Мужская гамета без жгутика называется
Бесполое размножениерастений и грибов осуществляется спорами, то есть специализированными клетками, служащими для размножения и расселения (рис. 42).

Рис. 42. Споры некоторых агариковых грибов.
Количество спор, образуемых одним растительным или грибным организмом, может быть весьма велико. Споры не дифференцированы в половом отношении. У водорослей и грибов они диплоидны или гаплоидны. Споры высших растений всегда гаплоидны. У части водорослей и многих грибов споры снабжены ундулиподиями (жгутиками) и подвижны – такие споры называются зооспорами.
Споры наземных растений обычно не имеют приспособлений для активного движения и защищены от высыхания твердой клеточной оболочкой, состоящей из двух слоев – тонкого, проницаемого внутреннего слоя (интины) и толстого, непроницаемого для воды и газов внешнего слоя (экзины).
Организм, производящий споры, получил название спорофита,а процесс образования спор – спорогенеза.У растений споры образуются в спорангиях (если зооспоры – то в зооспорангиях). Спорангий водорослей (низшие растения) обычно одноклеточный; содержимое этой единственной клетки делится, в результате чего формируются споры, которые выходят наружу при вскрывании оболочки спорангия. У высших растений спорангий – многоклеточный орган, имеющий одно- или многослойную стенку. Внутри многоклеточного спорангия формируется образовательная ткань – археспорий (от греческого «архе» – начало), из которой в дальнейшем образуются споры. Из клеток археспория споры возникают в результате мейоза. На гаплоидном организме (некоторые грибы) споры образуются в результате митотического деления. При прорастании такие споры воспроизводят новую особь, идентичную материнской. На диплоидном организме формированию спор предшествует мейоз. Из спор, образовавшихся в результате мейоза, при прорастании возникает гаплоидный организм, который будет размножаться половым способом (за счет образования гамет) – гаметофит (заросток),не идентичный диплоидному материнскому спорофиту.
У низших растений, мхов и даже у многих современных высших сосудистых споровых растений (хвощей, плаунов и папоротников) все споры одинаковы по размерам и физиологическим особенностям. Это равноспоровые организмы. У многих растений (некоторые плауны, папоротники, все голосеменные и цветковые) на одной и той же особи или на разных особях одного вида образуются споры двух типов, различные по размерам и физиологическим особенностям.
кие растения называются разноспоровыми (некоторые плауны и папоротники, все голосеменные и покрытосеменные). Относительно более мелкие споры (микроспоры) (рис. 43) образуются в микроспорангии, а более крупные (мегаспоры) – в мегаспорангии. Микроспоры, прорастая, дают начало однополому мужскому гаметофиту (мужской особи), на котором возникают мужские половые органы – антеридии. Мегаспоры при прорастании образуют женский гаметофит (женскую особь), несущий женские половые органы – архегонии.Разноспоровость эволюционно возникла у высших растений (некоторые плауновидные и папоротники, все голосеменные и покрытосеменные).
Рис. 43. Микроспоры цветковых растений . А – магнолия, Б – сусак, В – джузгун, Г – пион, Д – лютик, Е – истод.
Половое размножение – это такой тип размножения, при котором новые особи образуются в результате полового процесса. Для полового процесса необходимы, как правило, две родительские особи, продуцирующие два физиологически различных типа половых клеток (гамет)с перекомбинированными родительскими хромосомами, которые сливаются и формируют зиготу. Из зиготы в последующем развивается новая дочерняя особь
Организмы, на которых формируются гаметы, называются гаметофитами, а процесс образования гамет – гаметогенезом.У большинства растений и грибов гаметы возникают в особых органах – гаметангиях.Гаметы всегда гаплоидны. Гаметофиты равноспоровых растений обычно обоеполы и формируют как мужские, так и женские гаметангии. У разноспоровых растений обычно из микроспор вырастают гаметофиты с мужскими гаметангиями, а из мегаспор – гаметофиты с женскими гаметангиями.
