Царство эукариот
Эукариоты — это организмы, состоящие из клеток, в которых обязательно содержится особый органоид — ядро. Иначе эукариоты называются ядерными организмами. Эукариоты объединяются в надцарство Ядерные, или Эукариоты. По современной систематике в надцарство Эукариоты входят три царства: Растения, Животные и Грибы. Рассмотрим краткую характеристику этих царств.
Краткая характеристика особенностей и биолого-экологической роли растений
Растения образуют отдельное царство организмов потому, что имеют ряд признаков, сильно отличающих их от других организмов. Такими особыми признаками являются их способность к фотосинтезу, наличие четкого чередования поколений в цикле развития, практически полное отсутствие возможности активного перемещения во внешней среде (для подавляющего большинства); почти все растения являются продуцентами, т. е. они синтезируют органические вещества из неорганических, используя солнечную энергию. Для растений можно выделить следующие общие признаки.
1. Клетки растений содержат пигмент хлорофилл, способный усваивать солнечную энергию, необходимую для протекания фотосинтеза, в результате которого растения из углекислого газа, воды и других неорганических веществ синтезируют органические вещества; клетки растений запасают органические вещества или в виде жидких растительных жиров, или в виде углевода крахмала (относится к полисахаридам).
2. Растительные клетки имеют оболочку, которая кроме мембраны, образованной белками и липидами, содержит целлюлозу, придающую особую прочность и устойчивость клеткам растений; для растений большую роль играет осмос (односторонняя диффузия, при которой в клетку поступают вода и неорганические вещества за счет того, что в клетках концентрация веществ выше, чем в окружающей среде).
3. Большинство растений прикреплены к субстрату (месту своего произрастания), что делает невозможным активное перемещение всего растения в пространстве. Это привело к тому, что растения растут в течение всей своей жизни, занимая все большее и большее пространство как под землей, так и на ее поверхности; для растений характерны ростовые движения (рост стеблей направлен к солнцу), а также движение соцветий в направлении солнца — фототропизм (подсолнечник и др.).
4. Для растений характерно вегетативное, бесполое и половое размножение, а также четкое чередование полового (гаметофит) и бесполого (спорофит) поколений.
5. Расселение растений осуществляется переносом семян или спор, которые являются фазами покоя, позволяющими растениям перенести неблагоприятные условия внешней среды.
Различают низшие и высшие растения. К низшим растениям относятся зеленые, красные и бурые водоросли и лишайники (лишайники являются «комплексными» организмами, одновременно относящимися к двум царствам — к царству Растения и царству Грибы; это симбиотические организмы).
К высшим растениям относят Моховидные, Плауновидные, Хвощевидные, Папоротниковидные, Голосеменные и Покрытосеменные растения. Главной особенностью высших растений является то, что их тело расчленено на отдельные органы, а наиболее высокоорганизованные формы растений (семенные растения) размножаются семенами.
Биолого-экологическую роль растений трудно переоценить. Без этих организмов жизнь на Земле была бы невозможной. Они производят органические вещества, являющиеся пищей для всех гетеротрофных организмов. Кроме того, они поглощают углекислый газ и выделяют молекулярный кислород (O2), без которого невозможна жизнь аэробных организмов (в том числе и человека).
Растения образуют растительные сообщества, составляющие основу всех природных сообществ, т. е. биогеоценозов. В сени лесов и других растительных сообществ животные и другие организмы находят и «стол», и «кров».
Велика роль растений и в жизни человека. Растения являются источником растительной пищи, сырьем для производства одежды, бумаги, различных технических изделий и произведений искусства (скульптур, рам для картин, художественных изделий из бересты, соломы, тканей и др.).
Особенности и биолого-экологическая роль животных
Животные образуют особое царство Животные и характеризуются рядом признаков, отличающих их от организмов других царств. Рассмотрим наиболее важные из них.
1. Все животные являются гетеротрофами, и для них характерны следующие способы питания: хищничество, паразитизм (питание живыми биоорганическими веществами), сапрофитизм и поедание детрита; их главная экологическая роль — консументы разных порядков, но целый ряд животных являются редуцентами (например, дождевой червь, являющийся детритофагом).
2. Животные запасают либо жиры, либо гликоген (животный крахмал) в отличие от растений и грибов.
3. Клетки животных отграничены от других клеток либо мембраной, либо клеточной оболочкой, которая упрочнена или хитином, или жироподобными веществами в комплексе с белковыми соединениями.
