Химический состав хроматина
Хроматин – основной компонент клеточного ядра – достаточно легко получить из выделенных интерфазных ядер и из выделенных митотических хромосом. Для этого используют его свойство переходить в растворенное состояние при экстракции водными растворами с низкой ионной силой или просто деионизованной водой. При этом участки хроматина набухают и переходят в гель. Чтобы такие препараты перевести в настоящие растворы, необходимы сильные механические воздействия: встряхивание, перемешивание, дополнительная гомогенизация. Это, конечно, приводит к частичному разрушению исходной структуры хроматина, дробит его на мелкие фрагменты, но практически не меняет его химического состава.
Фракции хроматина, полученные из разных объектов, обладают довольно однообразным набором компонентов. Было найдено, что суммарный химический состав хроматина из интерфазных ядер и митотических хромосом мало отличаются друг от друга. Главными компонентами хроматина являются ДНК и белки, среди которых основную массу составляют гистоны и негистоновые белки (см табл. 3).
Таблица 3. Химический состав хроматина. Содержание белков и РНК дано по отношению к ДНК
|
Источник хроматина |
ДНК |
Гистоны |
Негистоновые белки |
РНК |
|
Стебель зародыша гороха |
1,0 |
1,03 |
0,29 |
0,26 |
|
Печень крысы |
1,0 |
1,16 |
0,67 |
0,043 |
|
Клетки Hela (опухоль человека) |
1,0 |
1,02 |
0,71 |
0,09 |
|
Тимус теленка |
1,0 |
1,14 |
0,33 |
0,007 |
|
Эритроциты курицы |
1,0 |
1,08 |
0,54 |
0,02 |
В среднем в хроматине около 40% приходится на ДНК и около 60 % на белки, среди которых специфические ядерные белки-гистоны, составляют от 40 до 80% от всех белков, входящих в состав выделенного хроматина. Кроме того в состав хроматиновой фракциии входят мембранные компоненты, РНК, углеводы, липиды, гликопротеиды. Вопрос о том, насколько эти минорные компоненты входят в структуру хроматина еще не решен. Так, например, РНК может представлять собой транскрибируемую РНК, которая еще не потеряла связь с матрицей ДНК. Другие же минорные компоненты могут представлять собой вещества соосажденных фрагментов ядерной оболочки.
В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, ассоциированной с гистонами (см.
с. 57). Поэтому укоренилось другое название хроматина – нуклеогистон. Именно за счет ассоциации гистонов с ДНК образуются очень лабильные, изменчивые нуклеиново-гистоновые комплексы, где отношения ДНК : гистон равно примерно единице, т.е. они присутствуют в равных весовых количествах. Эти нитчатые фибриллы ДНП и есть элементарные хромосомные или хроматиновые нити, толщина которых в зависимости от степени упаковки ДНК может колебаться от 10 до 30 нм. Эти фибриллы ДНП могут в свою очередь дополнительно компактизоваться с образованием более высоких уровней структуризации ДНП, вплоть до митотической хромосомы. Роль некоторых негистоновых белков заключается именно в образовании высоких уровней компактизации хроматина.
ДНК хроматина
В препарате хроматина на долю ДНК приходится обычно 30-40%. Эта ДНК представляет собой двухцепочечную спиральную молекулу подобно чистой выделенной ДНК в водных растворах. Об этом говорят многие экспериментальные данные. Так, при нагревании растворов хроматина наблюдается повышение оптической плотности раствора, так называемый гиперхромный эффект, связанный с разрывом межнуклеотидных водородных связей между цепями ДНК, подобно тому, что происходит при нагревании (плавлении) чистой ДНК.
Вопрос о размере, длине молекул ДНК в составе хроматина имеет важное значение для понимания структуры хромосомы в целом. При стандартных методах выделения ДНК хроматина обладает молекулярной массой 7-9 х 106, что значительно меньше молекулярной массы ДНК из кишечной палочки (2,8 х 109).
кую сравнительно малую молекулярную массу ДНК из препаратов хроматина можно объяснить механическими повреждениями ДНК в процессе выделения хроматина. Если же выделять ДНК в условиях, исключающих встряхивание, гомогенизацию и другие воздействия, то удается из клеток получить молекулы ДНК очень большой длины. Длина молекул ДНК из ядер и хромосом эукариотических клеток может быть изучена с помощью метода светооптической радиоавтографии, подобно тому как это изучалось на прокариотических клетках.
Было обнаружено, что в составе хромосом длина индивидуальных линейных ( в отличие от прокариотических хромосом) молекул ДНК может достигать сотен микрометров и даже нескольких сантиметров. Так, у разных объектов были получены молекулы ДНК от 0,5 мм до 2 см. Эти результаты показали, что есть близкое совпадение между расчетной длиной ДНК на хромосому и радиоавтографическим наблюдением.
После мягкого лизиса клеток эукариот можно прямо определять молекулярные массы ДНК физико-химическими методами. Было показано, что максимальная молекулярная масса молекулы ДНК дрозофилы равна 41 х 109, что соответствует длине около 2 см. У некоторых дрожжей на хромосому приходится молекула ДНК с молекулярной массой 1 х 108-109, которая имеет размеры около 0,5 мм.
Такие длинные ДНК представляют собой одну молекулу, а не несколько более коротких, сшитых гуськом с помощью белковых связок, как считали некоторые исследователи. К этому заключению пришли после того, как оказалось, что длина молекул ДНК не изменяется после обработки препаратов протеолитическими ферментами.