В связи с тем, что разные растения способны вырабатывать различные по величине и подвижности типы гамет, различают несколько типов полового процесса. В простейшем случае у некоторых одноклеточных водорослей и грибов, лишенных твердой оболочки, сливаются целые одноклеточные организмы. Такой половой процесс называется хологамией.
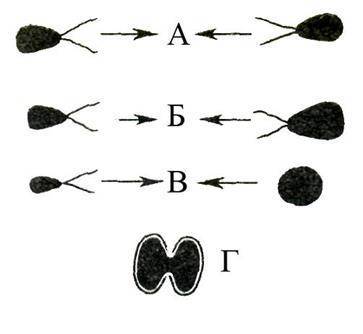
Если в половом процессе участвуют специализированные половые клетки – гаметы, то такой процесс называется гаметогамией (рис. 46). Гаметы, образующиеся в гаметангиях, могут быть морфологически однотипны, тогда их называют изогаметами. Попарное их слияние основано лишь на физиологическом различии, а половой процесс такого типа называют изогамным. Изогамия встречается у некоторых водорослей и у очень немногих грибов. Если подвижные гаметы различаются по величине (одна превосходит другую в несколько раз), то слияние таких гамет обусловливает гетерогамный половой процесс. У некоторых грибоподобных организмов (оомицеты), водорослей и у всех высших растений половой процесс оогамный.Женская гамета (яйцеклетка) при оогамной форме полового процесса неподвижна, лишена жгутиков (ундулиподиев), имеет крупные размеры и большой запас питательных веществ.
жская гамета (сперматозоид) – маленькая, подвижная, со жгутиками (ундулиподиями), либо без жгутиков (тогда она называется спермий); состоит из крупного ядра и очень небольшого количества цитоплазмы. Неподвижная яйцклетка образуется либо в гаметангии, называемом оогонием (у водорослей и некоторых грибов), либо в архегонии (высшие растения, исключая цветковые).
Оогоний (от греческого «оон» – яйцо, «гоне» – рождение) обычно состоит из одной клетки, реже (у харовых водорослей) многоклеточный. Архегоний(от греческого «архе» – начало) – женский половой орган высших архегониальных растений – мхов, плаунов, хвощей, папоротников и голосеменных. Это всегда многоклеточное образование, состоящее из расширенного брюшка, в котором помещается яйцеклетка, и удлиненной шейки. Над яйцеклеткой находится брюшная канальцевая клетка. Наружные клетки архегония стерильны и образуют однослойную стенку. По каналу шейки, заполненному слизью, сперматозоиды проникают в брюшко и один из них сливается с яйцеклеткой.
Сперматозоиды у споровых растений образуются в гаметангиях, называемых антеридиями. Антеридий(от греческого «антерос» – цветущий) – это одноклеточный (у большинства низших растений) или многоклеточный (у высших споровых растений) мужской половой орган, имеющий у разных систематических групп различную (округлую или овальную) форму. Сперматозоиды, созревшие в антеридиях, могут достичь яйцеклетки только в присутствии капельно-жидкой воды. Наличие ее – обязательное условие для осуществления процесса оплодотворения у всех групп растительных организмов, исключая семенные растения. У цветковых растений произошла полная редукция гаметангиев, однако половые клетки (спермии и яйцеклетки) образуются и участвуют в половом процессе.
Помимо типичного полового процесса, в котором обязательно участвуют две гаметы, существует особый тип полового процесса, при котором зародыш развивается из неоплодотворенной яйцеклетки. Это явление у растений чаще всего известно под названием апомиксиса(от греческого «апо» – от (без) и «миксис» – смешение). Апомиксис широко встречается у многих покрытосеменных растений (часть розоцветных, сложноцветных), особенно у ряда культурных растений (свекла, хлопчатник, лен, табак).
