К клеточным органоидам относятся
Определение понятия
Органоиды – это постоянные, обязательно присутствующие структуры клетки, выполняющие специфические функции и имеющие определенное строение.
Органоиды (синоним: органеллы) — это органы клетки, маленькие органы. По строению органоиды можно разделить на две группы: мембранные, в состав которых обязательно входят мембраны, и немембранные. В свою очередь, мембранные органоиды могут быть одномембранными – если образованы одной мембраной и двумембранными – если оболочка органоидов двойная и состоит из двух мембран.
Включения — это непостоянные структуры клетки, которые появляются в ней и исчезают в процессе метаболизма. Различают трофические, секреторные, экскреторные и пигментные включения.
Следует различать органоиды и включения.
Видео: Обзор клеточных структур
Видеообзор: https://youtu.be/URUJD5NEXC8
Органоиды (органеллы)
Мембрана (плазмолемма).
Ядро с ядрышком.
Эндоплазматическая сеть: шероховатая (гранулярная) и гладкая (агранулярная).
Видео: Синтез белков на гранулярном эндоплазматическом ретикулуме
Комплекс (аппарат) Гольджи
Видео: Комплекс Гольджи за работой
Митохондрии.
Рибосомы.
Лизосомы. Лизосомы (от гр. lysis — «разложение, растворение, распад» и soma — «тело») — это пузырьки диаметром 200—400 мкм. Имеют одномембранную оболочку, которая снаружи бывает покрыта волокнистым белковым слоем. Содержат набор ферментов (кислых гидролаз), которые при низких значениях рН в кислой среде проводят гидролитическое (в присутствии воды) расщепление веществ: нуклеиновых кислот, белков, жиров, углеводов. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Основная функция лизосом — внутриклеточное переваривание различных химических соединений и клеточных структур. Автолиз – саморазрушение клетки, наступающее вследствие высвобождения содержимого лизосом. В норме автолиз имеет место при метаморфозах (исчезновение хвоста у головастика лягушек), инволюции матки после родов, в очагах омертвления тканей.
Пероксисомы. Пероксисомы — это микротельца (пузырьки-везикулы) 0,1-1,5 мкм в диаметре, окружённые мембраной. Внутреннее содержимое пероксисомы (матрикс) представлен мелкогранулярным содержимым с нуклеоидом (сердцевиной) в центре. В нуклеоиде часто видны кристаллоподобные структуры, которые состоят из упорядоченно упакованных фибрилл и трубочек. Пероксисомы обычно локализуются вблизи мембран гранулярной эндоплазматической сети. Последние являются местом их образования, хотя часть ферментов пероксисом синтезируется в гиалоплазме. В пероксисомах обнаружено множество видов ферментов, связанных с метаболизмом перекиси водорода, окислением различных веществ, биосинтезом желчных кислот. Это ферменты, ведущие окислительное дезаминирование аминокислот (оксидазы, уратоксидазы) с образованием вредной для клетки перекиси водорода. В пероксисомах окисляются некоторые фенолы, Д-аминокислоты, а также жирные кислоты с очень длинными (более 22 углеродных атомов) цепями, которые не могут быть до укорачивания окислены в митохондриях.
кие жирные кислоты содержатся, например, в рапсовом масле. Для расщепления перекиси водорода Н2О2 на воду и кислород пероксисомы иеют фермент каталазу. Таким образом, эти органеллы разрушают органические соединения с образованием «клеточного яда» в виде перекиси водорода, но одновременно способны нейтрализовать её с помощью каталазы. Источники: http://meduniver.com/Medical/gistologia/24.html, http://mscience.ru/edu…
Функции пероксисом: окисление жирных кислот, фотодыхание, разрушение токсичных соединений, синтез желчных кислот, холестерина, а также эфиросодержащих липидов, построение миелиновой оболочки нервных волокон, метаболизме фитановой кислоты и т. д. Пероксисомы печени и почек обезвреживают множество ядовитых веществ, попадающих в кровоток. В частности, почти половина поступающего в организм человека этанола окисляется до ацетальдегида каталазой пероксисом. Наряду с митохондриями пероксисомы являются главными потребителями O2 в клетке.
Протеасомы
Протеасомы – специальные органоиды для разрушения белков. Название «протеасома» – (protos — главный, первичный и soma — тело) показывает, что это органоид, способный к протеолизу – лизису белков. В клетке имеется протеасомы двух видов, различающиеся молекулярной массой: с коэффициентом седиментации (осаждения) 20S и 26S (S – единица Сведберга).
S – протеасома имеет форму полого цилиндра 15-17 нм и диаметром 11-12 нм. Она состоит из 4 лежащих друг на друге колец двух типов. Каждое кольцо содержит 7 белковых субъединиц и включает 12-15 полипептидов. На внутренней стороне цилиндра находятся 3 протеолитические камеры. Протеолиз происходит в центральной камере. В этой камере расщепляются токсичные или ставшие неполноценными и ненужными клетке белки.
Маркировкой ненужных белков занимается специфическая система ферментов – система убиквитирования. Система присоединяет белок убиквитин (ubique — вездесущий) к молекуле белка, который должен быть уничтожен. Сигналами для убиквитирования и последующей деградации могут служить нарушения в структуре белковых молекул. Имеются данные о связи некоторых наследственных заболеваний человека (фиброкистоз, синдром Ангельмана) с нарушениями в ферментах реакции убиквитирования. Предполагается, что нарушения в работе протеасомной системы деградации белка являются причиной некоторых нейродегенеративных болезней. Источники: http://mscience.ru/edu_articles/biology_articles/161-peroksisomy-i-prote…
Видео: Протеасомы.
Фагосомы
Микрофиламенты. Каждый микрофиламент — это двойная спираль из глобулярных молекул белка актина.
этому содержание актина даже в немышечных клетках достигает 10 % от всех белков.
В узлах сети микрофиламентов и в местах их прикрепления к клеточным структурам находятся белок a-актинин, а также, белки миозин и тропомиозин.
Микрофиламенты образуют в клетках более или менее густую сеть. Так, например, в микрофаге насчитывается около 100.000 микрофиламентов. Функции микрофиламентов:
— миграция клеток в эмбриогенезе,
— передвижение макрофагов,
— фаго- и пиноцитоз,
— рост аксонов (у нейронов),
— образование каркаса для микроворсинок и обеспечение всасывания в кишечнике и реабсорбции в почечных канальцах.
Промежуточные филаменты. Являются компонентом цитоскелета. Они толще микрофиламентов и имеют тканеспецифическую природу:
— в эпителии они образованы белком кератином,
— в клетках соединительной ткани — виментином,
— в гладких мышечных клетках — десмином,
— в нервных клетках они называются нейрофиламентами и тоже образованы особым белком.
Промежуточные филаменты часто располагаются параллельно поверхности клеточного ядра.
Микротрубочки. Микротрубочки образуют в клетке густую сеть. Она начинается от перинуклеарной области (от центриоли) и радиально распространяется к плазмолемме, следуя за изменениями её формы. Также микротрубочки идут вдоль длинной оси отростков клеток.