Общее количество ДНК, входящее в ядерные структуры клеток, в геном организмов, колеблется от вида к виду, хотя у микроорганизмов количество ДНК на клетку значительно ниже, чем у беспозвоночных, высших растений и животных. Так, у мыши на ядро приходится почти в 600 раз больше ДНК, чем у кишечной палочки. Сравнивая количество ДНК на клетку у эукариотических организмов, трудно уловить какие-либо корреляции между степенью сложности организма и количеством ДНК на ядро. Примерно одинаковое количество ДНК имеют такие различные организмы как лен, морской еж, окунь (1,4-1,9 пг) или рыба голец и бык (6,4 и 7 пг).
Значительны колебания количества ДНК в больших таксономических группах. Среди высших растений количество ДНК у разных видов может отличаться в сотни раз, так же, как и среди рыб, в десятки раз отличается количество ДНК у амфибий.
У некоторых амфибий в ядрах количество ДНК больше, чем в ядрах человека в 10-30 раз, хотя генетическая конституция человека несравненно сложнее, чем у лягушек. Следовательно, можно предполагать, что «избыточное» количество ДНК у более низко организованных организмов либо не связано с выполнением генетической роли, либо число генов повторяется то или иное число раз.
Таблица 4. Содержание ДНК в клетках некоторых объектов (пг, 10-12 г)
|
Объект |
Количество ДНК в пг |
|
Бактериофаги Т1 Т4 Бактерии кишечная палочка Грибы дрожжи нейроспора Высшие растения лен кукуруза лилия Беспозвоночные животные дрозофила сверчок домашний Рыбы осетр протоптерус Амфибии тритон обыкновенный амфиума Пресмыкающиеся черепаха зеленая Птицы курица Млекопитающие мышь человек
|
Количество ДНК на частицу 0,000007 0,000027 количество ДНК на клетку 0,009 количество ДНК на гаплоидную клетку 0,027 0,017 количество ДНК на диплоидную клетку 1,4 15,4 134,2
0,2 12,0
3,2 100,0
73,0 108,0
5,0
2,3
5,0 6,0 |
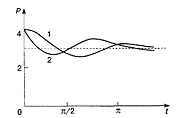
Разрешить эти вопросы оказалось возможным на основании изучения кинетики реакции ренатурации или гибридизации ДНК. Если фрагментированные молекулы ДНК в растворах подвергнуть тепловой денатурации, а затем инкубировать их при температуре несколько более низкой, чем та, при которой происходит денатурация, то идет восстановление исходной двуспиральной структуры фрагментов ДНК за счет воссоединения комплементарных цепей – ренатурация.
я ДНК вирусов и прокариотических клеток было показано, что скорость такой ренатурации прямо зависит от величины генома; чем больше геном, чем больше количество ДНК на частицу или клетку, тем больше нужно времени для случайного сближения комплементарных цепей и специфической реассоциации большего числа разных по нуклеотидной последовательности фрагментов ДНК (рис. 53). Характер кривой реассоциации ДНК прокариотических клеток указывает на отсутствие повторяющихся последовательностей оснований в геноме прокариот; все участки их ДНК несут уникальные последовательности, число и разнообразие которых отражает степень сложности генетической композиции объектов и, следовательно, их общей биологической организации.
Совсем другая картина реассоциации ДНК наблюдается у эукариотических организмов. Оказалось, что в состав их ДНК входят фракции, которые ренатурируют с гораздо более высокой скоростью, чем можно было бы предполагать на основании размера их генома, а также фракция ДНК, ренатурирующая медленно, подобно уникальным последовательностям ДНК прокариот. Однако для эукариот требуется значительно большее время для ренатурации этой фракции, что связано с общим большим размером их генома и с большим числом различных уникальных генов.
В той части ДНК эукариотов, которая отличается высокой скоростью ренатурации, различают две подфракции: 1) фракцию с высоко или часто повторяющимися последовательностями, где сходные участки ДНК могут быть повторены 106 раз; 2) фракцию умеренно повторяющихся последовательностей, встречающихся в геноме 102-103 раз.
к, у мыши во фракцию ДНК с часто повторяющимися последовательностями входит 10% от общего количества ДНК на геном и 15% приходится на фракцию с умеренно повторяющимися последовательностями. Остальные 75% от всей ДНК мыши представлены уникальными участками, соответствующими большому числу различных неповторяющихся генов.
Фракции с часто повторяющимися последовательностями могут обладать иной плавучей плотностью, чем основная масса ДНК, и поэтому могут быть выделены в чистом виде, как так называемые фракции сателлитной ДНК. У мыши эта фракция имеет плотность, равную 1,691 г/мл, а основная часть ДНК — 1,700 г/мл. Эти различия плотности определяются различиями в нуклеотидном составе. Например, у мыши в этой фракции имеется 35% Г и Ц пар, а в основном пике ДНК — 42%.
Как оказалось, сателлитная ДНК, или фракция ДНК с часто повторяющимися последовательностями, не участвует в синтезе основных типов РНК в клетке, не связана с процессом синтеза белка. Этот вывод сделан был на основании того, что ни один из типов РНК клетки (тРНК, иРНК, рРНК) не гибридизируется с сателлитными ДНК. Следовательно, на этих ДНК нет последовательностей, отвечающих за синтез клеточных РНК, т.е. сателлитные ДНК не являются матрицами для синтеза РНК, не участвуют в транскрипции.
