Генетическая теория
ТЕОРИЯ ГЕНЕТИЧЕСКОГО ЭНЕРГОИНФОРМАЦИОННОГО ЕДИНСТВА МИРА
Б.А.Астафьев
Доктор философских наук, доктор медицинских наук, Академик РАЕН и АБОП, профессор (г. Москва)
Явилась явь с лучами
Утреннего Солнца.
И осветила жизнь землян
Космичным Светом!
Начну со всем известной сказки о курочке Рябе. Эту сказку, наверное, многие из в ас пытались расшифровать. И я тоже, да всё не удавалось. А идея сказки такова. Золотые яйца не раскрывают смысл жизни. Сколько ни пыталась старуха через золотое яйцо познать истину жизни, но так и не смогла. Сколько ни пытался старик − творческое начало − Ян, олицетворяющий атеистическую науку, решить ту же задачу, − не смог. И лишь маленькое живое существо – мышонок, уронив яйцо на пол, завершила их потуги. Но курочка Ряба пообещала снести теперь простое природное, естественное яйцо, которое будет для старика и старухи чем-то особенным, истинным, дорогим, более ценное, чем золотое.
Всё так и случилось. Ни религия − хранительница прошлого, олицетворённая в сказке в образе старухи (Инь), ни атеистическая наука (старик − Ян), поодиночке оказались неспособными решить задачу, которая заложена в яйце Жизни – Геноме Мира. И лишь мышка помогла завершить эти тщетные потуги, т. е. исследования на экспериментальных животных.
Но мечта людей познать мир жила. А.Эйнштейн полагал, что “природа представляет собой реализацию простейших математических элементов”, и надеялся, что эту задачу кому-то в конце концов удастся решить. Тогда бы удалось ответить на огромное число вопросов, которые находятся за семью печатями. Нобелевский лауреат М.Борн высказал свою мечту в следующей фразе: “Было бы идеалом кратко обобщить все законы в едином Законе, универсальной формуле”. Нобелевский лауреат И.Пригожин высказал ту же идею: “Было бы поистине чудом открыть единые основания всех наук”.
Около 50 лет назад мы поставили себе целью найти этот единый Закон. Работая в разных областях медицины и биологии, одновременно мы изучили историю 5-ти веков развития науки. Мы исследовали около 1000 крупных научных открытий мировой науки. В 1992 г. нами была опубликована эволюционно-генетическая теория паразитизма, которая позволила увидеть Вселенную и Мир в ранее неизвестном ракурсе. Эта теория позволила всесторонне описать все нюансы паразито-хозяинных отношений и понять всю их глубину. В ней впервые прозвучало понятие асимметрии равновесия, т. е. не неравновесных или равновесных систем, о которых писали физии и биологи, а именно асимметрично-равновесные, что, как выяснилось позднее, определяется Базовым Геномом Мира.
Эволюционно-генетическая теория паразитизма натолкнула нас на мысль нас на мысль экстраполировать открытые нами и дуги учёными законы и закономерности развития биологических систем на нашу Вселенную и Мир в целом.
С тех пор система Мироздания заняла основное место в наших исследованиях.
В результате к 2005 году нами была разработана Теория генетического энергоинформационного единства Мира, т. е. единого происхождения Мира, опубликованная в пяти наших монографиях и 30 статьях, ссылки на которые имеются в работах многих других авторов.
А теперь остановлюсь на основных положениях Теории Мироздания − Теории генетического энергоинформационного единства Мира.
Научная теория – это система доказанных идей и представлений о Законах и закономерностях структурно-функциональной организации систем жизни и их эволюции.
Научная Теория генетического энергоинформационного единства Мира принадлежит к типу всеобщих теорий и является универсальной Всеобщей Теорией Творения и эволюции Мира. Почему так? – Потому, что она зиждется на открытых Базовом Геноме Мира и Всеобщих Законах Мира.
Базовый Геном Мира – это единый энергоматериальный алгоритм эволюционного развития информационно и генетически единого Мира Космоса. Он выражается формулой, составленной из элементарных, широко известных науке константных величин, объединённых в единую систему, определяющую алгоритм и направление эволюционного развития Мира и его систем. Геном Мира задаёт Всеобщие Законы Мира.
Объектом изучения Теории генетического энергоинформационного единства Мира является Мир. Он понимается нами как бытие и эволюционирование энергии, объединяющей Мировое Пространство, вселенные, галактики, планеты, сообщества, индивидуальные системы.
Предмет исследования − единый алгоритм творения и его эволюционные проявления.
Для большей ясности сравнения объекта и предмета исследования приведём такой пример. В медицине объектом исследования и лечения является человек, а предметом – для стоматолога – зубы, для кардиолога – сердце и сосуды и т. д.
Говоря об алгоритме развития Мира как о предмете исследования, следует заметить, что вопрос этот только предполагался для исследования. Это можно увидеть в цитируемых выше высказываниях А.Эйнштейна, М.Борна и И.Пригожина.
Методы исследования, используемые в теории: интеграция интуитивного восприятия идей с традиционными методами логического анализа, в том числе методы: статистичесий, общей теории систем, исторического анализа и аналогий, проектного моделирования и др.
Элементами теории генетического энергоинформационного единства Мира являются определения, понятия, формулы, модели. Важнейшими понятиями являются Базовый Геном Мира, Закон Творения, Всеобщие Законы Мира, эволюционная константа, эволюционные спираль и конус, квантованная энергия межфазовых переходов, зоны энергетической активностии др.
Основные понятия Теории генетического
энергоинформационного единства Мира
Базовый Геном Мира Космоса – единый энергетический алгоритм эволюционного развития информационно единого Мира Космоса.
Вита-частица (лат. vita – жизнь) – первичная энергоматериальная частица Мира Космоса, или первичный атом водорода, – носительница Базового Генома Атомно-молекулярного Мира.
Вселенная – часть эволюционирующего Мира. В Мире множество вселенных, развивающихся в соответствии с Базовым Геномом Мира и диктуемыми им Едиными Всеобщими Законами.
Ген (греч. genos – происхождение) – единица наследственной информации. Обеспечивает преемственность в поколениях того или иного признака, свойства организма.
Геном – структурно-функциональнаяформула совокупности генов, наследственный код системы (организма), определяющий своеобразие её развития и жизнедеятельности.
Геномы систем (подсистем) – основа их структурно-функциональной организации, завоёванной в ходе фило- и онтогенеза.
Гравитон – положительно заряженная компонента вита-частицы, несущая творяще-созидающий ген.
Антигравитон − отрицательно заряженная компонента вита-частицы, несущая стимулирующий ген.
Дух – высший энергоинформационный потенциал любой мировой системы.
Жизнь – бытие энергии.
Закон – правило структурно-функциональной организации Мира и его систем, нормированное Геномом Мира и обязательное для жизнеобеспечения и эволюционного развития.
Космос (греч. kosmos – вселенная) – существующий Мир в целом.
Мир – вся эволюционно структурированная и неструктурированная энергия пространственно-временного континуума. Мир включает в себя множество вселенных, одной из которых является наша Вселенная.
Мировой Дух – носитель животворящей Мировой энергии, кодированной Базовым Геномом Мира.
Научная теория – система основных идей, дающая представление о существенных закономерностях и связях Мира и его систем (подсистем).
Система – целое, составленное из ряда взаимодействующих элементов, каждый из которых или простая их сумма не обладают всем комплексом качеств, которыми обладает система.