клетках с ресничками микротрубочки образуют аксонему (осевую нить) ресничек.
Стенка микротрубочки состоит из одного слоя глобулярных субъединиц белка тубулина.
На поперечном срезе видно 13 таких субъединиц, образующих кольцо.
Его параметры таковы:
— внешний диаметр — dex = 24 нм,
— внутренний диаметр — din = 14 нм,
— толщина стенки — l стенки = 5 нм.
Как и микрофиламенты, микротрубочки образуются путём самосборки. Это происходит при сдвиге равновесия между свободной и связанной формами тубулина в сторону связанной формы.
В неделящейся интерфазной клетке создаваемая микротрубочками сеть играет роль цитоскелета, поддерживающего форму клетки.
Транспорт веществ по длинным отросткам нервных клеток идёт не внутри микротрубочек, а вдоль них по перитубулярному пространству. Но микротрубочки выступают при этом в роли направительных структур: Белки-транслокаторы (динеины и кинезины), перемещаясь по внешней поверхности микротрубочек, «тащат» за собой и мелкие пузырьки с транспортируемыми веществами.
В делящихся клетках сеть микротрубочек перестраивается и формирует веретено деления. Они связывают хроматиды хромосом с центриолями и способствуют правильному расхождению хроматид к полюсам делящейся клетки.
Клеточный центр.
Пластиды.
Вакуоли. Вакуоли – одномембранные органоиды. Они представляют собой мембранные «ёмкости», пузыри, заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Вакуоли характерны для растительных клеток. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы). Из органических веществ чаще запасаются сахара и белки. Сахара – чаще в виде растворов, белки поступают в виде пузырьков ЭПР и аппарата Гольджи, после чего вакуоли обезвоживаются, превращаясь в алейроновые зерна. В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции вакуолей.
стительные вакуоли отвечают за накопление воды и поддержание тургорного давления, накопление водорастворимых метаболитов – запасных питательных веществ и минеральных солей, окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян. Пищеварительные и автофагические вакуоли – разрушают органические макромолекулы; сократительные вакуоли регулируют осмотическое давление клетки и выводят ненужные вещества из клетки.
Эндоплазматическая сеть, аппарат Гольджи, лизосомы, пероксисомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Включения
Включения. Включения — это непостоянные структуры клетки, которые появляются в ней и исчезают в процессе метаболизма. Различают трофические, секреторные, экскреторные и пигментные включения.
Группа трофических включений объединяет углеводные, липидные и белковые включения. Наиболее распространенным представителем углеводных включений является гликоген — полимер глюкозы. На светооптическом уровне наблюдать включения гликогена можно при использовании гистохимической ШИК-реакции. В электронном микроскопе гликоген выявляется как осмиофильные гранулы, которые в клетках, где гликогена много (гепатоцитах), сливаются в крупные конгломераты — глыбки.
Липидными включениями наиболее богаты клетки жировой ткани — липоциты, резервирующие запасы жира для нужд всего организма, а также стероидпродуцирующие эндокринные клетки, использующие липид холестерин для синтеза своих гормонов.
ультрамикроскопическом уровне липидные включения имеют правильную округлую форму и в зависимости от химического состава характеризуются высокой, средней или низкой электронной плотностью.
Белковые включения, например, вителлин в яйцеклетках, накапливается в цитоплазме в виде гранул. Секреторные включения представляют собой разнообразную группу.
Секреторные включения синтезируются в клетках и выделяются (секретируются) в просветы протоков (клетки экзокринных желез), в межклеточную среду (гормоны, нейромедиаторы, факторы роста и др.), кровь, лимфу, межклеточные пространства (гормоны). На ультрамикроскопическом уровне секреторные включения имеют вид мембранных пузырьков, содержащих вещества разной плотности и интенсивности окраски, что зависит от их химического состава.
Экскреторные включения — это, как правило, продукты метаболизма клетки, от которых она должна освободиться. К экскреторным включениям относятся также инородные включения — случайно, либо преднамеренно (при фагоцитозе бактерий, например,) попавшие в клетку субстраты. Такие включения клетка лизирует с помощью своей лизосомальной системы, а оставшиеся частицы выводит (экскретирует ) во внешнюю среду. В более редких случаях попавшие в клетку агенты остаются неизменными и могут не подвергнуться экскреции — такие включения более правильно именовать чужеродными (хотя чужеродными для клетки являются и включения, которые она лизирует).
Пигментные включения хорошо выявляются как на светооптическом, так и на ультрамикроскопическом уровнях.
ень характерный вид они имеют на электронных микрофотографиях — в виде осмиофильных структур разных размеров и формы. Данная группа включений характерна для пигментоцитов. Пигментоциты, присутствуя в дерме кожи, защищают организм от глубокого проникновения опасного для него ультрафиолетового излучения, в радужке, сосудистой оболочке и сетчатке глаза пигментоциты регулируют поток света на фоторецепторные элементы глаза и предохраняют их от перераздражения светом. В процессе старения очень многие соматические клетки накапливают пигмент липофусцин, по присутствию которого можно судить о возрасте клетки. В эритроцитах и симпластах скелетных мышечных волокон присутствуют соответственно гемоглобин или миоглобин — пигменты-переносчики кислорода и углекислоты.
Источник: http://meduniver.com/Medical/gistologia/24.html MedUniver
Видео: .Внутренняя жизнь клетки. Упаковка белков
Источник: kineziolog.su
2.5.2.1. Мембранные органоиды (органеллы)
Эндоплазматическая сеть (эндоплазматический ретикулум, цитоплазматическая сеть) — совокупность сообщающихся между собой канальцев, вакуолей и «цистерн», стенка которых образована элементарными биологическими мембранами. Открыта К.Р. Портером в 1945 году. Открытие и описание эндоплазматической сети (ЭПС) обязано внедрению в практику цитологических исследований электронного микроскопа. Мембраны, образующие ЭПС, отличаются от плазмолеммы клетки меньшей толщиной (5-7 нм) и большей концентрацией белков, в первую очередь обладающих ферментативной активностью. Различают две разновидности ЭПС (рис. 8): шероховатую (гранулярную) и гладкую (агранулярную). Шероховатая ЭПС представлена уплощенными цистернами, на поверхности которых расположены рибосомы и полисомы. Мембраны гранулярной ЭПС содержат белки, способствующие связыванию рибосом и уплощению цистерн. Особенно хорошо развита шероховатая ЭПС в клетках, специализирующихся на белковом синтезе. Гладкую ЭПС формируют переплетающиеся канальцы, трубочки и небольшие пузырьки. Каналы и цистерны ЭПС этих двух разновидностей не разграничены: мембраны одного типа переходят в мембраны другого типа, формируя в области перехода так называемую переходную (транзиторную) ЭПС.
Основными функциями гранулярной ЭПС являются:
1) синтез на прикреплённых рибосомах белков (секретируемых белков, белков клеточных мембран и специфических белков содержимого мембранных органоидов); 2) гидроксилирование, сульфатирование, фосфорилирование и гликозилирование белков; 3) транспорт веществ в пределах цитоплазмы; 4) накопление как синтезируемых, так и транспортируемых веществ; 5) регуляция биохимических реакций, связанная с упорядоченностью локализации в структурах ЭПС веществ, вступающих в реакции, а также их катализаторов — ферментов.