Рис. 44.Различные типы полового размножения у растений, водорослей и грибов (схема). А – изогамия (характерна для некоторых водорослей); Б – гетерогамия (некоторые водоросли); В – оогамия (некоторые водоросли, все высшие растения); Г – конъюгация (некоторые водоросли).
Половая и бесполая формы размножения имеют определенные биологические преимущества. При половом размножении достигается комбинация наследственного материала родительских форм. Образующаяся при половом воспроизведении особь генетически нетождественна ни одному из своих родителей.
ловой процесс обеспечивает генетическую изменчивость организмов из поколения в поколение, а половое размножение дает виду преимущества, реализующиеся при естественном отборе. При бесполом размножении наследственные особенности передаются без изменений и могут легко закрепляться в ряду поколений. Однако чаще всего у растений осуществляется чередование полового и бесполого поколений. При чередовании поколений гаметофит закономерно сменяется спорофитом, который затем вновь сменяется следующим в череде поколений гаметофитом.
Гаметофит и спорофит могут быть одинаковыми как морфологически, так и по продолжительности жизни (изоморфное чередование поколений) (у многих водорослей) или резко различными (гетероморфное чередование) (у некоторых водорослей и всех высших растений).
Таким образом, у разных групп водорослей встречаются обе формы чередования поколений. Для высших растений характерно только гетероморфное чередование. Оба поколения развиваются либо независимо как самостоятельные особи, либо одно поколение «поселяется» на другом. Так, у мхов спорофит развивается на зеленом гаметофите. У семенных растений крайне редуцированный бесхлорофилльный женский гаметофит не покидает спорофит. У всех высших растений, кроме мхов, преобладает спорофит, а гаметофит развит слабее и недолговечен. Чередование поколений связано со сменой ядерных фаз – гаплоидной (n) и диплоидной (2n). Эта смена обеспечивается благодаря мейозу, протекающему у всех высших растений при спорогенезе, и половому процессу.
Диплоидный спорофит высших растений всегда производит гаплоидные мейоспоры. Из них вырастает гаплоидный гаметофит, продуцирующий гаплоидные митогаметы (половые клетки, образованные в процессе митоза). При слиянии гамет восстанавливается диплоидное число хромосом в зиготе, из которой вырастает диплоидный спорофит.
Задание 1
1. Заполните таблицу предоставив общую характеристику бесполого и полового размножения.
Таблица 3 — Общая характеристика бесполого и полового размножения
| Показатель | Способ размножения | |
| бесполое | половое | |
| Клеточные источники наследственной информации | ||
| Родители | ||
| Потомство | ||
| Главный клеточный механизм | ||
| Эволюционное значение |
Задание 2
1. Спорообразование у плесневого гриба. Нанесите пипеткой каплю воды на предметное стекло, возьмите пинцетом несколько нитей мицелия плесневого гриба, положите их в каплю воды на предметное стекло и накройте покровным стеклом.
ссмотрите препарат под малом и большом увеличением микроскопа. В поле зрения видны ветвящиеся полупрозрачные и тонкие нити – гифы. Найдите среди гиф спорангии – круглые черные коробочки на длинной ножке (спорангиецосце), заполненные мелкими спорами округлой формы. Некоторые из спорангиев могут оказываться разорванными, и в таких случаях вокруг них видны высыпавшиеся мелкие округлые клетки – споры. Внутри спорангия обнаруживается колонка – утолщенный вырост спорангиеносца; форма и размеры колонки постоянны у определенных видов грибов. Зарисуйте мицелий плесневого гриба, на рисунке обозначьте: мицелий, гифы, спорангий, споры.