Эволюционная спираль – траектория эволюционного структурно-функционального развития соответственно Базовому Геному Мира, и входящей в его состав эволюционной константы Се.
Эволюционный конус – стереометрическая фрактальная модель полного цикла жизнедеятельности космической системы любого эволюционного уровня, описываемая эволюционной константой.
Эмерджентность (англ. emergence – выход, появление) – скачкообразное (квантовое) приобретение эволюционирующей системой ряда принципиально новых качеств, которыми не обладала ни одна из составляющих еёподсистем (компонентов).
Энергия – субстанция, обладающая способностью к самодвижению, самоорганизации и саморазвитию.
Элементарные частицы в своём развитии пошли иным путём, чем атомно-молекулярный Мир. Они тоже развились из энергонов, но их развитие началось раньше появления на свет вита-частицы. Они генетическим путём построили вита-частицу. И это их доминирующее положение по отношению к сущностям (системам) Атомно-молекулярного Мира сохранилось и прявляется абсолютно во всех системах. Таким образом, Мир Космоса состоит из Тонкого Мира, т. е. Мира элементарных частиц и их комплексов, и Атомно-молекулярного Мира.
Мир элементарных частиц обогнал Мир атомно-молекулярный в своем эволюционном развитии. Именно элементарные частицы, также возникшие из энергонов Протоса-2, сформировали в конце концов вита-частицу, т. е. первичный атом водорода.
Сознание (Со-знание) – генноподобный отпечаток мысли с генетической записью мысли, хранящейся в памяти самого человека или Творца, Создателя, ангела и других управляющих сущностей Мира. Со-знание – слепок мысли, например, с отпечатка на атомах, молекулах кристаллов и другихмолекулярных или тонкоматериальных формирований.
Мысль – генетически воспроизведённое (рождённое) комплексное знание, данное человеку из различных источников его знаний, имеющее целевую направленность.
Мозг – своего рода процессор всех функций организма, включая мыслительную, гормональную, сердечно-сосудистую, зрительную, обонятельную, осязательную, слуховую, вкусовую, пишеварительную, мочеиспускательную, половую и др.
Воспроизведение из памяти тех или иных мыслей или событий – это подсознательный (но не интуитивный) или осознанный акт.
Интуиция – это воспринятое мозгом человека (или животного) истинное знание о наиболее значимых для него событиях, фактах, развивающее идеи, дающая им новый толчок, т.е. эмерджентное знание, записанное затем на структурах памяти – атомах тех или иных отделов головного мозга, ответственных за ту или иную деятельность человека.
Целевое направление жизнедеятельности – чрезвычайно важно для сохранения целостности организма. Отсутствие цели по жизни ведёт к знойному поведению, разрушающему структуры цельного организма.
Интеллектуальное творчество, раскрывающее Законы, закономерности Космоса, планеты, Солнечной и других космических систем и тел – это генетические онтогенетические накопления в генетике рода (в частности, моего, Наталиного), передающиеся через книги, статьи и другие публикации ныне живущим и будущим поколениям.
Назову некоторые основные постулаты теории генетического
энергоинформационного единства Мира. Все они важны для системы образования.
Назовём 6 основных постулатов теории генетического энергоинформационного единства Мира:
1. Мир – это генетически единая энергоинформационная иерархическая система, эволюционирующая по Единым Всеобщим Законам. На основе этой теории нам удалось дать дефиницию Закона как правила структурно-функциональной организации Мира и его систем, диктуемого Геномом Мира.До наших работ людям были известны лишь 9 Всеобщих Законов Мира. Это: Законы 1) гармонии (золотого сечения), 2) тройственного единства, 3) космической иерархии, 4) фазовой цикличности эволюционного развития, 5) гравитации, или всемирного тяготения, 6) подобия, 7) единства, гармоничного развития и борьбы противоположностей, 8) естественного космического отбора, 9) беспредельности эволюции. Остальные, известные до того людям законы, – это законы специальные, частные, т. е. относящиеся к отдельным специальностям или областям наук: физики, химии, биологии и др. К 2002 г. в результате наших исследований уже были известны 48 Всеобщих Законов Мира [3]. В настоящее время известно уже около 180 Законов Мира. Открытию большей части из них способствовала Периодическая система Всеобщих Законов Мира, открытая акад. Н.В.Масловой [6].
2. Универсальным измерителем эволюционных состояний систем Мира служит эволюционная константа (Се) – постоянная величина, определяющая эволюционное структурно-функциональное развитие и самоорганизацию энергоматерии систем Мира:
Сe = Pq : (p–F) = 0,417897343…
Формула эволюционной константы включает известные науке мировые константы и имеет величину, которую можно рассчитать до любого знака за запятой. Она помогает понять, что эволюционное развитие жизни происходит посредством энергетического квантования пространства-времени. Этот процесс последователен и дискретен (прерывист).
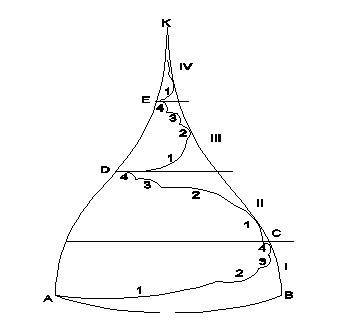
3. Одним из важнейших открытых нами Всеобщих Законов Мира является Закон периодизации эволюции. Суть его в том, что эволюция Мира и всех его систем осуществляется по эволюционной спирали. Она, в свою очередь, образует основу эволюционного конуса – сферической математической модели эволюционного развития любой системы Мира (рис. 1). Издревле (древнеегипетское учение Гермеса Трисмегиста (4 — 1 тысячелетия д.н., древнеегипетское учение – конец 2-го – начало 1-го тысячелетия до н. э.) был известен закон фазовой цикличности эволюционного развития, но оценка длительности фаз была далеко неточной. Поэтому этот закон использовался только как философский, но какие-либо расчёты, отвечающие истине, были невозможны. Нам впервые удалось открыть длительность фаз, которые рассчитываются благодаря знанию эволюционного алгоритма Мира.
Что такое эволюционный конус – это сферическая энергоинформационная модель эволюционного фазового развития систем. Чтобы лучше представить себе его, вспомните купол православной церкви. Каждый виток эволюционного конуса, как и эволюционной спирали, состоит из 4-х фаз, не равных по своей величине. Эволюционный конус, так же как и эволюционная спираль, делится на четыре основные фазы. Их расчёт производится умножением длительности периода эволюционного конуса, например 1000 лет на эволюционную константу Се = 0,4178973, а затем каждый из двух образовавшихся отрезков также умножается на Се. В результате первая фаза составит 0,34 часть высоты ЭК. Вторая и третья фазы – по 0,24 части высоты ЭК. Четвёртая фаза – 0,17 часть. Более тонкое структурирование эволюционной спирали производится последовательными умножениями на Се каждый раз отсекаемого верхнего отрезка ЭК, кратного Се. Например, 1000 лет х 0,4178973 = 417,8973 года. 417,8973 года х 0,4178973 = 174,63815 лет. И т. д. Энергетическая насыщенность по мере приближения к вершине ЭК всё более возрастает, и всё более ускоряется эволюционное развитие систем.
Эволюционный конус включает в себя множество разновеликих конусов, которые организованы принципиально по той же схеме и рассчитываются с использованием эволюционной константы, которая, как мы уже упоминали, является универсальным измерителем развития систем Мира.

Модель эволюционного конуса (ЭК)
АВK– эволюционный конус.