Гладкая ЭПС отличается отсутствием на мембранах белков (рибофоринов), связывающих субъединицы рибосом. Предполагается, что гладкая ЭПС образуется в результате формирования выростов шероховатой ЭПС, мембрана которых утрачивает рибосомы.
Функциями гладкой ЭПС являются: 1) синтез липидов, включая мембранные липиды; 2) синтез углеводов (гликогена и др.); 3) синтез холестерина; 4) обезвреживание токсических веществ эндогенного и экзогенного происхождения; 5) накопление ионов Са2+; 6) восстановление кариолеммы в телофазе митоза; 7) транспорт веществ; 8) накопление веществ.
Как правило, гладкая ЭПС развита в клетках слабее, чем шероховатая ЭПС, однако в клетках, вырабатывающих стероиды, триглицериды и холестерин, а также в клетках печени, осуществляющих детоксикацию различных веществ, она развита значительно лучше.
Рис. 9. Комплекс Гольджи:
1 – стопка уплощённых цистерн; 2 – пузырьки; 3 – секреторные пузырьки (вакуоли)
Переходная (транзиторная) ЭПС — это участок перехода гранулярной ЭПС в агранулярную ЭПС, который располагается у формирующейся поверхности комплекса Гольджи. Трубочки и канальцы переходной ЭПС распадаются на фрагменты, из которых образуются пузырьки, транспортирующие материал из ЭПС в комплекс Гольджи.
Пластинчатый комплекс (комплекс Гольджи, аппарат Гольджи) — органоид клетки, участвующий в окончательном формировании продуктов её жизнедеятельности (секретов, коллагена, гликогена, липидов и других продуктов), а также в синтезе гликопротеидов. Органоид назван по имени описавшего его в 1898 году итальянского гистолога К. Гольджи. Образован тремя составляющими (рис. 9): 1) стопкой уплощённых цистерн (мешочков); 2) пузырьками; 3) секреторными пузырьками (вакуолями). Зона скопления этих элементов получила название диктиосомы. Таких зон в клетке может быть несколько (иногда несколько десятков и даже сотен). Комплекс Гольджи располагается около ядра клетки, часто вблизи центриолей, реже рассеян по всей цитоплазме. В секреторных клетках он располагается в апикальной части клетки, через которую осуществляется выделение секрета путём экзоцитоза. От 3-х до 30-ти цистерн в виде изогнутых дисков диаметром 0,5-5 мкм образуют стопку. Смежные цистерны разделены пространствами в 15-30 нм. Отдельные группы цистерн в пределах диктиосомы отличаются особым составом ферментов, определяющих характер биохимических реакций, в частности процессинга белка и др.
Второй составляющий элемент диктиосомы — пузырьки представляют собой сферические образования диаметром 40-80 нм, умеренно плотное содержимое которых окружено мембраной. Пузырьки формируются путём отщепления от цистерн.
Третий элемент диктиосомы — секреторные пузырьки (вакуоли) представляют собой относительно крупные (0,1-1,0 мкм) сферические мембранные образования, содержащие секрет умеренной плотности, претерпевающий конденсацию и уплотнение (вакуоли конденсации).
Комплекс Гольджи отчётливо поляризован по вертикали. В нём выделяют две поверхности (два полюса):
1) цис-поверхность, или незрелую поверхность, которая имеет выпуклую форму, обращена к эндоплазматической сети (ядру) и связана с отделяющимися от неё мелкими транспортными пузырьками;
2) транс-поверхность, или поверхность, обращённую к плазмолемме вогнутой формы (рис. 8), со стороны которой от цистерн комплекса Гольджи отделяются вакуоли (секреторные гранулы).
Основными функциями комплекса Гольджи являются: 1) синтез гликопротеинов и полисахаридов; 2) модификация первичного секрета, его конденсация и упаковка в мембранные пузырьки (формирование секреторных гранул); 3) процессинг молекул (фосфорилирование, сульфатирование, ацилирование и т.п.); 4) накопление секретируемых клеткой веществ; 5) образование лизосом; 6) сортировка синтезированных клеткой белков у транс-поверхности перед их окончательным транспортом (производится посредством рецепторных белков, распознающих сигнальные участки макромолекул и направляющих их в различные пузырьки); 7) транспорт веществ: из транспортных пузырьков вещества проникают в стопку цистерн комплекса Гольджи с цис-поверхности, а выходят из неё в виде вакуолей с транс-поверхности. Механизм транспорта объясняют две модели: а) модель перемещения пузырьков, отпочковывающихся от предшествующей цистерны и сливающихся с последующей цистерной последовательно в направлении от цис-поверхности к транс-поверхности; б) модель перемещения цистерн, основанная на представлении о непрерывном новообразовании цистерн за счёт слияния пузырьков на цис-поверхности и последующем распаде на вакуоли цистерн, смещающихся к транс-поверхности.
Указанные выше основные функции позволяют констатировать, что пластинчатый комплекс — важнейший органоид клетки эукариот, обеспечивающий организацию и интеграцию внутриклеточного метаболизма. В этом органоиде протекают заключительные этапы формирования, созревания, сортировки и упаковки всех секретируемых клеткой продуктов, ферментов лизосом, а также белков и гликопротеинов поверхностного аппарата клетки и др. веществ.
Органоиды внутриклеточного переваривания. Лизосомы — это мелкие ограниченные элементарной мембраной пузырьки, содержащие гидролитические ферменты. Мембрана лизосом толщиной около 6 нм осуществляет пассивную компартментализацию, временно отделяя гидролитические ферменты (более 30 разновидностей) от гиалоплазмы. В неповреждённом состоянии мембрана устойчива к действию гидролитических ферментов и препятствует их утечке в гиалоплазму. В стабилизации мембраны важная роль принадлежит кортикостероидным гормонам. Повреждение мембран лизосом ведёт к самоперевариванию клетки гидролитическими ферментами.
Мембрана лизосом содержит АТФ-зависимый протонный насос, обеспечивающий закисление среды внутри лизосом. Последняя способствует активизации ферментов лизосом — кислых гидролаз. Наряду с этим мембрана лизосом содержит рецепторы, обусловливающие связывание лизосом с транспортными пузырьками и фагосомами. Мембрана обеспечивает также диффузию веществ из лизосом в гиалоплазму. Связывание части молекул гидролаз с мембраной лизосом ведёт к их инактивации.
Выделяют несколько разновидностей лизосом: первичные лизосомы (гидролазные пузырьки), вторичные лизосомы (фаголизосомы, или пищеварительные вакуоли), эндосомы, фагосомы, аутофаголизосомы, остаточные тельца (рис. 8).
Эндосомами называют мембранные пузырьки, переносящие макромолекулы от поверхности клетки в лизосомы путём эндоцитоза. В процессе переноса содержимое эндосом может не изменяться или претерпевать частичное расщепление. В последнем случае в эндосомы проникают гидролазы или эндосомы непосредственно сливаются с гидролазными пузырьками, вследствие чего среда постепенно закисляется. Эндосомы разделяют на две группы: ранние (периферические) и поздние (перинуклеарные) эндосомы.