Существует гипотеза о том, что высокоповторяющиеся последовательности, не участвующие непосредственно в синтезе белков, могут нести информацию, играющую важную структурную роль в сохранении и функционировании хромосом. К ним могут быть отнесены многочисленные участки ДНК, связанные с белками остова интерфазного ядра (см. ниже), участки начала репликации или транскрипции, а также участки ДНК, регулирующие эти процессы.
Методом гибридизации нуклеиновых кислот прямо на хромосомах (in situ) была изучена локализация этой фракции. Для этого на изолированной сателлитной ДНК с помощью бактериальных ферментов синтезировали меченую 3Н-уридином РНК. Затем цитологический препарат с хромосомами подвергали такой обработке, при которой происходит денатурация ДНК (повышенная температура, щелочная среда и др.). После этого на препарат помещали меченную 3Н РНК и добивались гибридизации между ДНК и РНК. Радиоавтографически было обнаружено, что большая часть метки локализуется в зоне первичных перетяжек хромосом, в зоне их центромерных участков. Метка обнаруживалась также и в других участках хромосом, но очень слабо (рис. 54).
За последние 10 лет сделаны большие успехи в изучении центромерных ДНК, особенно у дрожжевых клеток. Так у S. cerevisiae центромерная ДНК состоит из повторяющихся участков по 110 п.н. Она состоит из двух консервативных участков (I и III) и центрального элемента (II), обогащенного АТ-парами оснований. Сходное строение ДНК центромеры имеют хромосомы дрозофилы. Центромерная ДНК человека (альфоидная сателлитная ДНК) состоит из тандема мономеров по 170 п.н., организованных в группы димеров или пентамеров, которые в свою очередь образуют большие последовательности по 1-6 х 103 п.н. Такая самая большая единица повторена 100-1000 раз. С этой специфической центромерной ДНК комплексируются особые центромерные белки, участвующие в образовании кинетохора, структуры, обеспечивающей связь хромосом с микротрубочками веретена и в движении хромосом в анафазе (см. ниже).
ДНК с высокоповторяющимися последовательностями обнаружена также в теломерных участках хромосом многих эукариотических организмов (от дрожжей до человека). Здесь чаще всего встречаются повторы, в которые входят 3-4 гуаниновых нуклеотида. У человека теломеры содержат 500-3000 повторов TTAGGG. Эти участки ДНК выполняют особую роль — ограничивать хромосому с концов и предотвращать ее укорачивание в процессе многократной репликации.
Недавно было найдено, что высокоповторяющиеся последовательности ДНК интерфазных хромосом связываются специфически с белками — ламинами, подстилающими ядерную оболочку, и участвуют в заякоревании растянутых деконденсированных интерфазных хромосом, тем самым определяют порядок в локализации хромосом в объеме интерфазного ядра.
Сделано предположение, что сателлитная ДНК может участвовать в узнавании гомологичных районов хромосом при мейозе. По другим предположениям, участки с часто повторяющимися последовательностями играют роль разделителей (спейсеров) между различными функциональными единицами хромосомной ДНК, например между репликонами (см. ниже).
Как оказалось, фракция умеренно повторяющихся (от 102 до 105 раз) последовательностей принадлежит к пестрому классу участков ДНК, играющих важную роль в процессах создания аппарата белкового синтеза. В эту фракцию входят гены рибосомных ДНК, которые могут быть повторены у разных видов от 100 до 1000 раз. В эту фракцию входят многократно повторенные участки для синтеза всех тРНК. Более того, некоторые структурные гены, ответственные за синтез определенных белков, также могут быть многократно повторены, представлены многими копиями. Такими являются гены для белков хроматина — гистонов, повторяющихся до 400 раз.
Кроме того, в эту фракцию входят участки ДНК с разными последовательностями (по 100-400 нуклеотидных пар), также многократно повторенными, но рассеянными по всему геному. Их роль еще не до конца ясна. Высказывается предположение, что такие участки ДНК могут представлять собой акцепторные или регуляторные участки разных генов.
Итак, ДНК эукариотических клеток гетерогенна по составу, содержит несколько классов последовательностей нуклеотидов: часто повторяющиеся последовательности (> 106 раз), входящие во фракцию сателлитной ДНК и не транскрибирующиеся; фракция умеренно повторяющихся последовательностей (102-105), представляющих блоки истинных генов, а также короткие последовательности, разбросанные по всему геному; фракция уникальных последовательностей, несущая информацию для большинства белков клетки.
Исходя из этих представлений становятся понятными те различия в количестве ДНК, которые наблюдаются у разных организмов: они могут быть связаны с неодинаковой долей тех или иных классов ДНК в геноме организмов. Так, например, у амфибии Amphiuma (у которой ДНК в 20 раз больше, чем у человека) на долю повторяющихся последовательностей приходится до 80% от всей ДНК, у луков — до 70, у лосося — до 60% и т.п. Истинное же богатство генетической информации должна отображать фракция уникальных последовательностей. Не нужно забывать, что в нативной, нефрагментированной молекуле ДНК хромосомы все участки, включающие уникальные, умеренно и часто повторяющиеся последовательности, связаны в единую гигантскую ковалентную цепь ДНК.
Молекулы ДНК гетерогенны не только по участкам разной нуклеотидной последовательности, но и различны в отношении их синтетической активности.
Репликация эукариотических ДНК
Бактериальная хромосома реплицируется как одна структурная единица, имеющая одну стартовую точку репликации и одну точку терминации. Таким образом бактериальная циклическая ДНК является одним репликоном. От стартовой точки репликация идет в двух противоположных направлениях, так что по мере синтеза ДНК образуется так называемый глазок репликации, ограниченный с двух сторон репликационными вилками, что хорошо видн при электронномикроскопическом изучении вирусных и бактериальных реплицирующихся хромосом.