АСDEK–эволюционная спираль развития системы.
I, II, III, IV – фазы полного витка эволюционной спирали (ПВЭС) ACDEK.
I–II фазы – 0,5821027 часть ПВЭС, III–IV фазы – 0,4178973 часть ПВЭС.
K – точка квантового перехода системы на новый этап её развития. 1, 2, 3, 4 – фазы второго порядка (подэтапы) внутри этапов развития системы.
4. Теория вводит в научный оборот понятия о циклах, ритмах, осях, траектории эволюционного развития Мира, Вселенной, галактик, Солнца и планеты Земля. Фрактальная модель (в соответствии с которой рассчитываются фазы 2, 3, 4 и т. д. порядков) позволяет понять взаимодействие подсистем в единой системе.
5. Геном Мира ! ведущее понятие теории. Это ! обязательная и всеобщая норма энергии в некотором объёме пространства-времени. Только при наличии её возникает жизнь как бытие и эволюция одухотворённой энергоматерии. Геном Мира – первая всеобщая биотехнология рождения материальной жизни. Первичный энергоинформационный волновой пакет творения назван нами “вита-частицей”. Её структуру создают две противоположно вращающиеся частицы – гравитон и антигравитон.
Нами рассчитано, что шифром “запуска” алгоритма является перевес творяще-созидающей энергии над стимулирующей на 0,015227 часть их суммарной энергии.
Представьте себе, что женская половая клетка (гравитон) оплодотворяется мужской (антигравитон). По Закону подобия, который действует на любом этапе развития живого. При этом рождается зигота, которая несёт все генетические признаки (задатки) будущего существа. Самая первая вита-частица – это фактически Зигота Мира, давшая начало всему Космическому Миру. Именно с этого момента зародилась энергоматерия, которая до того была лишь нематериализованной энергией.
где ℮0,5H–D℮ – это стимулирующая энергия, равная энергии полумассы атома водорода минус 0,0076135 её часть; ℮0,5H+D℮ – это творяще-созидающая энергия полумассы атома водорода плюс 0,0076135 её часть; Pq – квантовая константа Астафьева[1], равная 1,05458846…; p = 3,141592653… – круговая (сферическая) константа; F = 0,618033989… – константа золотого сечения [1-5].
Геном – совокупность генов – генетический (наследственный) код организма (системы), определяющий своеобразие его развития и жизнедеятельности. Геном Мира содержит три гена: константный творяще-созидающий[℮0,5H+D℮ Pq : (p–F)], константный стимулирующий [℮0,5H–D℮ Pq : (p—F)]-1.Оба они содержат три известные физические константы: число p, константу золотого сечения F и квантовую константу Pq.Объединённые открытой нами формулой они формируют эволюционную константу Се = Pq : (p–F) = 0,417897343… Третий ген – это ген творения Се3∙ ℮n–1, или программный ген, творческий потенциал которого обусловлен качеством накопленной в фило- и онтогенезе (т. е. родовом и индивидуальном развитии) энергии.
Учитывая, что ген творения включает филогенетическую (собственно генетическую) и онтогенетическую (фенотипическую) энергетические составляющие, он может быть представлен формулой Се3∙ ℮f + o , где индекс f означает филогенетический опыт системы, а индекс о — её онтогенетический (фенотипический) опыт. Этот ген можно представить также в виде Се3∙ (℮f + ℮o). Ген Се3∙ ℮f. Это ген генотипической самоорганизации, например, вида, рода, семейства, отряда животного или растительного царства, а ген Се3∙ ℮o-ген фенотипической самоорганизации, квантового перехода с эмерджентными свойствами, выбора пути эволюционного развития собственно системы. Он же ! ген связи с окружающей средой.
Открытие Базового Генома Мира позволило сформулировать Закон Творения, который гласит: творение всех систем Мира происходит посредством волновой энергоинформационной передачи Базового Генома Мира.
6. В развитии любой системы существуют зоны отрицательной (—Сае) и положительной (+Сае) активности. Эти зоны вычислены с помощью разработанного нами математического инструментария и показаны на огромном фактическом материале. Так, Ньютон, Лейбниц, Ломоносов, Эйлер, Лавуазье, Планк, Эйнштейн и др. сделали свои выдающиеся открытия в высоко активных зонах. Установлено, что все известные нам войны, революции начинались в зонах отрицательной активности. В этих же зонах отрицательной активности происходили природные катаклизмы, техногенные катастрофы. Наоборот, в положительных зонах природные стихии, войны затихали и, как правило, не возникали или же возникали статистически реже.
Заключая эту краткую лекцию краткий обзор о базовых положениях теории генетического энергоинформационного единства Мира, укажем на её фундаментальные характеристики:
– всеобщность и универсальность как возможность применения к любым областям жизни,
– системность как возможность использования единого энергоинформационного алгоритма для изучения любых систем Мира,
–инструментальность как применимость Законов Мира, естественно, включая Главный Закон Мира – Закон творения, к раскрытию сущности любых явлений и построению будущих схем развития систем, отвечающих правилам и требованиям Законов мира,
– научность как строгую обоснованность любых научных положений, что в огромное число раз ускоряет развитие наук и позволяет практически безошибочно продвигаться к познанию истины,
– методологичность, т. е. предоставление человечеству системы принципов для построения методологий мышления и деятельности, познания, управления, воспитания, образования, стратегического планирования, прогнозирования в различных областях жизни общества,
–идеологичность как совокупность новейших отечественных эволюционных идей, позволяющих сориентировать ментальность человечества на оптимальные, природосообразные, а поэтому соответствующие естественным законам достижения.
Перейдём к теории стратегического прогнозирования, которая создана на базе теории генетического единства Мира.
Стратегический прогноз – это заблаговременное суждение (за один и более месяцев) о предстоящих событиях, их развитии и исходе.
Назову 7 базовых положений теории стратегического прогноза.
1) На нашу планету, как и на Солнечную систему в целом, одновременно действует множество космических тел. Всем известно имя выдающегося русского учёного А.Л.Чижевского – основателя гелиобиологии. Он впервые показал, что 11-летний солнечный цикл влияет на инфекционную заболеваемость, поведение людей, на различные исторические, социально-политические события.
Нами впервые установлено, что ещё более мощное влияние на земные стихийные и социальные события оказывают другие космические системы и их взаимные циклы.
2) На эволюционную цикличность систем, если они живут в соответствии с Всеобщими Законами Мира, внешние воздействия не оказывают влияние.
3) Состояние жизни на планете зависит от её нахождения в той или иной точке цикла собственной эволюционной спирали и других космических эволюционных спиралей.
4) Особенности состояния системы в прошлом, настоящем и будущем могут быть рассчитаны посредством создания фрактальной математической модели, представленной в виде эволюционного конуса, с расчётами узловых точек фаз 1-го, 2-го, 3-го порядков.Энергетичность квантовых точек 4-го и т. д. порядков резко снижается, вследствие чего они не влияют существенным образом на события на Земле.
5) В каждой точке эволюционного конуса взаимодействуют гравитационная (творяще-созидающая) и антигравитационная (стимулирующая) энергии.
6) Главной узловой точкой является завершающая полный эволюционный цикл высшая точка эволюционного конуса. По значимости затем следуют узловые точки переходов в 4-ю, 3-ю и 2-ю фазы.