Источник: poznayka.org
Сперматозоиды растений
Строение
Так как оплодотворение проходит не в жидкой среде, в большинстве покрытосеменных сперматозоиды лишены жгутиков и не способны к активному движению. Сперматозоид, что два или много жгутиков у некоторых грибов, бурых и зеленых водорослей, плаунов, хвощей, папоротников, мхов и некоторых голосеменных. Собственно сперматозоиды имеют мелкие размеры и лишены целюлюзнои оболочки (исключая саговая, в которых они достигают до 300 мкм в диаметре), морфологически отличаются от животных сперматозоидов. В целом джгутиконосний сперматозоид имеет следующее строение: головка, промежуточная часть и хвост. Ядро большое количество цитоплазмы невелика. Жгутик может быть различной длины и с выростами (ретронемамы или мастигонемамы) или без них, занимать переднее, заднее или оба положения.
Сперматогенез
Образование и созревания сперматозоидов у растений происходит в специализированных органах — антеридиях. В водорослей (кроме харовых), например, эти органы являются одноклеточными. В Красных водорослей, например, спермаций, что лишен жгутика, образуется в сперматангии.
Оплодотворение
Оплодотворение в некоторых голосеменных растений и водорослей происходит в водной среде. При этом женская половая клетка выделяет определенные химические вещества, приманивают спермации.
Сперматозоиды животных
Строение
Сперматозоиды у животных содержатся в биологической жидкости, называется спермой. Типичный животный сперматозоид состоит из головки, шейки, тела (или промежуточной части) и хвоста.
Головка содержит гаплоидны ядро, содержащее наследственную информацию, акросома (спереди от ядра и сразу под плазматической мембраной), которая содержит ферменты для разрушения оболочки яйцеклетки. В шейке находится базальное тело жгутика, имеет митохондрии. Тело сформировано так называемой митохондриальной спиралью — комплексом, составляют митохондрии, хвост непосредственно сформирован плотными белковыми фибриллами и фибриллярных футляром. У насекомых головка сперматозоида сильно удлиненная, иногда спирально закрученная и без заметного выделения тела переходит в длинный хвост (жгутик). Преимущественно этот хвост занимает заднее положение.
Сперматогенез
У человека и высших позвоночных животных образования мужских гамет осуществляется в мужской половой железе — яичке (в извитых семенных канальцах) и включает 4 последовательные фазы: размножение, рост, созревание и формирование. Процесс образования одного сперматозоида длится примерно 72 суток.
- Размножение происходит путем ряда мейотического деления, после которого будущая гамета (Сперматиды) получает гаплоидный набор хромосом;
- Рост, созревание и формирование проходит в эпителии извитых семенных канальцев.
Оплодотворение
После введения сперматозоидов в половых путей самки, они могут двигаться, используя жгутик, со скоростью до 3-4 мм в минуту.
Для оплодотворения яйцеклетки достаточно одной мужской гаметы (моноспермия). Вместе с тем возможны случаи одновременного оплодотворения несколькими гаметами (полиспермия), например, у птиц. После проникновения головки сперматозоида происходит блокировка оболочки яйцеклетки к проникновению других мужских гамет.
У человека примерно через 1-2 часа после полового акта с эякуляцией первые сперматозоиды достигают ампулярной части фаллопиевых труб (той части, где происходит оплодотворение).
Находиться и сохранять способность к заплидення в половых путях самки сперматозоиды могут от 2 до 7 суток.
Источник: info-farm.ru
Содержание раздела
ЭНЦИКЛОПЕДИЯ
Как и все живые организмы, растения размножаются. Существует три способа размножения растений — вегетативный, бесполый и половой. При вегетативном способе новая особь образуется из части вегетативных органов растений, т. е. листа, стебля или корня. Иногда новая особь возникает даже из отдельной клетки того или иного вегетативного органа растения. При бесполом размножении у растений образуются особые клетки (споры), из которых вырастают новые самостоятельно живущие особи, сходные с материнской. Этот способ размножения свойствен некоторым водорослям (рис. 1) и грибам (см. ст. «Грибы»). Половое размножение принципиально отличается от вегетативного и бесполого. Половой процесс в растительном мире крайне разнообразен и часто очень сложен, но по существу сводится к слиянию двух половых клеток — гамет, мужской и женской.