У эукариотических клеток организация репликации иного характера – полирепликоннная.. Как уже говорилось, при импульсном включении 3НТ множественная метка появляется практически во всехмитотических хромосомах. Это означает, что одновременно в интерфазной хромосоме существует множество мест репликации и множество автономных точек начала репликации. Более подробно это явление было изучено с помощью радиоавтографии меченых молекул, выделенных ДНК (рис. 55).Если клетки были импульсно мечены 3НТ, то в световом микроскопе на автографах выделенных ДНК можно видеть участки восстановленного серебра в виде пунктирных линий. Это небольшие отрезки ДНК, которые успели реплицироваться, а между ними расположены участки нереплицированной ДНК, которая не оставила радиоавтографа и поэтому остается невидимой. По мере увеличения времени контакта 3НТ с клеткой величина таких отрезков возрастает, а расстояние между ними уменьшается. Из этих экспериментв можно точно рассчитать скорость репликации ДНК у эукариотических организмов. Скорость движения репликационной вилки оказалась равной 1-3 т.п.н. в мин у млекопитающих, около 1 т.п.н. в мин у некоторых растений, что намного ниже скорости репликации ДНК у бактерий (50 т.п.н. в мин.). В этих же экспериментах была прямо доказана полирепликонная структура ДНК хромосом эукариот: по длине хромосомной ДНК, вдоль нее, располагается множество независимых участков репликации – репликонов. По расстоянию между средними точками смежных метящихся репликонов, т.е. по расстоянию между двумя соседними стартовыми точками репликации, можно узнать величину отдельных репликонов. В среднем величина репликонову высших животных составляет около 30 мкм или 100 т.п.н. Следовательно, в гаплоидном наборе млекопитающих должно быть 20 000-30 000 репликонов. У низших эукариот величина репликонов меньше, около 40 т.п.н. Так у дрозофилы на геном приходится 3500 репликонов, а у дрожжей – 400. Как говорилось, синтез ДНК в репликоне идет в двух противоположных направлениях. Это легко доказывается радиоавтографически: если клеткам после импульсной метки дать продолжить синтезировать ДНК некоторое время в среде без 3НТ, то произойдет падение включения его в ДНК, будет происходить как бы разбавление метки, и на радиоавтографе можно будет видеть симметричное, с двух сторон реплицируемого участка, уменьшение количества зерен восстановленного серебра.
Реплицирующиеся концы или вилки в репликоне прекращают движение, когда встретятся с вилками соседних репликонов (в терминальной точке, общей для соседних репликонов). В этом месте реплицированные участки соседних репликонов объединяются в единые ковалентные цепи двух новосинтезированных молекул ДНК. Функциональное подразделение ДНК хромосом на репликоны совпадает со структурным подразделением ДНК на домены или петли, основания которых, как уже упоминалось, скреплены белковыми связками.
Таким образом весь синтез ДНК на отдельной хромосоме протекает за счет независимого синтеза на множестве отдельных репликонов, с последующим соединением концов соседних отрезков ДНК. Биологический смысл этого свойства становится ясным при сравнении синтеза ДНК у бактерий и эукариот. Так бактериальная монорепликонная хромосома длиной в 1600 мкм синтезируется со скоростью около получаса. Если бы сантиметровая молекула ДНК хромосомы млекопитающих реплицировалась тоже как монорепликонная структура, то на это ушло бы около недели (6 суток). Но если в такой хромосоме расположено несколько сот репликонов, то для полной ее репликации понадобится всего около часа. На самом же деле время репликации ДНК у млекопитающих составляет 6-8 часов. Это связано с тем, что не все репликоны отдельной хромосомы включаются одновременно.
В некоторых случаях наблюдается одновременное включение всех репликонов или же появление дополнительных точек начала репликации, что дает возможность закончить синтез всех хромосом за минимально короткое время. Это явление происходит на ранних этапах эмбриогенеза некоторых животных. Так известно, что при дроблении яиц шпорцевых лягушек Xenopus laevis синтез ДНК занимает всего 20 минут, тогда как в культуре соматических клеток этот процесс продолжается около суток. Аналогичная картина наблюдается у дрозофилы: на ранних эмбриональных стадиях весь синтез ДНК в ядре занимает 3,5 минуты, а в клетках культуры ткани – 600 минут. При этом в клетках культуры величина репликонов оказалась почти в 5 раз больше, чем у эмбрионов.
Синтез ДНК по длине отдельной хромосомы происходит неравномерно. Было обнаружено, что в индивидуальной хромосоме активные репликоны собраны в группы, репликативные единицы, которые включают в себя 20-80 точек начала репликации. Это следовало из анализа радиоавтографов ДНК, где наблюдалась именно такая сблоченность реплицирующихся отрезков. Другим основанием для представления о существовании блоков или кластеров репликонов или репликационных единиц были эксперименты с включением в ДНК аналога тимидина — 5’-бромдезоксиуридина (BrdU). Включение BrdU в интерфазный хроматин приводит к тому, что во время митоза, участки с BrdU конденсируются в меньшей степени (недостаточная конденсация), чем те участки, где включался тимидин. Поэтому те участки митотических хромосом в которые включился BrdU, будут слабо окрашиваться при дифференциальной окраске. Это позволяет на синхронизированных культурах клеток выяснить последовательность включения BrdU, т.е. последовательность синтеза ДНК по длине одной взятой хромосомы. Оказалось, что происходит включение предшественника в большие участки хромосомы. Включение разных участков происходит строго последовательно в течение S-периода. Каждая хромосома характеризуется высокой стабильностью порядка репликации по своей длине, имеет свой специфический рисунок репликации.