7) Прогнозы имеют две составляющие. Это – даты ожидаемых катастрофических событий, рассчитанные в соответствии с расшифрованными космическими циклами, и содержание этих событий. Наш прогноз на основании Базового Генома Мира позволяет указывать наиболее сложные периоды в жизни нашей планеты. Привязка событий к различным географическим зонам возможна по результатам частоты всевозможных катастрофических событий в них.
Некоторые прогнозы на 2003 — 2013 годы
В 2001-2002 гг. нами были рассчитаны сложнейшие периоды в жизни планеты на 2003 г. Это 4 и 26 мая и 13 ноября 2003 г. Расчёты оказались столь точными, что совпали день в день, хотя периодичность космических циклов составляла сотни миллионов и сотни миллиардов лет [5, c. 28-29].
Прогнозы на предстоящие события изложены в таблице.
Стратегический прогноз на 2007-2013 гг.
| Дата квантового перехода | Прогноз глобальный | Прогноз для России | Эволюционные циклы |
| 31.01.2007 г. | Резкое усиление солнечной активности в 2006-м – январе 2007 г. с последующим потеплением климата на Земле в эти и последующие годы. Сбои в электромагнитных сетях и приборах в декабре 2006 г. – январе2007 г. | Тот же, что и глобальный прогноз. Высоко вероятно, что в значительной степени снизятся негативные процессы в обществе. | Спираль созвездия Ориона длительностью цикла 2,56 триллиона лет |
| 23.12.2012 г. | Множественные землетрясения, цунами, вулканические взрывы с возможным помутнением атмосферы вследствие обильных выбросов вулканического пепла («вулканическая зима»), ураганы, снегопады, ливни, наводнения, пожары. Сбои в электромагнитных сетях и приборах примерно за 1,5-1 месяц до квантового перехода. | Для центральной европейской части России наиболее возможно благоприятный, но в случае «вулканической зимы» её проявления будут и на территории России. | Онтологическая спираль Земли длительностью 4817 миллионов лет |
| 27.10.2013 г. | Возможен поворот земной оси на 180˚ со смещением северного полюса в сторону северо-запада Канады и Аляски.Стихийные события те же, что и в 2012 году. Сбои в электромагнитных сетях и приборах примерно за 1,5-1 месяц до квантового перехода. | В центральной европейской части России оптимальные условия для жизнедеятельности человека. | Прецессионная спираль Земли длительностью 25977 лет |
Заключить доклад хочу поэтическими строчками:
Россия – Родина моя –
Славна великими сынами –
Покрышкин, Жуков и Толстой,
Суворов,Глинка и Чайковский,
Матросов, Павлов, Циолковский,
Чижевский, Мусоргский, Островский,
Маресьев, Пушкин и Ушинский,
Карелин,Власов, Жаботинский,
Кутузов, Лермонтов, Флоренский,
Вавилов, Козырев, Павловский,
Поленов, Репин, Менделеев,
Жуковский, Королёв, Гагарин –
Да разве перечислишь всех,
Кто нашу Родину прославил,
Кто уберёг её просторы,
Её богатства от разора,
Кто Счастье Родины творил,
Кого мы все боготворим!
Именно в России, продолжая великие традиции русского народа, открыты Базовый Геном Мира, создана Теория генетического энергоинформационного единства Мира[1-5], Периодические системы Всеобщих Законов Мира [6] и Общих законов человеческого общества [7]. На этой наукоёмкой основе разработана и апробирована система природосообразного здоровьесберегающего образования [8]. Этот филогенетический и онтогенетический интегральный потенциал России даёт основание полагать, что она – основа будущего мира на планете.
Однако всему своё время. «Ибо истинно говорю вам: доколе не прейдёт небо и земля, ни одна иота или ни одна черта не прейдёт из закона, пока не исполнится всё». (Мат: 5, 18).[2]
Библиография
1. Astafiev Boris A. The constants (codes) of evolution of life’s systems (Constants)// International information intellectual novelty registration chamber/ Certificate-Licence. Registration № EIW 000187. Cipher 00041. Code 0005. January 29,1996. Series MO. Register № 000087.
2. АстафьевБ.А.ТеорияЕдинойЖивойВселенной (законы, гипотезы). –М.: Информациология, 1997. –148 с.
3. Астафьев Б.А. Основы Мироздания. – М.: Белые альвы, 2002. – 320 с.
4. Астафьев Б.А. Всеобщий Закон Творения. –М.: Институт холодинамики,2004. – 144 с.
5. Астафьев Б.А. Стратегический прогноз и управление на основе Генома Мира: теория и практика. – М.: Институт холодинамики, 2005. – 168 с.
6. Маслова Н.В. Периодическая система Всеобщих Законов Мира. – М.: Институт холодинамики, 2005. – 184 с.
7. Маслова Н.В. Периодическая система Общих законов человеческого общества. – М.: Институт холодинамики, 2006. – 296 с.
8. Маслова Н.В. Ноосферное образование. – М.: Институт холодинамики, 2002. 2-е изд. – 342 с.
[2] А стихом ранее сказано: «Не думайте, что Я пришёл нарушить закон или пророков; не нарушитьпришёл Я, но исполнить (Мат::5, 17».
Источник: cosmatica.org
Основные законы наследования[править]
Зарождение и развитие генетики: доменделевский период[править]
Важнейшие закономерности наследования были открыты чешским ученым Г.Менделем в опытах на растениях. Но сначала нужно коротко рассказать о результатах, полученных его предшественниками. Еще в середине XVIII в. ботаники перешли от наблюдений за наследованием признаков растений к экспериментальному его изучению. В 1760 г. И. Г. Кельрейтер, который часть жизни работал в России и был российским академиком, провел серию опытов по изучению передачи признаков при скрещивании растений. В опытах с табаком, дурманом и гвоздиками Кельрейтор показал, что после переноса пыльцы одного растения на пестик другого образуются семена, из которых вырастают растения-потомки, часто имеющие признаки, промежуточные между признаками растений-родителей. Он также обнаружил, что этот результат не зависит от того, с какого из родительских растений берется пыльца (то есть равноправие «отца» и «матери» в передаче признаков потомкам). Экспериментами Кельрейтера было показано существование пола у растений.
Но особенно важно то, что Кельрейтер ввел в науку новый метод изучения наследственности — метод искусственной гибридизации. При искусственном перенесении пыльцы с цветка одного сорта на пестик цветка другого сорта получается растение, происходящее от двух сортов сразу. Такое растение называется гибридом. При этом отцовское растение — это то, с которого взята пыльца, а материнское — то, которое этой пыльцой опылили и на котором созревают гибридные семена. Растения, выросшие из этих семян, ученые называют гибридами первого поколения. Используя этот метод, французские ботаники О.Саржэ и Ш.Ноден в середине XIX в., работая на семействе тыквенных, открыли явление доминантности. Скрещивая растения разных сортов с различающимися признаками, они наблюдали, что в первом гибридном поколении часто у всех потомков проявляются признаки только одного из родителей. Эти признаки, которые как бы «побеждают» признаки другого родителя, назвали доминантными (от лат. доминантис — господствующий). Сарже, Ноденом и другими учеными было обнаружено, что все гибриды первого поколения похожи друг на друга. Это наблюдение впоследствии стали называть правилом единообразия гибридов первого поколения. При этом часть признаков гибриды получают от одного сорта, а часть — от другого. Так что часть доминантных признаков потомок получал от отца, а часть — от матери.