Рис. 1. Бесполое размножение хламидомонады: 1 — хламидомонада; 2—образование зооспор; 3 — выбегание зооспор.
Гаметы возникают в определенных клетках или органах растений. В некоторых случаях гаметы одинаковы по размерам и форме, обе имеют жгутики и потому подвижны. Это изогамия (рис. 3). Иногда они несколько отличаются друг от друга размерами. Это гетерогамия (рис. 2). Но чаще — при так называемой оогамии — размеры гамет резко различны: мужская гамета, называемая сперматозоидом, небольшая, подвижная, а женская — яйцеклетка — неподвижная и крупная (рис. 4). Процесс слияния гамет называется оплодотворением. Гаметы имеют в своем ядре по одному набору хромосом, а в образовавшейся после слияния гамет клетке, которая называется зиготой, число хромосом удваивается (см. ст. «Клетка»). Зигота прорастает и дает начало новой особи.
рис. 2. Гетерогамия у хламидомонады. Обе гаметы подвижны, но отличаются размерами.
Половой процесс осуществляется у растения в определенное время и на определенном этапе развития. На протяжении своего развития растение может размножаться также и бесполым путем (с образованием спор) и вегетативно.
Рис. 3. Изогамия у хламидомонады: 1 — образование гамет; 2 — гаметы; 3 — слияние гамет; 4 — зигота (видны жгутики); 5 — зигота со сброшенными жгутиками; 6,7,8 — прорастание зиготы и образование четырех новых особей хламидомонады.
Половое размножение возникло в растительном мире в процессе эволюции. У синезеленых водорослей его еще нет. Они размножаются только вегетативно, путем деления клетки на две. У большинства водорослей и грибов, а также у всех высших наземных растений половой процесс отчетливо выражен. Половое размножение очень важно для организма, так как благодаря слиянию отцовской и материнской клеток создается новый организм, который имеет большую изменчивость, лучше приспособлен к условиям окружающей среды.
Наиболее прост процесс полового размножения у одноклеточных водорослей, например у хламидомонад. Хламидомонада размножается как бесполым, так и половым путем. При бесполом размножении хламидомонада теряет жгутики и делится на 2, 4 (реже 8) клетки-споры. Каждая из них снабжена двумя жгутиками. Это зооспоры. После разрушения оболочки клетки, внутри которой они сформировались, зооспоры разбегаются и дорастают до размеров материнской клетки (рис. 1).
Рис. 4 Оогамия у хламидомонады: 1 — неподвижная яйцеклетка, а — сперматозоид; 2 — оплодотворение (слияние сперматозоида с яйцеклеткой); 3 — зигота, покрывшаяся толстой оболочкой.
При половом размножении (рис. 3) содержимое клетки хламидомонады делится и образуется большое число гамет (32 или даже 64). Потом оболочка материнской клетки прорывается, и гаметы, имеющие по два жгутика, выходят в воду, плавают, соединяются попарно своими носиками, где расположены жгутики, и, наконец, полностью сливаются друг с другом. У большинства хламидомонад трудно отличить, какие гаметы мужские, какие женские. Они одинаковы по форме и одинаково подвижны. Однако есть некоторые виды хламидомонад, которые образуют неподвижные крупные женские гаметы (яйцеклетки), а другие особи — мелкие подвижные мужские гаметы (сперматозоиды). После слияния гамет жгутики исчезают, образуется зигота, которая сразу же покрывается оболочкой (рис. 4).
Через некоторое время зигота прорастает. Первое деление ее ядра редукционное — особое деление ядра, при котором число хромосом в клетке уменьшается вдвое (см. ст. «Клетка»). В результате второго деления каждого из ядер образуются 4 клетки с одним набором хромосом в их ядрах. Оболочка зиготы лопается, и новые клетки выходят в воду, плавают при помощи двух жгутиков. Достигнув размеров материнской клетки, они могут снова размножаться бесполым и половым путем.