Кластеры репликонов, объединенные в репликационные единицы, связаны с белками ядерного матрикса (см. ниже), которые вместе с ферментами репликации образуют т.н. кластеросомы – зоны в интерфазном ядре, в которых идет синтез ДНК.
Порядок, в котором активируются репликационные единицы, может, вероятно, определяться структурой хроматина в этих участках. Так, например, зоны конститутивного гетерохроматина (вблизи центромеры) реплицируются обычно в конце S-периода, также в конце S-периода удваивается часть факультативного гетерохроматина (например, X-хромосома самок млекопитающих). Особенно четко во времени последовательность репликации участков хромосом коррелирует с рисунком дифференциальной окраски хромосом: R-сегменты относятся к ранореплицирующимся, G-сегменты соответствуют участкам хромосом с поздней репликацией. C-сегменты (центромера) – места самой поздней репликации.
Так как в разных хромосомах величина и число разных групп дифференциально окрашенных сегментов различно, то это создает картину асинхронного начала и завершения репликации разных хромосом в целом. Во всяком случае, последовательность начала и окончания репликации отдельных хромосом в наборе не беспорядочная. Существует строгая последовательность репродукции хромосом относительно других хромосом в наборе.
Длительность процесса репликации отдельных хромосом прямо не зависит от их размеров. Так крупные хромосомы человека группы А (1-3) оказываются мечеными в течение всего S-периода, так же как и более короткие хромосомы группы В (4-5).
Таким образом, синтез ДНК в геноме эукариот начинается почти одновременно на всех хромосомах ядра в начале S-периода. Но при этом происходит последовательное и асинхронное включение разных репликонов как в разных участках хромосом, так и в разных хромосомах. Последовательность репликации того или иного участка генома строго детерминирована генетически. Это последнее утверждение доказывается не только картиной включения метки в разные отрезки S-периода, но также тем, что существует строгая последовательность появления в ходе S-периода пиков чувствительности определенных генов к мутагенам.
Источник: StudFiles.net
Химический состав хромосом
Физико-химическая организация хромосом эукариотической клетки
Изучение химической организации хромосом эукариотических клеток показало, что они состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин, получивший свое название за способность окрашиваться основными красителями.
Как было доказано многочисленными исследованиями (см. § 3.2), ДНК является материальным носителем свойств наследственности и изменчивости и заключает в себе биологическую информацию — программу развития клетки, организма, записанную с помощью особого кода. Количество ДНК в ядрах клеток организма данного вида постоянно и пропорционально их плоидности. В диплоидных соматических клетках организма ее вдвое больше, чем в гаметах. Увеличение числа хромосомных наборов в полипловдных клетках сопровождается пропорциональным увеличением количества ДНК в них.
Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки.
Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Являясь положительно заряженными основными белками, они достаточно прочно соединяются с молекулами ДНК, чем препятствуют считыванию заключенной в ней биологической информации. В этом состоит их регуляторная роль. Кроме того, эти белки выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах (см. разд. 3.5.2.2).
Число фракций негистоновых белков превышает 100. Среди них ферменты синтеза и процессинга РНК, редупликации и репарации ДНК. Кислые белки хромосом выполняют также структурную и регуляторную роль. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.
РНК хромосом представлена отчасти продуктами транскрипции, еще не покинувшими место синтеза. Некоторым фракциям свойственна регуляторная функция.
Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» списывания информации с молекулы ДНК.
Массовые соотношения ДНК: гистоны: негистоновые белки: РНК: липиды — равны 1:1:(0,2—0,5):(0,1—0,15):(0,01—-0,03). Другие компоненты встречаются в незначительном количестве.
Сохраняя преемственность в ряду клеточных поколений, хроматин в зависимости от периода и фазы клеточного цикла меняет свою организацию. В интерфазе при световой микроскопии он выявляется в виде глыбок, рассеянных в нуклеоплазме ядра. При переходе клетки к митозу, особенно в метафазе, хроматин приобретает вид хорошо различимых отдельных интенсивно окрашенных телец — хромосом.
Интерфазную и метафазную формы существования хроматина расценивают как два полярных варианта его структурной организации, связанных в митотическом цикле взаимопереходами. В пользу такой оценки свидетельствуют данные электронной микроскопии о том, что в основе как интерфазной, так и метафазной формы лежит одна и та же элементарная нитчатая структура. В процессе электронно-микроскопических и физико-химических исследований в составе интерфазного хроматина и метафазных хромосом были выявлены нити (фибриллы) диаметром 3,0—5,0, 10, 20—30 нм. Полезно вспомнить, что диаметр двойной спирали ДНК составляет примерно 2 нм, диаметр нитчатой структуры интерфазного хроматина равен 100—200, а диаметр одной из сестринских хроматид метафазной хромосомы — 500— 600 нм.
Наиболее распространенной является точка зрения, согласно которой хроматин (хромосома) представляет собой спирализованную нить. При этом выделяется несколько уровней спирализации (компак-тизации) хроматина (табл. 3.2).
Таблица 3.2. Последовательные уровни компактизации хроматина
| Фибрилла | Степень укорочения | Диаметр, нм | |
| по сравнению с предшествующей структурой | по сравнению с молекулой ДНК | ||
| ДНК | 1—2 | ||
| Нуклесомная нить | |||
| Элементарная хроматиновая фибрилла | 20—30 | ||
| Интерфазная хромонема | 100—200 | ||
| Метафазная хроматида | 500—600 |

Рис. 3.46. Нуклеосомная организация хроматина.