Ранние (периферические) эндосомы формируются на ранних этапах эндоцитоза после отделения пузырьков с захваченным содержимым от плазмолеммы. Они располагаются в периферических слоях цитоплазмы и характеризуются нейтральной или слабощелочной средой. В них происходит отщепление лигандов от рецепторов, сортировка лигандов и, возможно, возвращение рецепторов в специальных пузырьках в плазмолемму. Наряду с этим в ранних эндосомах может происходить расщепление ком-
Рис. 10 (А). Схема образования лизосом и их участия во внутриклеточном пищеварении. (Б) Электронная микрофотография среза вторичных лизосом (обозначены стрелками):
1 – образование из гранулярной эндоплазматической сети мелких пузырьков с ферментами; 2 – перенос ферментов в аппарат Гольджи; 3 – образование первичных лизосом; 4 – выделение и использование (5) гидролаз при внеклеточном ращеплении; 6 — фагосомы; 7 – слияние первичных лизосом с фагосомами; 8, 9 – образование вторичных лизосом (фаголизосом); 10 – экскреция остаточных телец; 11 – слияние первичных лизосом с разрушающимися структурами клетки; 12 – аутофаголизосома
плексов «рецептор-гормон», «антиген-антитело», ограниченное расщепление антигенов, инактивация отдельных молекул. В условиях закисления (рН=6,0) среды в ранних эндосомах может происходить частичное расщепление макромолекул. Постепенно, перемещаясь вглубь цитоплазмы, ранние эндосомы превращаются в поздние (перинуклеарные) эндосомы, располагающиеся в глубоких слоях цитоплазмы, окружающих ядро. Они достигают 0,6-0,8 мкм в диаметре и отличаются от ранних эндосом более кислым (рН=5,5) содержимым и более высоким уровнем ферментативного переваривания содержимого.
Фагосомы (гетерофагосомы) — мембранные пузырьки, которые содержат захваченный клеткой извне материал, подлежащий внутриклеточному перевариванию.
Первичные лизосомы (гидролазные пузырьки) — пузырьки диаметром 0,2-0,5 мкм, содержащие неактивные ферменты (рис.10). Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки осуществляют транспорт гидролитических ферментов из пластинчатого комплекса к органоидам эндоцитозного пути (фагосомам, эндосомам и т.п.).
Вторичные лизосомы (фаголизосомы, пищеварительные вакуоли) — пузырьки, в которых активно осуществляется внутриклеточное переваривание посредством гидролаз при рН≤5. Их диаметр достигает 0,5-2 мкм. Вторичные лизосомы (фаголизосомы и аутофаголизосомы) формируются путём слияния фагосомы с эндосомой или первичной лизосомой (фаголизосомы) либо путём слияния аутофагосомы (мембранного пузырька, содержащего собственные компоненты клетки) с первичной лизосомой (рис. 10) или поздней эндосомой (аутофаголизосомы). Аутофагия обеспечивает переваривание участков цитоплазмы, митохондрий, рибосом, фрагментов мембран и т.п. Убыль последних в клетке компенсируется их новообразованием, что ведёт к обновлению («омоложению») клеточных структур. Так, в нервных клетках человека, функционирующих многие десятилетия, большинство органоидов обновляется в течение 1 месяца.
Разновидность лизосом, содержащих непереваренные вещества (структуры), названа остаточными тельцами. Последние могут длительно находиться в цитоплазме или выделять своё содержимое путём экзоцитоза за пределы клетки (рис. 10). Распространённым видом остаточных телец в организме животных являются липофусциновые гранулы, представляющие собой мембранные пузырьки (0,3-3 мкм), содержащие труднорастворимый коричневый пигмент липофусцин.
Пероксисомы представляют собой мембранные пузырьки диаметром до 1,5 мкм, матрикс которых содержит около 15 ферментов (рис. 8). Среди последних наиболее важны каталаза, на которую приходится до 40% общего белка органоида, а также пероксидаза, оксидаза аминокислот и др. Пероксисомы образуются в эндоплазматическом ретикулуме и обновляются каждые 5-6 дней. Наряду с митохондриями, пероксисомы являются важным центром утилизации кислорода в клетке. В частности, под воздействием каталазы распадается перекись водорода (Н2О2), образующаяся в ходе окисления аминокислот, углеводов и др. веществ клетки. Таким образом, пероксисомы защищают клетку от повреждающего эффекта перекиси водорода.
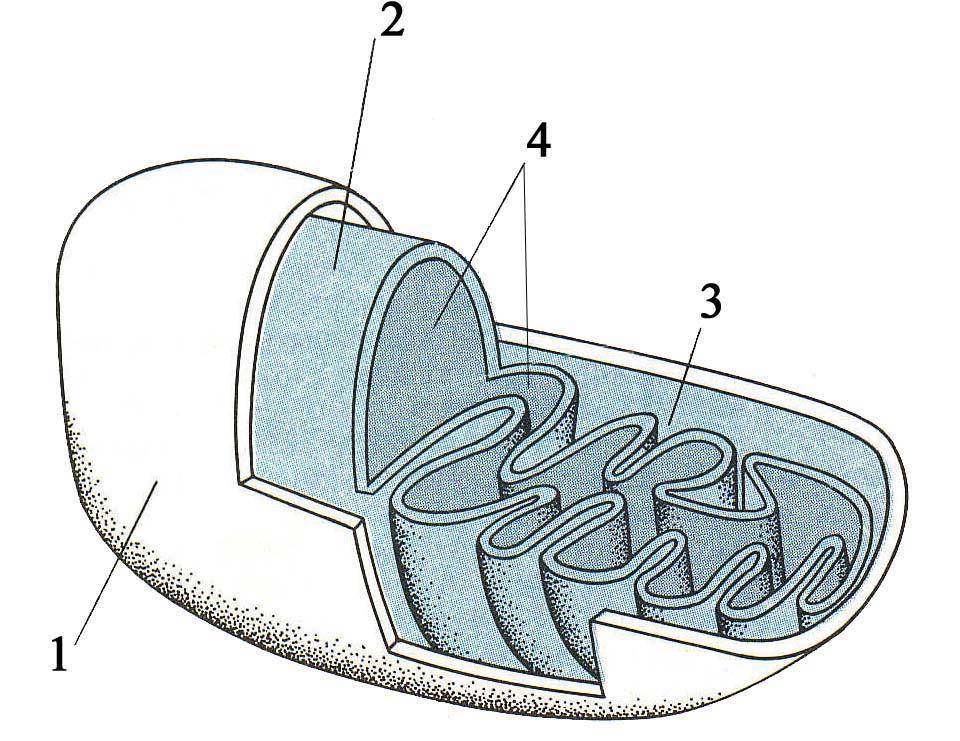
Органоиды энергетического обмена. Митохондрии описаны впервые Р. Келликером в 1850 году в мышцах насекомых под названием саркосом. Позднее они изучались и описывались Р. Альтманом в 1894 году как «биопласты», а в 1897 году К. Бенда назвал их митохондриями. Митохондрии представляют собой мембранные органоиды, обеспечивающие клетку (организм) энергией. Источником запасаемой в виде фосфатных связей АТФ энергии являются процессы окисления. Наряду с этим митохондрии участвуют в биосинтезе стероидов и нуклеиновых кислот, а также в окислении жирных кислот.