А что же происходит с «подавленными» признаками, которые не проявляются у гибридов первого поколения (позднее их назвали рецессивными от лат. рецессус — отступление). Исчезают ли они совсем? Оказывается, нет. Если скрещивать гибриды первого поколения между собой, то их потомки, гибриды второго поколения, отличаются по своим признакам друг от друга. Возникновение такого разнообразия признаков называют расщеплением . При этом часть гибридов второго поколения имеет те признаки, которые были у родителей исходных сортов и которые не проявлялись в первом гибридном поколении. Таким образом, эти признаки не исчезали, а лишь «маскировались» доминантными признаками. Все эти постепенно накапливающиеся факты требовали своего осмысления. Что и было сделано в работах Грегора Менделя.
Открытие законов наследственности Менделем[править]
Личность и биография Г. Менделя[править]
Грегор Мендель — чешский ученый. В его работах, выполненных в период с 1856 по 1863 г., были открыты основные законы наследственности. Иоганн Мендель родился в 1822 г. в семье крестьянина в Силезии (Австрийская империя). Окончив гимназию, он в 1843 г. был пострижен в монахи августинского монастыря в Брюнне (ныне — г. Брно в Чехии), приняв при этом имя Грегор. На средства этого монастыря он учился в Венском университете (1851—1853). Вернувшись в Брюнн, преподавал математику, физику и биологию (естественную историю) в школе. К этому периоду относятся опыты Менделя по гибридизации сортов гороха (1856—1863). В 1865 году по результатам своих работ он в два приема (8 февраля и 8 марта) сделал доклад на заседании Брюннского общества естествоиспытателей, а также опубликовал работу «Опыты над растительными гибридами» в трудах этого общества. В 1868 г. Мендель стал настоятелем монастыря и отошел от занятий наукой.
Работа Менделя «Опыты над растительными гибридами»[править]
Методология и методика Менделя — ключ к успеху[править]
Некоторые важные факты были накоплены до Менделя. В чем же состояло отличие его работ от работ предшественников?
Во-первых, Мендель сумел правильно выделить и поставить задачу, которую ему предстояло решать. Он решил посмотреть, как наследуются отдельные признаки.
Во-вторых, Мендель сумел наметить и провести трудоемкий эксперимент. Очень важный этап в постановке эксперимента — выбор подходящего объекта . Выбор гороха был не случаен. Горох легко выращивать, у него имеется много сортов, потомство от скрещивания которых хорошо размножается. Из 34 сортов гороха, бывших в его распоряжении, Мендель выбрал 22 «хороших» сорта, четко отличающихся по каким-либо признакам. Мендель работал с чистыми линиями гороха, разработав и применив на нем метод перекрестного опыления . Ученым было тщательно исследовано свыше 10000 растений и десятки тысяч семян, у которых изучались особенности наследования семи разных признаков. На рис. показаны примеры признаков гороха такого рода. Основные экспериментальные результаты Менделя, полученные при скрещивании растений, различающихся по одному признаку, приведены в таблице .
В-третьих, Мендель использовал точные количественные методы при анализе своих экспериментальных данных, что и позволило ему получить результаты, на основании которых он открыл законы наследственности. Зная теорию вероятностей (Мендель прямо пишет о ней в своей работе), он делает вывод о необходимости анализа большого числа объектов для устранения случайных отклонений. Именно с этим выводом и связан анализ столь большого числа растений.
Моногибридное скрещивание и его закономерности[править]
Первоначально Мендель скрещивал сорта, различающиеся одним изучаемым признаком (семена желтые — семена зеленые).
Скрещивание организмов двух чистых линий, различающихся по проявлениям одного изучаемого признака, за которые отвечают аллели одного гена, называется моногибридным скрещиванием.
При этом Мендель получил следующие результаты:
Р ж х з
F1 ж х ж
F2 3/4 ж : 1/4 з
Итак, в первом поколении гибридов проявлялось правило доминирования и единообразия: все гибриды имели признак одного из родителей. (Мендель проводил реципрокные скрещивания — брал пыльцу от разных родительских сортов в разных опытах — и убедился, что их результаты одинаковы.) Часто этот результат (единообразие гибридов первого поколения) называют первым законом Менделя. Сам Мендель «законов» не формулировал и не нумеровал. Однако данный результат был хорошо известен еще до работ Менделя.
«Первым законом Менделя» логичнее было бы называть закон расщепления: при моногибридном скрещивании во втором поколении гибридов в случае полного доминирования наблюдается расщепление по фенотипу в соотношении 3:1 : около 3/4 гибридов второго поколения имеют доминантный признак, около 1/4 — рецессивный.
Генетическая терминология и символика[править]
Ген — 1) дискретный наследственный фактор, определяющий проявления данного признака; 2) участок ДНК, кодирующий одну молекулу РНК.
Аллели — разновидности одного и того же гена, расположенные в идентичных локусах гомологичных хромосом.
Гомозигота — организм, который имеет два одинаковых аллеля данного гена (АА, аа).
Гетерозигота — организм, который имеет два разных аллеля данного гена (Аа).
Доминантный аллель — проявляется (определяет признак) и у гомозиготных, и у гетерозиготных по этому аллелю особей.
Рецессивный аллель — проявляется (определяет признак) только у гомозиготных по этому аллелю особей, в отсутствие доминантного аллеля.
Генотип — 1) совокупность всех генов данной особи; 2) сочетание аллелей данного гена у данной особи.
Фенотип — 1) совокупность всех признаков данной особи; 2) проявление данного признака у данной особи.
Моногибридное скрещивание — скрещивание чистых линий, различающихся одним изучаемым признаком, за который отвечают аллели одного гена (АА х аа).
Дигибридное скрещивание — скрещивание чистых линий, различающихся двумя изучаемыми признаками, за которые отвечают аллели двух генов (ААВВ х ааbb) (аналогично — тригибридное. тетрагибридное, полигибридное скрещивание).
Теория наследственности Менделя[править]
Важнейшая часть теории наследственности Менделя — гипотеза чистоты гамет, или закон чистоты гамет:
В каждую гамету попадает только один аллель из пары с вероятностью 50 %.
При генотипе Аа 50 % гамет будут нести аллель А, 50 % гамет — аллель а. Гаметы «чисты» от второго аллеля.
(По области применимости «закон чистоты гамет» — самый общий из всех «законов Менделя»).
Ди- и тригибридное скрещивание[править]
Статистический характер законов наследования. Условия выполнения законов Менделя[править]
Закон расщепления выполняется при определенных условиях:
Многочисленность потомства
Скрещиваются чистые линии
Слияние гамет неизбирательно
Все генотипы имеют равную плодовитость и жизнеспособность (не действует естественный отбор)
За каждый признак действительно отвечает один ген
Признак не сцеплен с полом (отвечающий за него ген не находится в половой хромосоме).
Судьба открытия Менделя. Переоткрытие законов Менделя[править]
Наука развивается закономерно. Когда в ней накапливается достаточное количество фактов, идеи некоторых опытов, обобщений или законов начинают «носиться в воздухе» и нередко практически одновременно открываются несколькими учеными в разных странах. Недаром так много законов и правил носит «двойные имена» (закон Био-Савара, закон Джоуля- Ленца, закон Бойля-Мариотта и т. д.). Два физика одновременно открыли ультрафиолетовые лучи, два химика в один и тот же год открыли кислород и т. д.