А — деконденсированная форма хроматина;
Б — электронная микрофотография эукариотического хроматина:
А — молекула ДНК накручена на белковые коры;
Б — хроматин представлен нуклеосомами, соединенными линкерной ДНК
Нуклеосомиая нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) (рис. 3.46).
Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки.
Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом (рис. 3.46, Б). В связи с этим геном человека, состоящий из 3 · 109 п. н., представлен двойной спиралью ДНК, упакованной в 1,5 · 107 нуклеосом.
Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области, расположенные с интервалами в несколько тысяч пар нуклеотидов, играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.
В результате нуклеосомной организации хроматина двойная спираль ДНК диаметром 2 нм приобретает диаметр 10—11 нм.
Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная, возможно, по типу соленоида. Такая Хроматиновая фибрилла, называемая также элементарной, имеет диаметр 20—30 нм (рис. 3.47).
| |
Рис. 3.47. Хроматиновая фибрилла диаметром 20—30 нм. А — соединение соседних нуклеосом с помощью гистона HI; Б — цепочка, образуемая нуклеосомами разделенными участками ДНК, свободными от белковых тел; В — возможная модель упаковки ДНК в хроматиновой фибрилле в виде соленоида |
Интерфазная хромонема. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы (рис. 3.48). Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки Хроматиновая фибрилла диаметром 20—30 нм преобразуется в структуру диаметром 100—200 нм, называемую интерфазной хромонемой.
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией (рис. 3.49). Они выявляются в интерфазном ядре в виде глыбок хроматина. Возможно, существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в метафазных хромосомах, что используют в цитогенетических исследованиях (см. разд. 3.5.2.3 и 6.4.3.6).
Неодинаковая степень компактизации разных участков интерфазных хромосом имеет большое функциональное значение. В зависимости от состояния хроматина выделяют эухроматиновые участки хромосом, отличающиеся меньшей плотностью упаковки в неделящихся клетках и потенциально транскрибируемые, и гетерохроматиновые участки, характеризующиеся компактной организацией и генетической инертностью. В их пределах транскрипции биологической информации не происходит.
Различают конститутивный (структурный) и факультативный гетерохроматин.
Конститутивный гетерохроматин содержится в околоцентромерных и теломерных участках всех хромосом, а также на протяжении некоторых внутренних фрагментов отдельных хромосом (рис. 3.50). Он образован только нетранскрибируемой ДНК. Вероятно, его роль заключается в поддержании общей структуры ядра, прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности.
Рис. 3.49. Структурные блоки в организации хроматина.
А — петельная структура хроматина;
Б — дальнейшая конденсация хроматиновых петель;
В — объединение петель, имеющих сходную структуру, в блоки с образованием окончательной формы интерфазной хромосомы
Рис. 3.50. Конститутивный гетерохроматин в метафазных хромосомах человека
Примером факультативного гетерохроматина служит тельце полового хроматина, образуемое в норме в клетках организмов гомогаметного пола (у человека гомогаметным является женский пол) одной из двух Х-хромосом. Гены этой хромосомы не транскрибируются. Образование факультативного гетерохроматина за счет генетического материала других хромосом сопровождает процесс клеточной дифференцировки и служит механизмом выключения из активной функции групп генов, транскрипция которых не требуется в клетках данной специализации. В связи с этим рисунок хроматина ядер клеток из разных тканей и органов на гистологических препаратах различается. Примером может служить гетерохроматизация хроматина в ядрах зрелых эритроцитов птиц.
Перечисленные уровни структурной организации хроматина обнаруживаются в неделящейся клетке, когда хромосомы еще недостаточно компактизованы, чтобы быть видимыми в световой микроскоп как отдельные структуры. Лишь некоторые их участки с более высокой плотностью упаковки выявляются в ядрах в виде хроматиновых глыбок (рис. 3.51).
Рис. 3.51. Гетерохроматин в интерфазном ядре
Компактные участки гетерохроматина сгруппированы около ядрышка и ядерной мембраны
Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе (см. разд. 2.4.2). В телофазе митоза происходит декомпак-тизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Описанная митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Степень компактизации хроматина в разные периоды митотического цикла клетки можно оценить по данным, приведенным в табл. 3.2.
Источник: studopedia.su
| Обратная связь ПОЗНАВАТЕЛЬНОЕ Сила воли ведет к действию, а позитивные действия формируют позитивное отношение Как определить диапазон голоса — ваш вокал Как цель узнает о ваших желаниях прежде, чем вы начнете действовать. Как компании прогнозируют привычки и манипулируют ими Целительная привычка Как самому избавиться от обидчивости Противоречивые взгляды на качества, присущие мужчинам Тренинг уверенности в себе Вкуснейший «Салат из свеклы с чесноком» Натюрморт и его изобразительные возможности Применение, как принимать мумие? Мумие для волос, лица, при переломах, при кровотечении и т.д. Как научиться брать на себя ответственность Зачем нужны границы в отношениях с детьми? Световозвращающие элементы на детской одежде Как победить свой возраст? Восемь уникальных способов, которые помогут достичь долголетия Как слышать голос Бога Классификация ожирения по ИМТ (ВОЗ) Глава 3. Завет мужчины с женщиной
Оси и плоскости тела человека — Тело человека состоит из определенных топографических частей и участков, в которых расположены органы, мышцы, сосуды, нервы и т.д.