М
Рис. 11. Схема строения митохондрии:
1 – наружная мембрана; 2 – внутренняя мембрана; 3 – кристы; 4 – матрикс
итохондрии имеют эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Их размеры составляют 0,2-2 мкм в ширину и 2-10 мкм в длину. Количество митохондрий в различных клетках варьирует в широких пределах, достигая в наиболее активных 500-1000. В клетках печени (гепатоцитах) их число составляет около 800, а занимаемый ими объем равен примерно 20% объема цитоплазмы. В цитоплазме митохондрии могут располагаться диффузно, однакообычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонема спермия). Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны — кристы (рис. 11, 12).
Н
Рис. 12. Электронная фотография митохондрии (поперечный разрез)
аружная мембрана митохондрий сходна с плазмолеммой. Она отличается высокой проницаемостью, обеспечивая проникновение молекул с массой менее 10 килодальтон из цитозоля в межмембранное пространство митохондрий. Наружная мембрана содержит порин и другие транспортные белки, а также рецепторы, распознающие переносимые белки в зонах слипания наружной и внутренней мембран.
Межмембранное пространство митохондрий шириной 10-20 нм содержит небольшое количество ферментов. Его ограничивает изнутри внутренняя мембрана митохондрий, содержащая транспортные белки, ферменты дыхательной цепи и сукцинатдегидрогеназу, а также комплекс АТФ-синтетазы. Внутренняя мембрана характеризуется низкой проницаемостью для мелких ионов. Она формирует складки толщиной 20 нм, которые располагаются чаще всего перпендикулярно продольной оси митохондрий, а в некоторых случаях (мышечные и др. клетки) — продольно. С повышением активности митохондрий количество складок (их общая площадь) возрастает. На кристах находятся оксисомы — грибовидные образования, состоящие из округлой головки диаметром 9 нм и ножки толщиной 3 нм. В области головки происходит синтез АТФ. Процессы окисления и синтеза АТФ в митохондриях разобщены, из-за чего не вся энергия накапливается в АТФ, рассеиваясь частично в виде тепла. Такое разобщение наиболее выражено, например, в бурой жировой ткани, используемой для весеннего «разогрева» находившихся в состоянии «зимней спячки» животных.
Внутренняя камера митохондрии (область между внутренней мембраной и кристами) заполнена матриксом (рис. 11, 12), содержащим ферменты цикла Кребса, ферменты белкового синтеза, ферменты окисления жирных кислот, митохондриальную ДНК, рибосомы и митохондриальные гранулы.
Митохондриальная ДНК представляет собственный генетический аппарат митохондрий. Она имеет вид кольцевой двухцепочечной молекулы, в которой содержится около 37 генов. Митохондриальная ДНК отличается от ядерной ДНК низким содержанием некодирующих последовательностей и отсутствием связей с гистонами. Митохондриальная ДНК кодирует иРНК, тРНК и рРНК, однако обеспечивает синтез только 5-6% митохондриальных белков (ферментов системы транспорта ионов и некоторых ферментов синтеза АТФ). Синтез всех других белков, а также удвоение митохондрий контролируются ядерной ДНК. Большая часть рибосомальных белков митохондрий синтезируется в цитоплазме, а затем транспортируется в митохондрии. Наследование митохондриальной ДНК у многих видов эукариот, включая человека, происходит только по материнской линии: митохондриальная ДНК отца исчезает при гаметогенезе и оплодотворении.
Митохондрии имеют относительно короткий жизненный цикл (около 10 суток). Разрушение их происходит путём аутофагии, а новообразование — путём деления (перешнуровки) предшествующих митохондрий. Последнему предшествует репликация митохондриальной ДНК, которая происходит независимо от репликации ядерной ДНК в любые фазы клеточного цикла.
У прокариот митохондрии отсутствуют, и их функции выполняет клеточная мембрана. Согласно одной из гипотез, митохондрии произошли из аэробных бактерий в результате симбиогенеза. Существует предположение об участии митохондрий в передаче наследственной информации.
Источник: StudFiles.net
Что такое органоиды клетки
Органоиды клетки, они же органеллы, представляют собой специализированные структуры собственно клетки, отвечающие за различные важные и жизненно необходимые функции. Почему же все-таки «органоиды»? Просто тут эти компоненты клетки сопоставляются с органами многоклеточного организма.
Какие органоиды входят в состав клетки
Также порой под органоидами понимается исключительно лишь постоянные структуры клетки, которые находятся в ее цитоплазме. По этой же причине ядро клетки и ее ядрышко не называют органоидами, равно как и не являются органоидами клеточная мембрана, реснички и жгутики. А вот к органоидам, входящим в состав клетки относятся: хромосомы, митохондрии, комплекс Гольджи, эндоплазматическая сеть, рибосомы, микротрубочки, микрофиламенты, лизосомы. По сути это и есть основные органоиды клетки.
Если речь идет о животных клетках, то в число их органоидов также входят центриоли и микрофибриллы. А вот в число органоидов растительной клетки еще входят только свойственные растениям пластиды. В целом состав органоидов в клетках может существенно отличатся в зависимости от вида самой клетки.
Рисунок строения клетки, включая ее органоиды.
Двумембраные органоиды клетки
Также в биологии существует такое явление как двумембраные органоиды клетки, к ним относятся митохондрии и пластиды. Ниже мы опишем свойственные им функции, впрочем, как всех других основных органоидов.
Функции органоидов клетки
А теперь коротко опишем основные функции органоидов животной клетки. Итак:
- Плазматическая мембрана – тонкая пленка вокруг клетки состоящая из липидов и белков. Очень важный органоид, который обеспечивает транспортировку в клетку воды, минеральных и органических веществ, удаляет вредные продукты жизнедеятельности и защищает клетку.
- Цитоплазма – внутренняя полужидкая среда клетки. Обеспечивает связь между ядром и органоидами.
- Эндоплазматическая сеть – она же сеть каналов в цитоплазме. Принимает активное участие в синтезе белков, углеводов и липидов, занимается транспортировкой полезных веществ.
- Митохондрии – органоиды, в которых окисляются органические вещества и синтезируются молекулы АТФ с участием ферментов. По сути митохондрии это органоид клетки, синтезирующий энергию.
- Пластиды (хлоропласты, лейкопласты, хромопласты) – как мы упоминали выше, встречаются исключительно у растительных клеток, в целом их наличие является главной особенностью растительного организма. Играют очень важную функцию, например, хлоропласты, содержащие зеленый пигмент хлорофилл, у растения отвечают за явление фотосинтеза.
- Комплекс Гольджи — система полостей, отграниченных от цитоплазмы мембраной. Осуществляют синтез жиров и углеводов на мембране.