Яркий пример закономерности развития науки дает и возникновение генетики. Годом ее истинного рождения часто считают 1900-й, когда три ботаника, работавшие в разных странах, одновременно пришли в своих исследованиях к выводам, сходным с выводами Менделя, а при подготовке своих результатов к публикации наткнулись на его работу. Эти ботаники — Гуго де Фриз из Голландии, который работал с маком и другими растениями; Карл Корренс из Германии, работавший с кукурузой; Эрих Чермак из Австрии, который, как и Мендель, работал с горохом. Независимо от них английский генетик Уильям Бэтсон в 1898 г. вел опыты по скрещиванию кур и пришел к тем же выводам. Работа Бэтсона вышла из печати в 1902 г. В том же году появилось сообщение французского зоолога Гено о скрещивании серых мышей с альбиносами, в результате которого в первом поколении были получены только серые мыши, а во втором — 198 серых особей и 72 альбиноса. Таким образом, была доказана применимость законов Менделя к животным. После переоткрытия законов Менделя их подтвердили на огромном количестве разнообразных признаков многих животных и растений. Заметим, что и сам Мендель ставил опыты не только на горохе. Он проверил справедливость своих выводов на фасоли, левкое, кукурузе и ночной красавице.
Ряд ученых просто повторили опыты Менделя на очень большом материале и подтвердили правильность его выводов. Так, например, были повторены опыты на горохе с расщеплением по отдельным признакам. Приведем результат только для расщепления по желтой и зеленой окраске семян. Бэтсон в 1905 г. на 16000 семян получил 75,3 % доминантных (желтых) и 24,7 % рецессивных (зеленых). Дэрбишер в 1909 г., рассмотрев 145000 семян, получил расщепление в отношении 75,1 % к 24,9 %. Вообще, начиная с 1900 г., опыты по гибридизации получили очень большой размах. Прежде всего, были изучены признаки, относящиеся к окраске и форме разных организмов, так как эти признаки легко различимы невооруженным глазом.
Приведем несколько примеров первых работ по изучению расщепления признаков. Было показано выполнение законов Менделя для цвета гусениц и цвета коконов шелкопряда. Установили, что отсутствие полос на раковине виноградной улитки доминирует над их присутствием, что длинная шерсть у ангорских кроликов — рецессивный признак по отношению к короткой шерсти, а отсутствие рогов у крупного рогатого скота — доминантный признак, и т. д.
Было обнаружено, что законы Менделя применимы не только к морфологическим, но и к физиологическим признакам. Так, Корренс (1904) показал на белене, что двухлетность доминирует над однолетностыо. Н. И. Вавилов показал, что невосприимчивость растений к заболеваниям, вызываемым грибками, наследуется по законам Менделя.
Позднее сходные результаты были получены и для животных. Например, было показано, что мыши чистой линии, выведенные в Принстонском университете, невосприимчивы к ряду вирусных заболеваний, поражающих нервную систему (к вирусу желтой лихорадки, вирусу энцефалита и др.). Мыши другой линии при заражении этими вирусами погибали в 100 % случаев. Гибриды этих двух линий все были устойчивы к болезни. А из гибридов второго поколения 1/4 погибала, а 3/4 были невосприимчивы к заболеваниям.
Как ясно из приведенных примеров, уже при возникновении генетики началось изучение генетических особенностей ряда хозяйственно важных объектов и признаков, имеющих большое практическое значение.
Законы Менделя и наследование у человека[править]
У человека давно известен ряд внешних признаков, которые наследуются по Менделю. Например, курчавые или волнистые волосы доминируют над прямыми. Темный цвет глаз доминирует над голубым. Такие признаки давно использовались для определения родства (или как доказательство невозможности родства). Так, например, в романе Агаты Кристи «Рождество Эркюля Пуаро» знаменитый сыщик по цвету глаз определяет, что одна из героинь в действительности не является внучкой хозяина поместья. Пуаро говорит ей: «Мадемуазель, если бы вы изучали законы Менделя, вам было бы известно, что в семье, где у обоих родителей голубые глаза, не бывает кареглазых детей».
- Кстати, тут Пуаро ошибся. Бывает, что у родителей с голубыми глазами и очень небольшим содержанием темного пигмента в радужной оболочке, рождаются кареглазые дети.
Один из дефектов зрения, так называемая врожденная куриная слепота , — сильное снижение зрения в сумерках — наследуется как доминантный признак. В одной французской семье этот дефект был прослежен на протяжении 10 поколений.
Таким образом, в соответствии с законом расщепления наследуются самые разные признаки — в том числе и такие, которые имеют важное значение для сельского хозяйства и медицины.
Развитие классической генетики в ХХ веке[править]
Множественные аллели[править]
Неполное доминирование и кодоминирование[править]
Уровни рассмотрения признака и относительность понятия доминирования[править]
Летальные аллели. Условные летали[править]
Приводят к смерти особей. Примером могут служить большинство рецессивных аллелей гена Y (yellow) у мышей, которые в гетерозиготном состоянии дают желтую окраску шерсти, а в гомозиготном приводят к гибели эмбрионов до рождения.
Школа Моргана. Создание хромосомной теории наследственности[править]
Вскоре после открытия Менделя (в 1870—1890) были описаны митоз и мейоз, индивидуальные хромосомы и их поведения в ходе митоза и мейоза, оплодотворение как слияние клеток и последующее слияние пронуклеусов, сформировалось понятие кариотипа как стабильного видового признака.
В 1883 г ван Бенеден установил, что число хромосом в соматических клетках у животных вдвое больше, чем в половых
В 1900 г произошло переоткрытие законов Менделя.
Т.Бовери доказал, что для нормального развития зародышей морского ежа необходимы все хромосомы; открыл диминуцию хроматина у аскариды и ее связь с дифференцировкой клеток.
Наконец, в 1902—1903 г была высказана гипотеза о связи между хромосомами и менделевскими факторами (генами) (У.Сэттон, Т.Бовери).
Всё это послужило предпосылками для создания в 1910-е годы хромосомной теории наследственности. Основная заслуга в её развитии принадлежит Томасу Гент Моргану (Нобелевская премия 1936 г) и сотрудникам его лаборатории, которые в качестве основного объекта использовали плодовую мушку дрозофилу.
Диминуция хроматина у аскариды и гипотеза Бовери[править]
Диминуция, то есть утрата кусков хромосом, у аскариды происходит в предшественниках всех клеток, кроме половых
Параллелизм между поведением хромосом и поведением менделевских факторов[править]
Не может не броситься в глаза параллелизм в поведении хромосом и менделевских факторов (аллелей):
За каждый признак В соматических клетках имеются отвечает пара аллелей, пары гомологичных хромосом, один из которых получен от одна из которых получена от отца, отца, другой - от матери другая - от матери
При образовании гамет При образовании гамет из пары аллелей данного из пары гомологичных хромосом гена в гамету попадает в гамету попадает только один только одна
Аллели, которые При образовании гамет гомологичные находятся в разных парах хромосомы разных пар гомологичных хромосом, распределяются по ним независимо наследуются (расщепляются) (случайно в любых сочетаниях) независимо
Видимо, первым, кто детально проанализировал этот параллелизм и на основании этого высказал предположение о том, что гены находятся в хромосомах, был американский генетик Уильям Сэттон (Саттон).