|
Главная количественная особенность генетического материала эукариот – наличие избыточной ДНК. Этот факт легко выявляется при анализе отношения числа генов к количеству ДНК в геноме бактерий и млекопитающих. Около 95% генома бактерий состоит из кодирующих последовательностей. Остальные 5%, по-видимому, заняты регуляторными элементами. Иная картина наблюдается у эукариотических организмов. Например, у человека насчитывают приблизительно 50 тысяч генов (имеется в виду только суммарная длина кодирующих участков ДНК – экзонов). В то же время размер генома человека 3×109 (три миллиарда) п.н. Это означает, что кодирующая часть его генома составляет всего 15 — 20 % от тотальной ДНК. Существует значительное число видов, геном которых в десятки раз больше генома человека, например некоторые рыбы, хвостатые амфибии, лилейные. Избыточная ДНК характерна для всех эукариот. По химическому составу хроматин представляет собой комплекс ДНК и белков. В ДНК эукариот выделяют 3 фракции: уникальные последовательности – у животных составляют 40-90% от всей ядерной ДНК; у растений – 12-60%. У человека около 50% ядерной ДНК представлено уникальными последовательностями длиной 1-2 тпн. Эти фрагменты разделены между собой умеренно повторяющимися последовательностями длиной 0,1-0,3 тпн. Большая часть уникальной ДНК является «некодирующей», так, например, в геноме человека из 50% уникальной ДНК на кодирующие участки приходится только 5%. Умеренно повторяющиеся последовательности ДНКкодируют, как правило, тРНК, рРНК, гистоны. Пример, гены 18S и 28S рРНК образуют тандемные пары (мономерные единицы расположены по типу «голова к хвосту»). Спейсер тоже транскрибируется, в его состав входит ген 5,8S рРНК. У человека и мыши гены рРНК располагаются в нескольких кластерах в акроцентрических хромосомах. Каждый кластер генов рРНК соответствует ядрышковому организатору. Гены ранних гистонов у морского ежа входят в состав одной единицы, в результате повторения которой образуется кластер тандемных генов, разделяемых спейсерами – Н1 Н4 Н2В Н3 Н2А. У дрозофилы порядок расположения гистоновых генов иной – Н1 Н3 Н4 Н2А Н2В. У некоторых млекопитающих гены гистонов располагаются по отдельности. Сателлитная ДНК– высокоповторяющиеся последовательности ДНК. Располагаются в центромерных, теломерных районах и в участках интеркалярного гетерохроматина. У разных видов на долю сателлитной ДНК приходится от 0,3% до 28% от всей ядерной ДНК. Близкие виды, например мышь и крыса, имеют совершенно различные высокочастотные последовательности, у крысы их нуклеотидный состав не отличается от основной ДНК, тогда как геном мыши содержит четкий АТ-богатый сателлит. Это означает, что высокочастотная ДНК способна к быстрым изменениям в ходе видообразования. Последовательности сателлитной ДНК – это короткие тандемные повторы. У человека сателлитная ДНК делится на I, II, III классы (с длиной повторяющейся последовательности 1-20 п.н.). Также выделяют альфа, бета и гамма типы (с длиной повтора 170 п.н., 68 п.н. и 220 п.н. соответственно). Сателлитные повторы имеют различную степень обогащенности АТ- и ГЦ –пар (тип 1 обогащен парами АТ). Сателлитная ДНК специфично распределена по разным хромосома. Так, в прицентромерном гетерохроматине хромосом 3 и 4 представлен сат ДНК 1. В прицентромерном гетерохроматине хромосом групп Д и G локализованы сатДНК1 и 3 классов. Распределение альфа, бета и гамма сателлитных ДНК также хромосом специфично. Альфа сателлитные повторы располагаются в основном в центромерных участках хромосом. Имеют элементапрную единицу повтора около 170 п.н. и могут присутствовать в тандеме, достигающем длины до 1 млн.п.н. В сателлитной ДНК выделяют 2 типа последовательностей: — микросателлиты –единица повтора состоит из 1-5 нуклеотидов. Общая длина кластера – несколько десятков нуклеотидов. Пример: (АТ)n; (A)n; (AC)n. В кодирующей части генов чаще встречаются тринуклеотидные повторы. Ди-, тетра- и пентануклеотидные повторы редки в кодирующей части генома, так как увеличение их числа обязательно приведет к сдвигу рамки считывания. Микросателлитные последовательности обнаружены в геноме как прокариот, так и эукариот. Из-за высокой скорости мутирования микросателлиты обеспечивают генетическое разнообразие геномов. — минисателлиты –единица повтора состоит из 10-100 пн. Общий размер кластера 0,5-100 тпн. Общий размер кластера 0,5-100 тпн. Их еще называют VNTR – вариабельные по числу тандемные повторы. Они могут находиться внутри или между генами. Число копий каждой специфической последовательности в разных локусах варьирует от 1000 до 5000 п.н. Вариации длин таких областей лежат в основе метода ДНК-фингерпринтинга (метод отпечатков). По локализации выделяют: — сателлитные повторы в области промотора – (А)n – стимулирует экспрессию гена, т.