- Лизосомы — тельца, отделенные от цитоплазмы мембраной. Имеющиеся в них особые ферменты ускоряют реакцию расщепления сложных молекул. Также лизосома является органоидом, обеспечивающим сборку белка в клетках.
- Вакуоли — полости в цитоплазме, заполненные клеточным соком, место накопления запасных питательных веществ; они регулируют содержание воды в клетке.
В целом все органоиды являются важными, ведь они регулируют жизнедеятельность клетки.
Источник: www.poznavayka.org
1. Ведущую роль в эволюции играет:
а – мутационная изменчивость;
б – модификационная изменчивость;
в – групповая изменчивость;
г – ненаследственная изменчивость.
2. Основным критерием вида является:
а – физиологический;
б – географический;
в – экологический;
г – все эти критерии
3. Более чем одно ядро может встречаться в клетках:
а – простейших;
б – мышц;
в – соединительной ткани;
г – все ответы верны.
4. Последовательное уменьшение числа пальцев у предков лошади служит примером:
а – гомологического ряда;
б – филогенетического ряда;
в – ароморфоза;
г – конвергенции.
5. Микроэволюция приводит к образованию новых:
а – семейных групп;
б – подвидов и видов;
в – родов;
г – классов.
6. Закон Моргана касается:
а – дигибридного скрещивания;
б – чистоты гамет;
в – неполного доминирования;
г – сцепления генов.
7. Основное количество солнечной энергии в океане запасает:
а – фитопланктон;
б – зоопланктон;
в – рыба и морские млекопитающие;
г – крупные придонные водоросли.
8. Число нуклеотидов, вмещающихся в рибосому, равно:
а – один;
б – три;
в – шесть;
г – девять.
9. К обезьянолюдям относятся:
а – кроманьонец;
б – австралопитек;
в – питекантроп;
г – неандерталец.
10. При дигибридном скрещивании число классов по фенотипу во втором поколении равно:
а – четыре;
б – девять;
в – шестнадцать;
г – ни один ответ не верен.
Ответы:
1) а.
2) г.
3) б.
4) б.
5) б.
6) г.
7)
8)
9)
10) в.
Задание 2. Задание содержит вопросы, к каждому из которых даны несколько вариантов ответа; среди них правильных может быть от нуля до пяти.
1. Какие клеточные органоиды имеют ДНК:
а – центриоль;
б — вакуоль;
в- митохондрии;
г — ядро;
д — лизосомы.
2. Какие из перечисленных клеточных структур имеют двойную мембрану:
а – вакуоль;
б – митохондрии;
в – хлоропласты;
г – мембрана прокариот;
д – мембрана эукариот;
е – ядро;
3. К гетеротрофам относятся:
а – фитопланктон;
б – грибы;
в – птицы;
г – бактерии;
д — хвойные.
4. Для вирусов характерны следующие признаки:
а – не могут размножаться вне клеток тех организмов, в которых паразитируют;
б – имеют хитиновую клеточную оболочку;
в – содержат лишь один тип нуклеиновых кислот – либо РНК, либо ДНК;
г – могут являться причиной раковых опухолей;
д – представлены только одноцепочечными молекулами нуклеиновых кислот.
5. Единицей эволюционного процесса является:
а – вид;
б – совокупность особей;
в – популяция;
Задание 3.
Источник: biologia.neznaka.ru
2.5.2.1. Мембранные органоиды (органеллы)
Эндоплазматическая сеть (эндоплазматический ретикулум, цитоплазматическая сеть) — совокупность сообщающихся между собой канальцев, вакуолей и «цистерн», стенка которых образована элементарными биологическими мембранами. Открыта К.Р. Портером в 1945 году. Открытие и описание эндоплазматической сети (ЭПС) обязано внедрению в практику цитологических исследований электронного микроскопа. Мембраны, образующие ЭПС, отличаются от плазмолеммы клетки меньшей толщиной (5-7 нм) и большей концентрацией белков, в первую очередь обладающих ферментативной активностью. Различают две разновидности ЭПС (рис. 8): шероховатую (гранулярную) и гладкую (агранулярную). Шероховатая ЭПС представлена уплощенными цистернами, на поверхности которых расположены рибосомы и полисомы. Мембраны гранулярной ЭПС содержат белки, способствующие связыванию рибосом и уплощению цистерн. Особенно хорошо развита шероховатая ЭПС в клетках, специализирующихся на белковом синтезе. Гладкую ЭПС формируют переплетающиеся канальцы, трубочки и небольшие пузырьки. Каналы и цистерны ЭПС этих двух разновидностей не разграничены: мембраны одного типа переходят в мембраны другого типа, формируя в области перехода так называемую переходную (транзиторную) ЭПС.
Основными функциями гранулярной ЭПС являются:
1) синтез на прикреплённых рибосомах белков (секретируемых белков, белков клеточных мембран и специфических белков содержимого мембранных органоидов); 2) гидроксилирование, сульфатирование, фосфорилирование и гликозилирование белков; 3) транспорт веществ в пределах цитоплазмы; 4) накопление как синтезируемых, так и транспортируемых веществ; 5) регуляция биохимических реакций, связанная с упорядоченностью локализации в структурах ЭПС веществ, вступающих в реакции, а также их катализаторов — ферментов.
Гладкая ЭПС отличается отсутствием на мембранах белков (рибофоринов), связывающих субъединицы рибосом. Предполагается, что гладкая ЭПС образуется в результате формирования выростов шероховатой ЭПС, мембрана которых утрачивает рибосомы.
Функциями гладкой ЭПС являются: 1) синтез липидов, включая мембранные липиды; 2) синтез углеводов (гликогена и др.); 3) синтез холестерина; 4) обезвреживание токсических веществ эндогенного и экзогенного происхождения; 5) накопление ионов Са2+; 6) восстановление кариолеммы в телофазе митоза; 7) транспорт веществ; 8) накопление веществ.
Как правило, гладкая ЭПС развита в клетках слабее, чем шероховатая ЭПС, однако в клетках, вырабатывающих стероиды, триглицериды и холестерин, а также в клетках печени, осуществляющих детоксикацию различных веществ, она развита значительно лучше.
Рис. 9. Комплекс Гольджи:
1 – стопка уплощённых цистерн; 2 – пузырьки; 3 – секреторные пузырьки (вакуоли)
Переходная (транзиторная) ЭПС — это участок перехода гранулярной ЭПС в агранулярную ЭПС, который располагается у формирующейся поверхности комплекса Гольджи. Трубочки и канальцы переходной ЭПС распадаются на фрагменты, из которых образуются пузырьки, транспортирующие материал из ЭПС в комплекс Гольджи.