4. Большинство животных (особенно высокоорганизованные) способны к активному перемещению в пространстве, имеют нервную систему и развитые органы чувств, поэтому многие из них растут только до определенной стадии своего развития, хотя новые клетки образуются в течение всей жизни, но этот процесс связан с постоянным обновлением организма (это не относится к клеткам, образующим нервную систему).
5. Для большинства животных характерно только половое размножение, поэтому в циклах развития отсутствует строгое чередование полового и бесполого поколений (только у низкоорганизованных видов царства Животные наблюдается такое чередование поколений, например у малярийного плазмодия; низкоорганизованные животные способны и к вегетативному, и к бесполому размножению).
Велика биолого-экологическая роль животных. Они являются промежуточными звеньями в реализации круговорота веществ в природе, а некоторые из них — и завершающей (детритофаги). Животные способствуют процессам размножения растений (например, насекомые-опылители); важна их роль и в расселении растений — семена распространяются за счет прикрепления к телу животных, за счет выделения из пищеварительной системы при поедании плодов и т. д. Велика роль животных в жизни человека: они «снабжают» его пищей, сырьем для разных отраслей промышленности, являются источником эстетического наслаждения и т. д.
Особенности и эколого-биологическая роль грибов
В современной систематике грибы выделены в отдельное царство эукариотических организмов — Грибы, так как их биологические особенности сочетают в себе признаки и животных, и растений. Рассмотрим некоторые наиболее важные признаки грибов.
1. Грибы, как и животные, являются гетеротрофами, но они могут быть паразитами (например, головневые грибы), сапрофитами и детритофагами; в отличие от животных хищничество для грибов не характерно (оно отсутствует).
2. Грибы являются как одноклеточными, так и многоклеточными; их тело представляет собой грибницу (или мицелий), состоящую из переплетения тончайших нитей — гиф. Различают низшие и высшие грибы. Низшие грибы, как правило, одноклеточны (клетки у них многоядерны); к ним относят мукор, фитофторы и др. Высшие грибы многоклеточны, их клетки одно- или многоядерны; к ним относят шляпочные и другие грибы. Грибы тесно соединены с субстратом и не способны к активному перемещению в пространстве (как растения) и растут в течение всей жизни.
3. Для грибов характерно вегетативное, половое и бесполое размножение, при этом у многих видов наблюдается четкое чередование полового и бесполого поколений (как у растений).
4. Из углеводов грибы запасают гликоген (подобно животным), хотя есть грибы, у которых запасным углеводом является крахмал.
5. Клетки грибов имеют очень прочные оболочки (как у растений), но у многих грибов прочность клеточных оболочек связана с содержанием в этих оболочках хитина (как у животных).
Все перечисленное выше делает необходимым выделение царства грибов, как отдельного царства организмов. Изучением грибов занимается наука микология, но традиционно в школьном курсе грибы изучаются совместно с ботаникой (наукой о растениях), так как по большинству явных признаков грибы располагаются ближе к растениям, чем к животным.
Велика биолого-экологическая роль грибов. Паразитические грибы являются фактором эволюции, так как способствуют выживанию более приспособленных, «сильных» особей того или иного вида. Грибы-сапрофиты и детритофаги способствуют переходу органических соединений в неорганические, что приводит к нормальному ходу круговорота веществ в природе.
Значительна роль грибов в жизнедеятельности человека. Грибы-паразиты вызывают различные трудноизлечимые заболевания. Многие грибы используют в биотехнологии (например, гриб пенициллум является основой в технологии получения пенициллина, важнейшего антибиотика, применяемого при лечении многих заболеваний). Грибы применяют в кондитерской и хлебопекарной промышленности (дрожжевые грибки) в пивоваренной промышленности, при производстве алкогольных напитков. Многие грибы съедобны и употребляются в пищу. Важно помнить о существовании ядовитых грибов и уметь отличать их от съедобных. Ядовитые грибы можно использовать для изготовления лекарственных препаратов. Эти грибы применяют для создания галлюцинаций при камлании (шаманы в религиозных обрядах северных народов России, Аляски и др.).
Источник: www.polnaja-jenciklopedija.ru
Симбиогенез — основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная — симбиотическая гипотеза (симбиогенез). Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки. Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.

Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий. Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде. Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.
Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза. Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.
Источник: biology.su
Строение эукариотической клетки[править | править код]
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органеллы (другое название, реже употребляемое в научной литературе[комм. 1], — органоиды), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Деление на царства[править | править код]
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств из-за биохимических особенностей. Также некоторые авторы выделяют царства протистов или простейших и хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалье-Смита, все эукариоты подразделяются на два монофилетических таксона (поддомена) — униконтов (Unikonta) и биконтов (Bikonta). Положение таких эукариот, как Collodictyon и Diphylleia rotans, на данный момент не определено.