Открытие хромосомного определения пола[править]
Дрозофила как объект генетических исследований[править]
Сцепленное наследование и группы сцепления[править]
Открытие кроссинговера. Трехфакторные скрещивания и линейность расположения генов в хромосомах[править]
Цитологические доказательства кроссинговера[править]
Генетический анализ кроссинговера дает возможность построить модель хромосомы. В 30-х годах в лаборатории Т. Моргана, Ф. Г. Добржанский начал сопоставлять генетическую и цитологическую карты дрозофилы. Он показал, что обе карты сходны, их элементы параллельно чередуются: определенные диски гигантских хромосом и гены в группах сцепления. Большинство генов располагается в участках эухроматина. В Y-хромосоме очень мало генов, и она почти целиком состоит из гетерохроматина. Первые прямые цитогенетические доказательства прохождения кроссинговера были получены в 1931 г. Г. Крейтоном и Б. Мак-Клинток на кукурузе и К. Штерном на дрозофиле. Штерн добился создания гетероморфности для пары XX-хромосом у дрозофилы, используя транслокацию одной хромосомы из пары на XY-хромосому, для другого гомолога транслокацию с четвертой хромосомой (микрохромосомой). В первом случае он получил комплексную хромосому, состоящую из всего тела хромосомы и половины Y-хромосомы. Эта хромосома имела необычную форму, отличающую ее от нормальной Х-хромосомы и от других хромосом дрозофилы. Она приобрела Г -образную форму вместо обычной прямой, и в ней был расположен доминирующий ген красных глаз cr+ и рецессивный круглых глаз B+, , а вторая часть прикрепилась к маленькой четвертой хромосоме, благодаря чему не терялась при редукционном делении, что было важно для сохранения жизнеспособности таких особей. Самки с разными по форме Х-хромосомами были скрещены самцами, имеющими обычные прямые Х-хромосомы, в которых находились рецессивные гены cr окраски глаз (цвета гвоздики) и круглой их формы В+, то есть самцы были с круглыми глазами гвоздичного цвета. В результате этого скрещивания были получены самки четырех типов: с полосковидными глазами цвета гвоздики, получившие от матери разделенную на две части Х-хромосому; кругло- и красноглазые, пучившие от матери Г — образную хромосому; в результате кроссинговера — круглоглазые с глазами гвоздичного цвета, у которых должна быть короткая изогнутая Х-хромосома с прикрепленным к ней участком Y-хромосомы, и с полосковидными красными глазами . От отца же все самки должны были получить прямую Х-хромосому. И действительно, цитологическое исследование самок всех четырех типов подтвердило, что у 369 самок из 374 строение хромосом соответствовало предсказанному на основании их признаков, и только в пяти случаях отмечалось несоответствие, что можно было объяснить двойным кроссинговером (Иванова О. А., 1974). Весь опыт состоял в том, что у мух, для которых факт кроссинговера регистрировался чисто генетически, затем изучались комплексы их хромосом. Во всех случаях прохождение генетического кроссинговера сопровождалось предсказанным изменением в морфологии хромосом. Хромосомная теория наследственности Т. Моргана состоит в следующем:
- гены находятся в хромосомах и расположены в линейной последовательности на определенных расстояниях друг от друга;
- гены, расположенные в одной хромосоме, составляют группу сцепления. Число групп сцепления соответствует гаплоидному числу хромосом;
- признаки, гены которых находятся в одной хромосоме, наследуются сцепленно, так как их гены передаются в половые клетки чаще в тех же сочетаниях, в которых они были в хромосомах исходных родительских форм;
- в потомстве гетерозиготных родителей новые сочетания генов, расположенных в одной паре хромосом, могут возникать в результате кроссинговера в процесс е мейоза. Частота кроссинговера зависит от расстояния между генами;
- кроссинговер бывает одинарным, двойным и множественным. Одинарный кроссинговер происходит чаще, чем двойной и тройной;
- вследствие интерференции один обмен препятствует осуществлению другого обмена вблизи места, где обмен уже произошел;
- учитывая линейное расположение генов в хромосоме и частоту кроссинговера как показателя расстояния между генами, можно построить карты хромосом;
- за единицу расстояния между генами принята частота кроссинговера (сантиморган, сМ).
Генетическое картирование. Построение первых генетических карт[править]
Современные методы генетического картирования. Построение генетической карты хромосом человека[править]
Основные положения хромосомной теории наследственности[править]
В результате изучения генетики пола, сцепленного наследования и кроссинговера, Морганом и его сотрудниками были сформулированы основные положения хромосомной теории наследственности. (Они в большинстве учебников четко не формулируются, число их может сильно различаться.)
- Гены находятся в хромосомах.
- Каждая хромосома содержит несколько (множество) генов.
- Гены, расположенные в одной хромосоме, наследуются совместно (сцепленно) и образуют группу сцепления.
- Хромосомы сохраняют свою целостность и индивидуальность в течение всего клеточного цикла, в связи с этим группы сцепления постоянны.
- Число групп сцепления равно гаплоидному числу хромосом данного вида (у гетерогаметного пола больше на одну).
- Сцепление аллелей, расположенных в одной хромосоме, может нарушаться за счет кроссинговера.
- Гены расположены в хромосомах в линейной последовательности.
- Частота кроссинговера (нарушения сцепления) пропорциональна расстоянию между генами.
Зарождение и развитие биохимической генетики[править]
Биохимическая генетика-раздел генетики, изучающий механизмы генетического контроля биохимических процессов в живых организмах. Развитие биохимической генетики стало возможным после выяснения в 40—50-х гг. 20 в., что в генах содержится информация о точном строении ферментов, управляющих всеми процессами в организме, то есть что наследственная программа реализуется совокупностью биохимических процессов, содержание и скорость течения которых определяются генами.
Мысль о том, что реализация генетической (наследственной) информации может заключаться в обеспечении как организма в целом, так и каждой отдельной клетки сведениями о времени и порядке осуществления определенных реакций, высказывалась еще в 19 в. Напр., амер. биолог Э. Вильсон писал, что «наследственность — это повторение в последовательных поколениях одинаковых форм обмена». Однако доказательство этого стало возможным лишь в результате развития биохимии и изучения биохимич. основ жизнедеятельности животных, растений и микроорганизмов, которое позволило достаточно подробно представить сложную картину генетического контроля обмена веществ и выявить последовательность синтеза отдельных соединений в организме.
Одним из основных методов исследования в 40—60-е гг. было изучение измененных (мутантных) форм различных микроорганизмов, нуждающихся в дополнительном поступлении каких-либо питательных веществ или с измененной чувствительностью к различным веществам (в том числе ядам), нарушающим обмен веществ. Позднее более широкое распространение получил метод условно летальных мутаций: изучались организмы с наследственными изменениями, обусловливающими возможность осуществления каких-либо жизненно важных процессов только в строго определенных условиях (напр., при определенной температуре), без соблюдения которых они погибали.
Особенно важное значение исследования в области биохимической генетики имеют для изучения наследственности человека, и в частности для развития медицинской генетики. Установлено, что первопричиной многочисленных наследственных нарушений обмена веществ, многих болезней крови и кроветворной системы, нервной системы, пороков развития и т. п. являются изменения в структуре или активности генов и связанные с этим нарушения процессов синтеза тех или иных ферментов, управляющих определенными биохимическими реакциями.
Зарождение и развитие медицинской генетики[править]
Задачи по генетике[править]
1. У матери группа крови А, у отца — О. Во всех ли случаях можно переливать кровь обоих родителей их детям? Ответ обоснуйте.
2. При скрещивании серого кролика-самца с гималайскими и белыми самками все потомство было серым, а при скрещивании другого серого самца с теми же самками половина потомства было серым, половина — гималайским. Объясните этот результат и определите генотипы упомянутых в условии кроликов исходя из того, что за эти типы окраски отвечает один ген.