Период от появления гаметы и до образования новых гамет называют циклом развития растения. У некоторых многоклеточных водорослей обе половые клетки неподвижны. Так, у спирогиры при половом процессе содержимое одной клетки переливается в другую, где и происходит слияние их цитоплазмы, ядер и образуется зигота. У других многоклеточных водорослей процесс полового размножения более сложен.
Этот процесс очень разнообразен у наземных растений. У мхов, папоротников, голосеменных, например у хвойных, а также у цветковых растений он происходит по-разному. В связи с выходом из воды на сушу у мхов, папоротников, хвощей, плаунов и у семенных растений сильно усложнилось не только строение, но и процесс размножения. У них, как и у многих водорослей, наблюдается правильное чередование бесполого и полового поколений. Зигота прорастает без редукционного деления, и развивающаяся из нее особь имеет двойной набор хромосом.
Рис. 5. Развитие мхов: 1 — взрослое растение; 2 — прорастание споры (2 фазы); 3 — предросток с молодыми растениями мха; 4 — мужское растение с антеридиями; 5 — антеридий; 6 — сперматозоид; 7 — архегоний, готовый к оплодотворению; 8 — оплодотворенный архегоний; 9 — первые стадии развития спорогона; 10 — молодой спорогон внедрился ножкой в верхушку стебля; 11 — зрелая коробочка с клетками (а), из которых в дальнейшем образуются споры; 12 — подготовка клеток к редукционному делению; 13 — гаплоидные споры.
Это бесполое поколение, так как на таком растении образуются споры. При формировании их происходит редукционное деление, в результате которого спора получает один набор хромосом. Прорастающая спора дает начало половому поколению — организму, который образует половые клетки — гаметы. Все клетки этой особи несут один набор хромосом. Образующаяся в результате оплодотворения гамет зигота снова прорастает и дает бесполое поколение (с двойным набором хромосом). В цикле развития растения может преобладать половое (мхи) или бесполое (остальные высшие растения) поколение.
Рассмотрим цикл развития мха кукушкин лен (рис. 5). Стебли этого мха небольшие, крепкие, с многочисленными мелкими, узкими, жесткими листьями. На верхушке некоторых из этих стеблей развиваются коробочки, сидящие на удлиненной ножке и покрытые колпачком, как капюшоном (1). Коробочка на ножке называется спорогоном. В самой коробочке, покрытой крышечкой, образуется масса спор. Они мелкие, как пыль. При их образовании происходит редукционное деление, и споры
получают по одному набору хромосом (гаплоидные споры). После их созревания колпачок сбрасывается, крышечка коробочки отскакивает, и споры высыпаются наружу. Споры попадают на почву и при влажной погоде прорастают (2). Образуется зеленая ветвистая многоклеточная нить, стелющаяся по влажной поверхности почвы, а погруженные в почву бесцветные нити всасывают почвенные растворы. Зеленая нить называется предростком (3). На предростке образуются почки. Из почек развиваются новые стебли кукушкина льна.
На верхушках одних стеблей появляются многоклеточные небольшие кувшинообразные выросты, сидящие на небольшой ножке. Это женские половые органы, или архегоний. В их нижней расширенной части помещается одна неподвижная яйцеклетка. На верхушках других стеблей мха вырастают многоклеточные, но одностенные удлиненные мешочки — антеридии (4, 5). Внутри них образуются многочисленные мелкие мужские гаметы — сперматозоиды (6). Во время дождя или обильной росы мешочки лопаются вверху, и из них в слизистой массе выступает множество сперматозоидов, снабженных двумя жгутиками, с помощью которых они двигаются к верхушке тех стеблей кукушкина льна, где находятся архегоний. Проникнув через шейку архегония внутрь, сперматозоид сливается с яйцеклеткой (7, 8). В результате образуется зигота, которая прорастает без редукции хромосом здесь же, на верхушке стебля кукушкина льна, образуя бесполое поколение — спорогон, состоящий из коробочки и ножки. Ножка спорогона внедряется в ткани стебля и высасывает из него питательные вещества (9, 10), В коробочке спорогона образуются споры (11, 12, 13). Таков цикл развития мхов. У них преобладает половое поколение (само растение мха).