Отличия эукариот от прокариот[править | править код]
Важнейшая, основополагающая особенность эукариот связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие, среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд лет назад, от них около 1,2 млрд лет назад произошли эукариотические организмы.
Примечания[править | править код]
Комментарии[править | править код]
Источники[править | править код]
Литература[править | править код]
- На русском языке
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, т. 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Медников Б. М. Биология: формы и уровни жизни. — М.: Просвещение, 1995.
- Тейлор Д., Грин Н., Стаут У. Биология. В трёх томах
- Малахов В. В. Основные этапы эволюции эукариотных организмов. — 2003
- Федонкин М. А. Сужение геохимического базиса жизни и эвкариотизация биосферы: причинная связь. — 2003
- Шестаков С. В. О ранних этапах биологической эволюции с позиции геномики. — 2003
- Марков А. В. Проблема происхождения эукариот
- Марков А. В, Куликов А. М. Происхождение эвкариот: выводы из анализа белковых гомологий в трёх надцарствах живой природы
- Заварзин Г. А. Эволюция микробных сообществ.
- Колчанов Н. А. Эволюция регуляторных генетических систем.
- Розанов А. Ю., Федонкин М. А.. Проблема первичного биотопа эвкариот. — 1994.
- Богданов Ю. Ф., Дадашев С. Я., Гришаева Т. М. Сравнительная геномика и протеомика дрозофилы, нематоды Бреннера и арабидопсиса. Идентификация функционально сходных генов и белков синапсиса мейотических хромосом
- Ермилова Е. В., Залуцкая Ж. М., Лапина Т. В. Подвижность и поведение микроорганизмов. Т. 2: Эукариоты
- Греннер Д., Марри Р., Мейес П., Родуэлл В. Биохимия человека
- На других языках
- Bisby FA, Roskov YR, Ruggiero MA, Orrell TM, Paglinawan LE, et al. Species 2000 & ITIS catalogue of life: 2007 annual checklist. Species 2000. Retrieved Jan. 2007. 21, 2008
- Patterson DJ. The diversity of eukaryotes. Am Nat. 1999
- Stechmann A, Cavalier-Smith T. Rooting the eukaryote tree by using a derived gene fusion. Science. 2002
- Richards TA, Cavalier-Smith T. Myosin domain evolution and the primary divergence of eukaryotes. Nature. 2005
- Stechmann A, Cavalier-Smith T. Phylogenetic analysis of eukaryotes using heat-shock protein Hsp90. J Mol Evol. 2003
- Makiuchi T, Nara T, Annoura T, Hashimoto T, Aoki T. Occurrence of multiple, independent gene fusion events for the fifth and sixth enzymes of pyrimidine biosynthesis in different eukaryotic groups. Gene. 2007
- Kim E, Simpson AGB, Graham LE. Evolutionary relationships of apusomonads inferred from taxon-rich analyses of 6 nuclear encoded genes. Mol Biol Evol. 2006
- Nozaki H, Matsuzaki M, Misumi O, Kuroiwa H, Higashiyama T, et al. Phylogenetic implications of the CAD complex from the primitive red alga Cyanidioschyzon merolae (Cyanidiales, Rhodophyta). J Phycol. 2005
- Adl SM, Simpson AGB, Farmer MA, Andersen RA, Anderson OR, et al. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. J Eukaryot Microbiol. 2005
- Keeling PJ, Burger G, Durnford DG, Lang BF, Lee RW, et al. The tree of eukaryotes. Trends Ecol Evol. 2005
- Simpson AGB, Roger AJ. The real ‘kingdoms’ of eukaryotes. Curr Biol. 2004
- Parfrey LW, Barbero E, Lasser E, Dunthorn M, Bhattacharya D, et al. Evaluating support for the current classification of eukaryotic diversity. PLoS Genet. 2006
- Burki F, Shalchian-Tabrizi K, Minge M, Skjaeveland A, Nikolaev SI, et al. Phylogenomics reshuffles the eukaryotic supergroups. PLoS ONE. 2007
- Bodyl A. Do plastid-related characters support the chromalveolate hypothesis? J Phycol. 2005
- Stiller JW, Riley J, Hall BD. Are red algae plants? A critical evaluation of three key molecular data sets. J Mol Evol. 2001
- Grzebyk D, Katz ME, Knoll AH, Quigg A, Raven JA, et al. Response to comment on «The evolution of modern eukaryotic phytoplankton». Science. 2004
- Yoon HS, Grant J, Tekle YI, Wu M, Chaon BC, et al. Broadly sampled multigene trees of eukaryotes. BMC Evol Biol. 2008
- Jarvis P, Soll M. Toc, Tic, and chloroplast protein import. Biochim Biophys Acta. 2001