Пластинчатый комплекс (комплекс Гольджи, аппарат Гольджи) — органоид клетки, участвующий в окончательном формировании продуктов её жизнедеятельности (секретов, коллагена, гликогена, липидов и других продуктов), а также в синтезе гликопротеидов. Органоид назван по имени описавшего его в 1898 году итальянского гистолога К. Гольджи. Образован тремя составляющими (рис. 9): 1) стопкой уплощённых цистерн (мешочков); 2) пузырьками; 3) секреторными пузырьками (вакуолями). Зона скопления этих элементов получила название диктиосомы. Таких зон в клетке может быть несколько (иногда несколько десятков и даже сотен). Комплекс Гольджи располагается около ядра клетки, часто вблизи центриолей, реже рассеян по всей цитоплазме. В секреторных клетках он располагается в апикальной части клетки, через которую осуществляется выделение секрета путём экзоцитоза. От 3-х до 30-ти цистерн в виде изогнутых дисков диаметром 0,5-5 мкм образуют стопку. Смежные цистерны разделены пространствами в 15-30 нм. Отдельные группы цистерн в пределах диктиосомы отличаются особым составом ферментов, определяющих характер биохимических реакций, в частности процессинга белка и др.
Второй составляющий элемент диктиосомы — пузырьки представляют собой сферические образования диаметром 40-80 нм, умеренно плотное содержимое которых окружено мембраной. Пузырьки формируются путём отщепления от цистерн.
Третий элемент диктиосомы — секреторные пузырьки (вакуоли) представляют собой относительно крупные (0,1-1,0 мкм) сферические мембранные образования, содержащие секрет умеренной плотности, претерпевающий конденсацию и уплотнение (вакуоли конденсации).
Комплекс Гольджи отчётливо поляризован по вертикали. В нём выделяют две поверхности (два полюса):
1) цис-поверхность, или незрелую поверхность, которая имеет выпуклую форму, обращена к эндоплазматической сети (ядру) и связана с отделяющимися от неё мелкими транспортными пузырьками;
2) транс-поверхность, или поверхность, обращённую к плазмолемме вогнутой формы (рис. 8), со стороны которой от цистерн комплекса Гольджи отделяются вакуоли (секреторные гранулы).
Основными функциями комплекса Гольджи являются: 1) синтез гликопротеинов и полисахаридов; 2) модификация первичного секрета, его конденсация и упаковка в мембранные пузырьки (формирование секреторных гранул); 3) процессинг молекул (фосфорилирование, сульфатирование, ацилирование и т.п.); 4) накопление секретируемых клеткой веществ; 5) образование лизосом; 6) сортировка синтезированных клеткой белков у транс-поверхности перед их окончательным транспортом (производится посредством рецепторных белков, распознающих сигнальные участки макромолекул и направляющих их в различные пузырьки); 7) транспорт веществ: из транспортных пузырьков вещества проникают в стопку цистерн комплекса Гольджи с цис-поверхности, а выходят из неё в виде вакуолей с транс-поверхности. Механизм транспорта объясняют две модели: а) модель перемещения пузырьков, отпочковывающихся от предшествующей цистерны и сливающихся с последующей цистерной последовательно в направлении от цис-поверхности к транс-поверхности; б) модель перемещения цистерн, основанная на представлении о непрерывном новообразовании цистерн за счёт слияния пузырьков на цис-поверхности и последующем распаде на вакуоли цистерн, смещающихся к транс-поверхности.
Указанные выше основные функции позволяют констатировать, что пластинчатый комплекс — важнейший органоид клетки эукариот, обеспечивающий организацию и интеграцию внутриклеточного метаболизма. В этом органоиде протекают заключительные этапы формирования, созревания, сортировки и упаковки всех секретируемых клеткой продуктов, ферментов лизосом, а также белков и гликопротеинов поверхностного аппарата клетки и др. веществ.
Органоиды внутриклеточного переваривания. Лизосомы — это мелкие ограниченные элементарной мембраной пузырьки, содержащие гидролитические ферменты. Мембрана лизосом толщиной около 6 нм осуществляет пассивную компартментализацию, временно отделяя гидролитические ферменты (более 30 разновидностей) от гиалоплазмы. В неповреждённом состоянии мембрана устойчива к действию гидролитических ферментов и препятствует их утечке в гиалоплазму. В стабилизации мембраны важная роль принадлежит кортикостероидным гормонам. Повреждение мембран лизосом ведёт к самоперевариванию клетки гидролитическими ферментами.
Мембрана лизосом содержит АТФ-зависимый протонный насос, обеспечивающий закисление среды внутри лизосом. Последняя способствует активизации ферментов лизосом — кислых гидролаз. Наряду с этим мембрана лизосом содержит рецепторы, обусловливающие связывание лизосом с транспортными пузырьками и фагосомами. Мембрана обеспечивает также диффузию веществ из лизосом в гиалоплазму. Связывание части молекул гидролаз с мембраной лизосом ведёт к их инактивации.
Выделяют несколько разновидностей лизосом: первичные лизосомы (гидролазные пузырьки), вторичные лизосомы (фаголизосомы, или пищеварительные вакуоли), эндосомы, фагосомы, аутофаголизосомы, остаточные тельца (рис. 8).
Эндосомами называют мембранные пузырьки, переносящие макромолекулы от поверхности клетки в лизосомы путём эндоцитоза. В процессе переноса содержимое эндосом может не изменяться или претерпевать частичное расщепление. В последнем случае в эндосомы проникают гидролазы или эндосомы непосредственно сливаются с гидролазными пузырьками, вследствие чего среда постепенно закисляется. Эндосомы разделяют на две группы: ранние (периферические) и поздние (перинуклеарные) эндосомы.
Ранние (периферические) эндосомы формируются на ранних этапах эндоцитоза после отделения пузырьков с захваченным содержимым от плазмолеммы. Они располагаются в периферических слоях цитоплазмы и характеризуются нейтральной или слабощелочной средой. В них происходит отщепление лигандов от рецепторов, сортировка лигандов и, возможно, возвращение рецепторов в специальных пузырьках в плазмолемму. Наряду с этим в ранних эндосомах может происходить расщепление ком-
Рис. 10 (А). Схема образования лизосом и их участия во внутриклеточном пищеварении. (Б) Электронная микрофотография среза вторичных лизосом (обозначены стрелками):
1 – образование из гранулярной эндоплазматической сети мелких пузырьков с ферментами; 2 – перенос ферментов в аппарат Гольджи; 3 – образование первичных лизосом; 4 – выделение и использование (5) гидролаз при внеклеточном ращеплении; 6 — фагосомы; 7 – слияние первичных лизосом с фагосомами; 8, 9 – образование вторичных лизосом (фаголизосом); 10 – экскреция остаточных телец; 11 – слияние первичных лизосом с разрушающимися структурами клетки; 12 – аутофаголизосома
плексов «рецептор-гормон», «антиген-антитело», ограниченное расщепление антигенов, инактивация отдельных молекул. В условиях закисления (рН=6,0) среды в ранних эндосомах может происходить частичное расщепление макромолекул. Постепенно, перемещаясь вглубь цитоплазмы, ранние эндосомы превращаются в поздние (перинуклеарные) эндосомы, располагающиеся в глубоких слоях цитоплазмы, окружающих ядро. Они достигают 0,6-0,8 мкм в диаметре и отличаются от ранних эндосом более кислым (рН=5,5) содержимым и более высоким уровнем ферментативного переваривания содержимого.
