояний. Гидролазы, которые они размещают, могут повредить цитоплазму, когда они высвобождаются внутри клетки или внеклеточные структуры, если они сбрасываются снаружи. Кроме того, перегрузка лизосом непереваренными материалами и последующее расширение могут поставить под угрозу нормальную клеточную функцию. Благодаря выяснению этих механизмов, многие заболевания оказались проявлениями клеточной пищеварительной патологии:
- внутриклеточные выделения лизосомального содержимого (клеточные структуры повреждаются и часто клетки погибают) ;
- силикозы, воспаления, интоксикации лизосомотропными веществами.
Функциональные механизмы
Лизосомы играют важную роль во внутриклеточном пищеварении. Этот процесс происходит в вакуолях, ограниченных мембраной, вторичными лизосомами, в пределах которых кислотный рН в диапазоне от 4,5 до 5,0, соответствующий требованиям лизосомиальных гидролаз, активно поддерживается протонным насосом, расположенным в мембране.
Эта кислотность объясняет «лизосомотропизм» очень многих слабо основных соединений: они концентрируются в лизосомах с протонацией и приводят к таким явлениям, как жизненная окраска (например, нейтральный красный, оранжевый акридин), вакуолизация осмотическим вызовом воды, например, хлорохин, и различные фармакологические эффекты.
Исключительно гидролазы и лизосомиальная кислотность сбрасываются во внеклеточную полость, где они выполняют своё пищеварительное действие. Субстраты лизосомального пищеварения поступают извне или изнутри клетки. В первом случае (гетерофагия) захват материалов осуществляется фагоцитозом, пиноцитозом или любой другой формой эндоцитоза, т. е. процессом, включающим инвагинацию плазматической мембраны, которая закрывается вокруг захваченного материала и отделяется в цитоплазме в виде герметичной вакуоли, содержащей этот материал.
Объекты и молекулы, охватываемые таким образом, часто фиксируются специфическими рецепторами, расположенными на плазматической мембране (эндоцитоз интерпозированным рецептором). Это явление позволяет клеткам выбирать определённые питательные вещества в их окружающей среде, например, липопротеины низкой плотности, несущие холестерин. Это даёт им, кроме того, способ ограничивать действие гормонов и других активных агентов.
Источник: nauka.club