- Marin B, Nowack ECM, Melkonian M. A plastid in the making: primary endosymbiosis. Protist. 2005
- Nowack ECM, Melkonian M, Glockner G. Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr Biol. 2008
- Theissen U, Martin W. The difference between organelles and endosymbionts. Curr Biol. 2006
- Bhattacharya D, Archibald JM. The difference between organelles and endosymbionts — response to Theissen and Martin. Curr Biol. 2006
- Okamoto N, Inouye I. The katablepharids are a distant sister group of the Cryptophyta: a proposal for Katablepharidophyta divisio nova/Kathablepharida phylum novum based on SSU rDNA and beta-tubulin phylogeny. Protist. 2005
- Andersen RA. Biology and systematics of heterokont and haptophyte algae. Am J Bot. 2004
- Cavalier-Smith T. Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins and the eukaryote family tree. J Eukaryot Microbiol. 1999
- Graham LE, Wilcox LW. Algae. Upper Saddle River, NJ: Prentice Hall; 2000
- Schnepf E, Elbrachter M. Dinophyte chloroplasts and phylogeny: a review. Grana. 1999
- Kohler S, Delwiche CF, Denny PW, Tilney LG, Webster P, et al. A plastid of probable green algal origin in apicomplexan parasites. Science. 1997
- Kohler S. Multi-membrane-bound structures of Apicomplexa: I. the architecture of the Toxoplasma gondii apicoplast. Parasitol Res. 2005
- Hopkins J, Fowler R, Krishna S, Wilson I, Mitchell G, et al. The plastid in Plasmodium falciparum asexual blood stages: a three-dimensional ultrastructural analysis. Protist. 1999
- Tomova C, Geerts WJC, Muller-Reichert T, Entzeroth R, Humbel BM. New comprehension of the apicoplast of Sarcocystis by transmission electron tomography. Biol Cell. 2006
- Moore RB, Obornik M, Janouskovec J, Chrudimsky T, Vancova M, et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature. 2008
- Stiller JW, Reel DC, Johnson JC. A single origin of plastids revisited: convergent evolution in organellar genome content. J Phycol. 2003
- Larkum AWD, Lockhart PJ, Howe CJ. Shopping for plastids. Trends Plant Sci. 2007
- McFadden GI, van Dooren GG. Evolution: red algal genome affirms a common origin of all plastids. Curr Biol. 2004
- Stiller JW, Hall BD. The origin of red algae: implications for plasmid evolution. Proc Natl Acad Sci U S A. 1997
- Sanchez-Puerta MV, Bachvaroff TR, Delwiche CF. Sorting wheat from chaff in multi-gene analyses of chlorophyll c-containing plastids. Mol Phylogenet Evol. 2007
- Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, et al. The evolution of modern eukaryotic phytoplankton. Science. 2004
- Fast NM, Kissinger JC, Roos DS, Keeling PJ. Nuclear-encoded, plastid-targeted genes suggest a single common origin for apicomplexan and dinoflagellate plastids. Mol Biol Evol. 2001
- Bucknam J, Boucher Y, Bapteste E. Refuting phylogenetic relationships. Biol Direct. 2006
- Gupta RS, Golding GB. Evolution of HSP70 gene and its implications regarding relationships between archaebacteria, eubacteria, and eukaryotes. J Mol Evol. 1993
- Gupta RS, Singh B. Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr Biol. 1994
- Gomez-Lorenzo MG, Spahn CMT, Agrawal RK, Grassucci RA, Penczek P, et al. Three-dimensional cryo-electron microscopy localization of EF2 in the Saccharomyces cerevisiae 80S ribosome at 17.5 angstrom resolution. EMBO J. 2000
- Jorgensen R, Merrill AR, Andersen GR. The life and death of translation elongation factor 2. Biochem Soc Trans. 2006
- Moreira D, Le Guyader H, Philippe H. The origin of red algae and the evolution of chloroplasts. Nature. 2000
- Germot a, Philippe H. Critical analysis of eukaryotic phylogeny: a case study based on the HSP70 family. J Eukaryot Microbiol. 1999
- Philippe H, Delsuc F, Brinkmann H, Lartillot N. Phylogenomics. Annu Rev Ecol Evol Syst. 2005
- Wiens JJ. Missing data and the design of phylogenetic analyses. J Biomed Inform. 2006
- Philippe H, Snell EA, Bapteste E, Lopez P, Holland PWH, et al. Phylogenomics of eukaryotes: Impact of missing data on large alignments. Mol Biol Evol. 2004