Фагосомы (гетерофагосомы) — мембранные пузырьки, которые содержат захваченный клеткой извне материал, подлежащий внутриклеточному перевариванию.
Первичные лизосомы (гидролазные пузырьки) — пузырьки диаметром 0,2-0,5 мкм, содержащие неактивные ферменты (рис.10). Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки осуществляют транспорт гидролитических ферментов из пластинчатого комплекса к органоидам эндоцитозного пути (фагосомам, эндосомам и т.п.).
Вторичные лизосомы (фаголизосомы, пищеварительные вакуоли) — пузырьки, в которых активно осуществляется внутриклеточное переваривание посредством гидролаз при рН≤5. Их диаметр достигает 0,5-2 мкм. Вторичные лизосомы (фаголизосомы и аутофаголизосомы) формируются путём слияния фагосомы с эндосомой или первичной лизосомой (фаголизосомы) либо путём слияния аутофагосомы (мембранного пузырька, содержащего собственные компоненты клетки) с первичной лизосомой (рис. 10) или поздней эндосомой (аутофаголизосомы). Аутофагия обеспечивает переваривание участков цитоплазмы, митохондрий, рибосом, фрагментов мембран и т.п. Убыль последних в клетке компенсируется их новообразованием, что ведёт к обновлению («омоложению») клеточных структур. Так, в нервных клетках человека, функционирующих многие десятилетия, большинство органоидов обновляется в течение 1 месяца.
Разновидность лизосом, содержащих непереваренные вещества (структуры), названа остаточными тельцами. Последние могут длительно находиться в цитоплазме или выделять своё содержимое путём экзоцитоза за пределы клетки (рис. 10). Распространённым видом остаточных телец в организме животных являются липофусциновые гранулы, представляющие собой мембранные пузырьки (0,3-3 мкм), содержащие труднорастворимый коричневый пигмент липофусцин.
Пероксисомы представляют собой мембранные пузырьки диаметром до 1,5 мкм, матрикс которых содержит около 15 ферментов (рис. 8). Среди последних наиболее важны каталаза, на которую приходится до 40% общего белка органоида, а также пероксидаза, оксидаза аминокислот и др. Пероксисомы образуются в эндоплазматическом ретикулуме и обновляются каждые 5-6 дней. Наряду с митохондриями, пероксисомы являются важным центром утилизации кислорода в клетке. В частности, под воздействием каталазы распадается перекись водорода (Н2О2), образующаяся в ходе окисления аминокислот, углеводов и др. веществ клетки. Таким образом, пероксисомы защищают клетку от повреждающего эффекта перекиси водорода.
Органоиды энергетического обмена. Митохондрии описаны впервые Р. Келликером в 1850 году в мышцах насекомых под названием саркосом. Позднее они изучались и описывались Р. Альтманом в 1894 году как «биопласты», а в 1897 году К. Бенда назвал их митохондриями. Митохондрии представляют собой мембранные органоиды, обеспечивающие клетку (организм) энергией. Источником запасаемой в виде фосфатных связей АТФ энергии являются процессы окисления. Наряду с этим митохондрии участвуют в биосинтезе стероидов и нуклеиновых кислот, а также в окислении жирных кислот.
М
Рис. 11. Схема строения митохондрии:
1 – наружная мембрана; 2 – внутренняя мембрана; 3 – кристы; 4 – матрикс
итохондрии имеют эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Их размеры составляют 0,2-2 мкм в ширину и 2-10 мкм в длину. Количество митохондрий в различных клетках варьирует в широких пределах, достигая в наиболее активных 500-1000. В клетках печени (гепатоцитах) их число составляет около 800, а занимаемый ими объем равен примерно 20% объема цитоплазмы. В цитоплазме митохондрии могут располагаться диффузно, однакообычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонема спермия). Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны — кристы (рис. 11, 12).
Н
Рис. 12. Электронная фотография митохондрии (поперечный разрез)
аружная мембрана митохондрий сходна с плазмолеммой. Она отличается высокой проницаемостью, обеспечивая проникновение молекул с массой менее 10 килодальтон из цитозоля в межмембранное пространство митохондрий. Наружная мембрана содержит порин и другие транспортные белки, а также рецепторы, распознающие переносимые белки в зонах слипания наружной и внутренней мембран.
Межмембранное пространство митохондрий шириной 10-20 нм содержит небольшое количество ферментов. Его ограничивает изнутри внутренняя мембрана митохондрий, содержащая транспортные белки, ферменты дыхательной цепи и сукцинатдегидрогеназу, а также комплекс АТФ-синтетазы. Внутренняя мембрана характеризуется низкой проницаемостью для мелких ионов. Она формирует складки толщиной 20 нм, которые располагаются чаще всего перпендикулярно продольной оси митохондрий, а в некоторых случаях (мышечные и др. клетки) — продольно. С повышением активности митохондрий количество складок (их общая площадь) возрастает. На кристах находятся оксисомы — грибовидные образования, состоящие из округлой головки диаметром 9 нм и ножки толщиной 3 нм. В области головки происходит синтез АТФ. Процессы окисления и синтеза АТФ в митохондриях разобщены, из-за чего не вся энергия накапливается в АТФ, рассеиваясь частично в виде тепла. Такое разобщение наиболее выражено, например, в бурой жировой ткани, используемой для весеннего «разогрева» находившихся в состоянии «зимней спячки» животных.
Внутренняя камера митохондрии (область между внутренней мембраной и кристами) заполнена матриксом (рис. 11, 12), содержащим ферменты цикла Кребса, ферменты белкового синтеза, ферменты окисления жирных кислот, митохондриальную ДНК, рибосомы и митохондриальные гранулы.
Митохондриальная ДНК представляет собственный генетический аппарат митохондрий. Она имеет вид кольцевой двухцепочечной молекулы, в которой содержится около 37 генов. Митохондриальная ДНК отличается от ядерной ДНК низким содержанием некодирующих последовательностей и отсутствием связей с гистонами. Митохондриальная ДНК кодирует иРНК, тРНК и рРНК, однако обеспечивает синтез только 5-6% митохондриальных белков (ферментов системы транспорта ионов и некоторых ферментов синтеза АТФ). Синтез всех других белков, а также удвоение митохондрий контролируются ядерной ДНК. Большая часть рибосомальных белков митохондрий синтезируется в цитоплазме, а затем транспортируется в митохондрии. Наследование митохондриальной ДНК у многих видов эукариот, включая человека, происходит только по материнской линии: митохондриальная ДНК отца исчезает при гаметогенезе и оплодотворении.
Митохондрии имеют относительно короткий жизненный цикл (около 10 суток). Разрушение их происходит путём аутофагии, а новообразование — путём деления (перешнуровки) предшествующих митохондрий. Последнему предшествует репликация митохондриальной ДНК, которая происходит независимо от репликации ядерной ДНК в любые фазы клеточного цикла.
У прокариот митохондрии отсутствуют, и их функции выполняет клеточная мембрана. Согласно одной из гипотез, митохондрии произошли из аэробных бактерий в результате симбиогенеза. Существует предположение об участии митохондрий в передаче наследственной информации.
Источник: StudFiles.net