к. обуславливает жесткую структуру двойной спирали – на этом участке не образуются нуклеосомы, что облегчает доступ ДНК-полимераз и факторов транскрипции к промотору. Длина полипролиновых, полиаланиновых и полиглутаминовых участков, кодируемых микросателлитными тринуклеотидными повторами, влияет на белок-белковые взаимодействия, в том числе и с факторами активации транскрипции. — сателлитные повторы в интронах – часто ингибируют экспрессию гена. (АС)21 в интроне гена рецептора эпидермального фактора роста снижает транскрипцию гена на 80%. — сателлитные повторы в нетранслируемой 5-области гена (между промотором и геном) – (С)n, (AC)n, (GC)n, (AT)n – ингибируют экспрессию гена. — повторы в транслируемой области гена. Пример – хорея Гентингтона – увеличение длины повтора (CAG)n в первом экзоне гена белка гентингтина приводит к удлинению полиглутаминового участка в белке. Синтезируется токсичная форма белка, что приводит к гибели нейронов. Это пример динамических мутаций или экспансии тринуклеотидных повторов – связаны с изменением длины тринуклеотидных повторов. Скорость мутирования связана с числом копий триплета. После превышения определенного порога длины эти повторы становятся нестабильными и их длина увеличивается в последующих поколениях. У здоровых людей число повторов колеблется от 6 до 39. У больных – 36-180. У взрослых болезнь проявляется при 40-55 повторах, при числе повторов больше 70 заболевание проявляется уже у детей. То есть для данных мутаций характерно явление антиципации –возрастание пенетрантности заболевания в ряду поколений, более раннее начало заболевания. Впервые динамические мутации описаны в 1991 году. Известны они только у людей. Всего пока описано 16 наследственных заболеваний (табл. 1). Все они связаны с поражением головного мозга и подкорковых структур. Экспансия происходит как в мейозе, так и в митозе, затрагивает чаще аллели с изначально большим числом повторов. Динамические мутации являются причиной развития синдрома фрагильной Х-хромосомы или синдрома Мартина-Белл. Синдром Мартина-Белл – одна из наиболее частых наследственных форм умственной отсталости (частота заболевания составляет 1 случай на 4000 мужчин и 1 случай на 8000 женщин). Синдром связан с образованием ломкого сайта в Х-хромосоме из-за увеличения повторов (CGG) в 5-нетранслируемой области гена FMR1, который экспрессируется в мозге и семенниках. Ген кодирует РНК-связывающий белок, циркулирующий в клетке между ядром и цитоплазмой. У здоровых людей ген содержит 2-54 триплета (CGG). Увеличение длины до 200 триплетов – это предмутационное состояние, не изменяющее фенотип человека. Увеличение числа копий свыше 200 триплетов – полная мутация и проявление синдрома. Увеличение числа копий триплета приводит к гиперметилированию регуляторной зоны гена (промотора), в результате резко снижается или полностью ингибируется транскрипция ген. Таблица 1 Болезни экспансии повторяющихся последовательностей
Другими наследственными заболеваниями, обусловленными динамическими мутациями, являются атаксия Фридрейха, миотоническая дистрофия первого и второго типа, болезнь Кеннеди, спиноцеребеллярная атаксия, прогрессирующая миоклонус-эпилепсия. Еще одна категория повторов в геноме эукариот – короткие и длинные разбросанные по геному ДНК-повторы (не тандемные). Короткие рассеянные по геному элементы – SINE – последовательности длиной менее 500 п.н. Их копийность в геноме человека составляет не менее 500 000. К этой группе повторов относится семейство Alu-повторов (они содержат сайты рестрикции для эндонуклеазы AluI). Alu-повторы в геноме человека составляют более 5% от суммарного количества ДНК. Функция их не ясна. Другая группа рассеянных повторов – LINE. У человека обнаружено одно семейство этих повторов – L1-повторы. Длина повтора около 6400 п.н., суммарно представлены в геноме до 100000 раз. Проявляет свойства ретротранспозонов (транскрипция последовательности, синтез на мРНК с помощью обратной транскриптазы новой копии ДНК повтора, новая копия интегрирует в хромососму на новом месте). Эухроматическая часть генома построена по принципу чередования (интерсперсии) уникальных и повторяющихся последовательностей. Условно выделяют два основных типа интерсперсии, получивших названия по тем видам, у которых они впервые были описаны: интерсперсия типа «ксенопус» и типа «дрозофила». Примерно в 50 % генома Xenopus laevis уникальные последовательности из 800-1200 п.н. чередуются с повторяющимися, средний размер которых 300 п.н. В остальной части геномов типа «ксенопус» расстояния между соседними повторами значительно превышают 1-2 п.н. Структура генома типа «ксенопус» широко распространена, особенно среди животных. Млекопитающие и человек также относятся к этому типу организации генома. Особенность генома человека и других приматов составляют интерсперсные высокочастотные повторы длиной около 300 п.н. У человека эти повторы содержат сайт, разрезаемый ферментом рестрикции Alu I. У D. melanogaster параметры интерсперсии резко отличаются от видов с типом генома «ксенопус»: повторяющиеся последовательности длиной 5600 п.н. чередуются с уникальными, длина которых не менее 13000 п.н. Интересно отметить, что у домашней мухи геном устроен по типу «ксенопус». Этот факт прямо указывает на то, что в ходе эволюции возможны очень быстрые преобразования характера чередования последовательностей и в эухроматической части генома. Птицы по параметрам интерсперсии занимают промежуточное положение между типом «ксенопус» и типом «дрозофила». Как показывают результаты исследований последних лет, многие виды животных и растений по организации генома не могут быть строго отнесены ни к тому, ни к другому типу. Так, в геномах млекопитающих встречаются длинные повторы – в несколько тысяч пар нуклеотидов, в геномах лилейных до 90% ДНК может быть представлено повторяющимися последовательностями. Например, геном гороха не содержит уникальных последовательностей, превышающих по длине 300 п.н.
Источник: megapredmet.ru |