- Patron NJ, Inagaki Y, Keeling PJ. Multiple gene phylogenies support the monophyly of cryptomonad and haptophyte host lineages. Curr Biol. 2007
- Hackett JD, Yoon HS, Li S, Reyes-Prieto A, Rummele SE, et al. Phylogenomic analysis supports the monophyly of cryptophytes and haptophytes and the association of Rhizaria with Chromalveolates. Mol Biol Evol. 2007
- McFadden GI. Primary and secondary endosymbiosis and the origin of plastids. J Phycol. 2001
- Rodriguez-Ezpeleta N, Brinkmann H, Burey SC, Roure B, Burger G, et al. Monophyly of primary photosynthetic eukaryotes: green plants, red algae, and glaucophytes. Curr Biol. 2005
- Nosenko T, Bhattacharya D. Horizontal gene transfer in chromalveolates. BMC Evol Biol. 2007
- Lane CE, van den Heuvel K, Korera C, Curtis BA, Parsons BJ, et al. Nucleomorph genome of Hemiselmis andersenii reveals complete intron loss and compaction as a driver of protein structure and function. Proc Natl Acad Sci U S A. 2007
- Douglas S, Zauner S, Fraunholz M, Beaton M, Penny S, et al. The highly reduced genome of an enslaved algal nucleus. Nature. 2001
- Vørs N. Ultrastructure and autecology of the marine, heterotrophic flagellate Leucocryptos marina (Braaud) Butcher 1967 (Kathablepharidaceae/Kathablepharidae), with a discussion of the genera Leucocryptos and Katablepharis/Kathablepharis. Eur J Protistol. 1992
- McFadden GI, Gilson PR, Hill DRA. Goniomonas: ribosomal RNA sequences indicate that this phagotrophic flagellate is a close relative of the host component of cryptomonads. Eur J Phycol. 1994
- Maddison WP. Gene trees in species trees. Syst Biol. 1997
- Stiller JW. Plastid endosymbiosis, genome evolution and the origin of green plants. Trends Plant Sci. 2007
- Steiner JM, Yusa F, Pompe JA, Loffelhardt W. Homologous protein import machineries in chloroplasts and cyanelles. Plant J. 2005
- Stoebe B, Kowallik KV. Gene-cluster analysis in chloroplast genomics. Trends Genet. 1999
- Durnford DG, Deane JA, Tan S, McFadden GI, Gantt E, et al. A phylogenetic assessment of the eukaryotic light-harvesting antenna proteins, with implications for plastid evolution. J Mol Evol. 1999
- Rissler HM, Durnford DG. Isolation of a novel carotenoid-rich protein in Cyanophora paradoxa that is immunologically related to the light-harvesting complexes of photosynthetic eukaryotes. Plant Cell Physiol. 2005
- Stoebe B, Martin W, Kowallik KV. Distribution and nomenclature of protein-coding genes in 12 sequenced chloroplast genomes. Plant Mol Biol Rep. 1998
- Loffelhardt W, Bohnert HJ, Bryant DA. The complete sequence of the Cyanophora paradoxa cyanelle genome (Glaucocystophyceae). Plant Syst Evol. 1997
- O’Kelly C. Relationships of eukaryotic algal groups to other protists. In: Berner T, editor. Ultrastructure of microalgae. Boca Raton, FL: CRC Press; 1993
- Stiller JW, Harrell L. The largest subunit of RNA polymerase II from the Glaucocystophyta: functional constraint and short-branch exclusion in deep eukaryotic phylogeny. BMC Evol Biol. 2005
- Baldauf SL, Roger AJ, Wenk-Siefert I, Doolittle WF. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science. 2000
- Burger G, Saint-Louis D, Gray MW, Lang BF. Complete sequence of the mitochondrial DNA of the red alga Porphyra purpurea: cyanobacterial introns and shared ancestry of red and green algae. Plant Cell. 1999
- Secq MPO, Goer SL, Stam WT, Olsen JL. Complete mitochondrial genomes of the three brown algae (Heterokonta: Phaeophyceae) Dictyota dichotoma, Fucus vesiculosus and Desmarestia viridis. Curr Genet. 2006
- Kim E, Lane CE, Curtis BA, Kozera C, Bowman S, et al. Complete sequence and analysis of the mitochondrial genome of Hemiselmis andersenii CCMP644 (Cryptophyceae). BMC Genomics. 2008
- Gibbs SP. The Chloroplasts of some algal groups may have evolved from endosymbiotic eukaryotic algae. Ann N Y Acad Sci. 1981
- Rumpho ME, Summer EJ, Manhart JR. Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis. Plant Physiol. 2000
- Leander BS, Keeling PJ. Morphostasis in alveolate evolution. Trends Ecol Evol. 2003
- Moriya M, Nakayama T, Inouye I. A new class of the stramenopiles, Placididea classis nova: description of Placidia cafeteriopsis gen. et sp nov. Protist. 2002