3. Редиски бывают белые, розовые и красные; круглые, овальные и длинные. За каждый из признаков (цвет и форму) отвечают два аллеля одного гена. Предложите механизм наследования этого признака и предскажите, каким будет расщепление от скрещивания розовой овальной редиски и красной овальной.
4. Дальтоник — носитель гена серповидноклеточной анемии женится на женщине — носительнице генов обоих этих заболеваний. Каким будет расщепление по фенотипу среди их потомства?
5. У мышей рецессивный аллель с подавляет проявление других генов окраски (мыши с генотипом сс — альбиносы, аллель С не влияет на окраску), а аллель А в гетерозиготном состоянии обуславливает желтую окраску, а в гомозиготном летален. Каким будет расщепление по цвету в потомстве от скрещивания СсАа х СсАа? Укажите также фенотипы родителей.
6. У цветного горошка при наличии доминантных аллелей С и Р окраска цветков красная, при отсутствии хотя бы одного из них — белая. Каким будет расщепление по цвету от скрещивания СсРр х Ссрр?
7. У прыгунчиков есть две независимо наследуемые рецессивные мутации — одна из них (а) отвечает за хлопанье ушами, а вторая (в) — за их редукцию (такую сильную, что хлопать уже нечем). У нормальных прыгунчиков уши ослиные. Каким будет расщепление по фенотипу от скрещивания АаВв х АаВв?
8. У бракозявров ген А отвечает за фермент, превращающий зеленое вещество в более яркое желтое, а ген В — за фермент, превращающий желтое вещество в еще более яркое красное. Окраска определяется самым ярким из присутствующих веществ, рецессивные аллели дают неработающие ферменты .
9. Гены А, В и С у бракозявров разными путями превращают зеленый пигмент в красный (если он присутствует — окраска красная, отсутствует — зеленая). Каким будет расщепление по цвету от скрещивания ААВвСс х ааВвсс?
10. У ананасов есть три типа листьев — с шипами (Ш), с шипами только на верхушке (ШВ) и без шипов (БШ). Скрещивание чистых линий дало следующие результаты
Р_____ ; F1; F2
ШВ х Ш ; ШВ; 3ШВ : 1 Ш
БШ х ШВ; БШ; 3БШ : 1ШВ
БШ х Ш ; БШ; 12БШ : 3ШВ : 1 Ш
Объясните эти результаты и определите генотипы родительских линий.
11. У дрозофилы есть две независимо наследуемые рецессивные мутации. Одна из них вызывает скручивание крыльев, а другая — их недоразвитие (причем степень редукции крыльев такова, что скручиваться уже нечему). Есть две чистые линии мух, одна из которых имеет скрученные крылья, а другая — недоразвитые. Какое расщепление по фенотипу будет в F1 и в F2 от скрещивания этих линий?
12. У бракозявров ген А кодирует фермент, который превращает зеленое вещество 1 в желтое вещество 2, гены В и С — ферменты, превращающие желтое вещество 2 в красное вещество 3. Рецессивные аллели дают неактивные формы ферментов. Красное в-во ярче желтого, желтое ярче зеленого. Окраска определяется самым ярким веществом. Каким будет расщепление по окраске в F1 при скрещивании двух тригетерозигот?
13. Курицу с оперенными ногами и розовидным гребнем скрестили с голоногим петухом, имевшим гороховидный гребень. В потомстве от этого скрещивания были получены цыплята с оперенными ногами и гороховидным гребнем, с оперенными ногами и ореховидным гребнем, с оперенными ногами и розовидным гребнем и с оперенными ногами и простым гребнем в соотношении 1:1:1:1. Объясните эти результаты и укажите генотипы исходных птиц.
14. Частота встречаемости всех генотипов по группам крови АВО в данной популяции равная, вероятность образования любых пар генотипов родителей тоже равная. Какова вероятность того, что у матери с IV группой крови (АВ) родится ребенок с III группой (В) ?
15. При скрещивании серого кролика Коли с белым Василисой получено все потомство серое, а при скрещивании сестры Коли, серого кролика Маши, с гималайским кроликом Петей — 1/2 серых и 1/2 гималайских. При скрещивании Пети с Василисой все потомки оказались гималайскими. а) Считая, что эти типы окраски определяются тремя аллелями одного гена, обозначьте генотипы всех упомянутых особей; укажите, какой аллель над каким доминирует. б) Какое получится расщепление, если всех потомков от скрещивания Маши и Пети скрестить с Машей?
16. В норме у бракозявров закладываются длинные пальцы вместе с межпальцевыми тканями. Затем включается программа гибели межпальцевых клеток — пальцы отделяются друг от друга. Мутация р+ нарушает программу гибели клеток (при этом у взрослого бракозявра пальцы соединены перепонками). При мутации v+ пальцы редуцируются полностью. Нормальные аллели — р- и v-. Каким будет фенотипическое расщепление в F2 от скрещивания p+p+v+v+ x p-p-v-v-, если доминантными являются а) p+ и v+ ? б) p+ и v- ?
17. У крыс окраска определяется генами А и В следующим образом А-В- — серая А-вв — желтая ааВ- — черная аавв- кремовая Для проявления этих генов необходимо присутствие аллеля С, особи сс — альбиносы. Все эти гены наследуются независимо. Потомство от скрещивания четырех разных линий крыс-альбиносов с гомозиготной серой линией было серым. В F2 наблюдались следующие расщепления Линия Серые Желтые Черные Кремовые Альбиносы альбиносов 1 3/4 — — — 1/4 2 9/16 3/16 — — 4/16 3 9/16 — 3/16 — 4/16 4 27/64 9/64 9/64 3/64 1/4 Определите генотипы родительских линий альбиносов (1 и 2 или 3 и 4 по выбору). Приведите ход рассуждений.
18. При скрещивании дрозофил с изогнутыми крыльями между собой в потомстве наблюдалось расщепление 2/3 изогнутые : 1/3 нормальные, а при скрещивании изогнутые х нормальные — расщепление 1/2 изогнутые : 1/2 нормальные. Объясните этот результат.
19. У рыбки с механизмом определения пола ХУ получены такие результаты наследования окраски: 1) белая самка х красный самец — все потомки F1 красные, в F2 половина самок красные, половина белые, все самцы красные; 2) красный самец из F1 первого скрещивания х белая самка — в F1 все самки белые, все самцы красные; 3) красная самка х белый самец — в F1 все красные, в F2 самки все красные, половина самцов красные, половина белые. Объясните эти результаты, предположив, как наследуется окраска.
20. Родители имеют группы крови А и В, а частоты всех генотипов по генам системы АВО в данной популяции равны. Какова вероятность рождения у них ребенка с группой крови О?
21. В Киевском зоопарке была выведена порода умных и красивых обезьян. При скрещивании этих обезьян с обычными в поколении F1 все обезьяны были обычными. В F2 наблюдалось соотношение фенотипов 45 обычных:15 красивых:3 умных:1 красивая и умная а) Предложите модель наследования ума и красоты.
б) Каким будет расщепление при скрещивании гибрида F1 с умной и красивой обезьяной?
(Автор задачи — А. Г. Козленко)
22. Ген А подавляет ген В, ген В подавляет ген С и активирует ген D. Рецессивные аллели не активны. При активном гене С окраска красная (красный цвет ярче зеленого), при отсутствии активности С и активности D окраска зеленая, в остальных случаях — белая. Каким будет расщепление по цвету от скрещивания АаBbCcDd x AaBbCcDd?
Источник: ru.wikibooks.org