- Kim E, Archibald JM. Diversity and evolution of plastids and their genomes. In: Sandelius AS, Aronsson H, editors. The Chloroplast: Interactions with the environment. Heidelberg: Springer; 2008
- Harper JT, Keeling PJ. Nucleus-encoded, plastid-targeted glyceraldehyde-3-phosphate dehydrogenase (GAPDH) indicates a single origin for chromalveolate plastids. Mol Biol Evol. 2003
- Takishita K, Ishida KI, Maruyama T. Phylogeny of nuclear-encoded plastid-targeted GAPDH gene supports separate origins for the peridinin- and the fucoxanthin derivative-containing plastids of dinoflagellates. Protist. 2004
- Takishita K, Kawachi M, Noel MH, Matsumoto T, Kakizoe N, et al. Origins of plastids and glyceraldehyde-3-phosphate dehydrogenase genes in the green-colored dinoflagellate Lepidodinium chlorophorum. Gene. 2008
- Martin W, Rujan T, Richly E, Hansen A, Cornelsen S, et al. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc Natl Acad Sci U S A. 2002
- Ohta N, Matsuzaki M, Misumi O, Miyagishima S, Nozaki H, et al. Complete sequence and analysis of the plastid genome of the unicellular red alga Cyanidioschyzon merolae. DNA Res. 2003
- Bachvaroff TR, Puerta MVS, Delwiche CF. Chlorophyll c-containing plastid relationships based on analyses of a multigene data set with all four chromalveolate lineages. Mol Biol Evol. 2005
- Bodyl A, Moszczynski K. Did the peridinin plastid evolve through tertiary endosymbiosis? A hypothesis. Eur J Phycol. 2006
- Lee RE, Kugrens P. Katablepharis ovalis, a colorless flagellate with interesting cytological characteristics. J Phycol. 1991
- Lee RE, Kugrens P, Mylnikov AP. The structure of the flagellar apparatus of two strains of Katablepharis (Cryptophyceae). Br Phycol J. 1992
- Clay B, Kugrens P. Systematics of the enigmatic kathablepharids, including EM characterization of the type species, Kathablepharis phoenikoston, and new observations on K. remigera com. nov. Protist. 1999
- Domozych DS, Wells B, Shaw PJ. Scale biogenesis in the green alga, Mesostigma viride. Protoplasma. 1992
- Domozych DS, Stewart KD, Mattox KR. Development of the cell wall in Tetraselmis: role of the Golgi apparatus and extracellular wall assembly. J Cell Sci. 1981
- Gupta RS. Protein phylogenies and signature sequences: a reappraisal of evolutionary relationships among archaebacteria, eubacteria, and eukaryotes. Microbiol Mol Biol Rev. 1998
- Boorstein WR, Ziegelhoffer T, Craig EA. Molecular evolution of the HSP70 multigene family. J Mol Evol. 1994
- Maddison DR, Maddison WP. MacClade 4: analysis of phylogeny and character evolution. Sunderland, MA: Sinauer Associates Inc; 2001
- Inagaki Y, Simpson AGB, Dacks JB, Roger AJ. Phylogenetic artifacts can be caused by leucine, serine, and arginine codon usage heterogeneity: dinoflagellate plastid origins as a case study. Syst Biol. 2004
- Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006
- Lartillot N, Brinkmann H, Philippe H. Suppression of long-branch attraction artefacts in the animal phylogeny using a site-heterogeneous model. BMC Evol Biol. 2007
- Abascal F, Zardoya R, Posada D. ProtTest: selection of best-fit models of protein evolution. Bioinformatics. 2005
- Schmidt HA, Strimmer K, Vingron M, von Haeseler A. TREE-PUZZLE: maximum likelihood phylogenetic analysis using quartets and parallel computing. Bioinformatics. 2002
- Desper R, Gascuel O. Fast and accurate phylogeny reconstruction algorithms based on the minimum-evolution principle. J Comput Biol. 2002
- Felsenstein J. Seattle: Department of Genome Sciences, University of Washington; 2005
Источник: ru.wikipedia.org
Особенности строения клеток
Клетки живых организмов классифицируют по разным признакам. Один из них — организация наследственного материала, заключенного в молекулах ДНК. Эукариоты — это организмы, в клетках которых находится оформленное ядро. Это двумембранная органелла, содержащая генетический материал. У прокариот данная структура отсутствует. К таким организмам относятся все виды бактерий и архей.
Строение прокариотических клеток
Отсутствие ядра не означает, что у прокариотических организмов нет наследственного материала. Он также закодирован в последовательности нуклеотидов. Однако располагается генетическая информация не в оформленном ядре, а представлена единственной кольцевой молекулой ДНК. Она называется плазмида. Такая молекула прикрепляется к внутренней поверхности плазматической мембраны. Клетки данного типа также лишены целого ряда определенных органелл. Прокариотические организмы характеризуются примитивностью, мелкими размерами и низким уровнем организации.
Что такое эукариот?
К этой многочисленной группе организмов относятся все представители растений, животных и грибов. Вирусы являются неклеточными формами жизни, поэтому в данной классификации не рассматриваются.
Поверхностный аппарат клеток прокариот представлен плазматической мембраной, а внутреннее содержимое — цитоплазмой. Это внутренняя полужидкая среда, которая выполняет опорную функцию, объединяет все структуры в единое целое. Для клеток прокариот также характерно наличие определенного ряда органоидов. Это комплекс Гольджи, эндоплпзматическая сеть, пластиды, лизосомы. Некоторые считают, что эукариоты — это организмы в клетках которых отсутствуют митохондрии. Но это совсем не так. Эти органеллы в клетках эукариот служат местом образования молекул АТФ-носителя энергии в клетке.
Эукариоты: примеры организмов
Эукариотами являются три царства живой природы. Однако несмотря на общие черты, у их клеток есть существенные отличия. К примеру, растительные характеризуются содержанием специализированных органелл хлоропластов. Именно в них происходит сложный фотохимический процесс превращения неорганических веществ в глюкозу и кислород. Животные клетки таких структур не имеют. Они способны усваивать только готовые питательные вещества. Отличаются эти структуры и строением поверхностного аппарата. В животных клетках над плазматической мембраной находится гликокаликс. Он представляет собой вязкий поверхностный слой, сосотоящий из белков, липидов и углеводов. Для растений характерна клеточная стенка. Она располагается над плазматической мембраной. Клеточная стенка образована сложными углеводами целлюлозой и пектином, которые придают ей прочность и жесткость.
Что такое эукариот, который представлен группой грибов? Клетки этих удивительных организмов сочетают в себе черты строения как растений, так и животных. В состав их клеточной стенки входят углеводы целлюлоза и хитин. Однако их цитоплпзма не содержит хлоропластов, поэтому они, подобно клеткам животных, способны только к гетеротрофному способу питания.
Прогрессивные черты строения эукариот
Почему все эукариоты являются организмами, которые достигли высокого уровня развития и распространения по планете? Прежде всего, благодаря высокому уровню специализации их органелл. Кольцевая молекуда ДНК, которая содержится в клетках бактерий, обеспечивает самый простой способ их размножения — деление клетки надвое. В результате данного процесса образуются точные генетические копии дочерних клеток. Размножение такого типа, безусловно, обеспечивает преемственность поколений и обеспечивает достаточно быстрое воспроизведение подобных клеток. Однако о появлении новых признаков в ходе деления надвое и речи идти не может. А это означает, что к изменяющимся условиям приспособиться эти организмы не смогут. Для клеток эукариот характерен половой процесс. В его ходе происходит обмен генетической информацией и ее рекомбинация. В результате на свет появляются особи с новыми, часто полезными признаками, которые закреплены в их генотипе и могут передаваться из поколения в поколение. Это и есть проявление наследственной изменчивости, которая является основой эволюции.
Итак, в нашей статье мы рассмотрели, что такое эукариот. Это понятие означает организм, клетки которого содержат ядро. К этой группе организмов относятся все представители растительного и животного мира, а также грибов. Ядро является постоянной клеточной структурой, которая обеспечивает хранение и передачу наследственной информиции организмов, закодированной в последовательности нуклеотидов молекул ДНК.
Источник: fb.ru
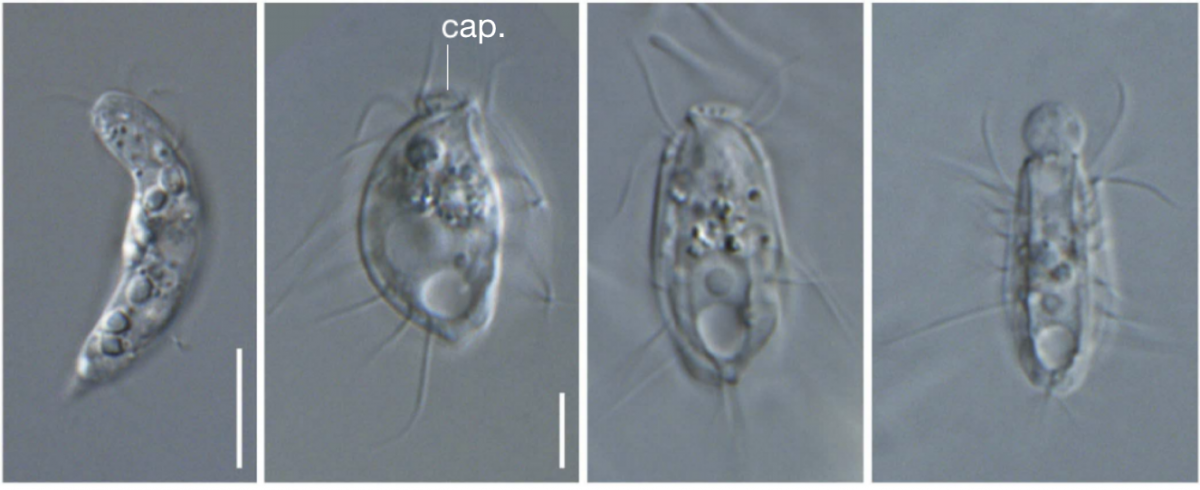
Во время туристического похода учащаяся Университета Дэлхаузи (Канада) Яна Эглит (Yana Eglit) из любопытства собрала образцы почвы за пределами города Галифакс. Позже оказалось, что в них содержатся микроорганизмы, которые не подходят ни к одному из существующих царств эукариотов. Исследование опубликовано в журнале Nature.
Сегодня существует несколько способов деления живых организмов на царства. Некоторые допускают до 20 вариантов, однако в основную классификацию входят пять царств: прокариоты (доядерные организмы), протисты, грибы, растения и животные. Впервые ее обосновал во второй половине XX столетия американский эколог Роберт Хардинг Уиттекер. Те микроорганизмы, которые попали в Университет Дэлхаузи, не соответствуют ни одному из этих царств, но они принадлежат группе микробов, которых называют хемимастиготами (hemimastigotes).
Хемимастиготы — одноклеточные микроорганизмы, о которых до недавнего времени практически ничего не знали. Их обнаружили еще в позапрошлом столетии, но не могли найти им место на дереве эволюции. В образцах почвы нашли два вида хемимастиготов. Первый — Spironema, который наблюдали и ранее, а второй — новый и до этого момента неизвестный. Его назвали Hemimastix kukwesjijk — в честь огра, фигурирующего в местных легендах. По словам нашедшей их студентки, они ведут себя необычно и заметить их удалось далеко не сразу.
Ученые воспользовались методами транскриптомики одиночных клеток, чтобы окончательно убедиться, что эти микроорганизмы не подходят ни к одному из существующих царств. По их словам, для поиска общего предка между этими хемимастиготами и любым другим организмом необходимо отправиться примерно на миллиард лет назад.
«Это открытие буквально перерисовывает наше древо жизни, добавляя новое ответвление на самых низких уровнях. Оно расширяет понимание эволюции сложных клеток и их происхождение, задолго до появления на Земле животных» — рассказал один из участников исследования Аластер Симпсон (Alastair Simpson).
Сейчас сотрудники лаборатории развели большое количество Hemimastix kukwesjijk, чтобы продолжить их детальное изучение. По их мнению, эти организмы могут привнести новую информацию в историю эволюции. Однако эта теория еще не стала общепризнанной, и в дальнейшем возможны дискуссии о месте хемимастиготов на древе жизни.
В прошлом месяце палеонтологи нашли место появления первых позвоночных. Это позволило заполнить многие бреши ранней эволюции позвоночных и показать, что первые шаги их развития проходили на морском мелководье — на глубине максимум несколько десятков метров.
Источник: naked-science.ru