Рис. 7. Папоротник: 1 — внешний вид папоротника (бесполое поколение); 2 — долька листа с нижней стороны (видны сорусы, одетые покрывалом); 3 — разрез соруса, а — спорангии, б — покрывало; 4 — отдельный спорангий, из которого высыпаются споры.
Теперь рассмотрим цикл развития у папоротника щитовника, распространенного по тенистым местам в лиственных лесах (рис. 6, 7). Из верхушки его подземного корневища ежегодно вырастает пучок перистосложных листьев (1). На нижней поверхности листьев вдоль средней жилки легко заметить кучки спорангиев — так называемые сорусы, закрытые покрывалом, напоминающим в поперечном разрезе раскрытый зонтик (2, 3). Двояковыпуклый спорангий имеет вид чечевицы и расположен на ножке. Внутри спорангия масса мелких спор, возникших в результате редукционного деления.
Рис. 8. Половое размножение папоротника: 1 — заросток, а — архегонии, б — антеридии, в — ризоиды; 2 — из зрелого антеридия выходят сперматозоиды; 3 — архегонии, готовый к оплодотворению; 4 — заросток с молодым спороцоном, а — первый лист, б — корешок.
В сухую погоду, когда споры уже созрели, спорангий вскрывается (4). Высыпавшиеся от резкого толчка споры рассеиваются и попадают на поверхность почвы. Попав в благоприятные условия — тепло и влагу, спора прорастает и образует очень маленькую (2—5 мм в диаметре) тонкую зеленую пластинку сердцевидной формы — заросток (рис. 8). Своей нижней поверхностью заросток плотно прижимается к земле благодаря ризоидам, всасывающим из почвы растворы минеральных солей. Заросток папоротника обоеполый: на его нижней поверхности расположены женские (архегонии) и мужские (антеридии) половые органы. Заросток и представляет собой половое поколение папоротника. Во время дождя или обильной росы многожгутиковые сперматозоиды выходят из антеридия в воду и направляются к архегонию. Там происходит оплодотворение, после чего получается зигота — клетка с двойным набором хромосом. Она прорастает здесь же, на заростке, и образуется зародыш. Разрастаясь все больше и больше, он образует все части взрослого растения: стебель, лист, корни. Затем на нижней поверхности листа взрослого растения снова появляются сорусы со спорангиями.
Рис. 9. Мужские и женские шишки сосны: 1 — собрание мужских шишек; 2 — молодая женская шишка на верхушке побега; 3 — прошлогодняя женская шишка; 4 — чешуйки мужской шишки (вид сбоку и снизу); 5 — чешуйки женской шишки (вид с наружной и внутренней стороны); 6 — чешуйка зрелой женской шишки с двумя крылатыми семенами, отдельно изображены крылышко и семя; 7 — продольный разрез мужской шишки, на оси расположены чешуйки со спорангиями; 8 — отдельный спорангий со спорами (пылинками).
Таким образом, в цикле развития папоротника преобладает бесполое поколение, формирующее спорангии со спорами (сам папоротник). Половое поколение (заросток) имеет незначительные размеры и существует недолго. Оба поколения существуют раздельно, самостоятельно. Подобным же образом размножаются хвощи и плауны, которые вместе с папоротниками объединяют в класс папоротникообразных.
По-другому идет размножение у семенных растений. У них рассеиваются не споры, а семена. Однако и у этих растений тоже образуются споры, а также двоякого рода клетки полового размножения: мужские и женские.
У голосеменных, например у сосны, ели, образуются мужские и женские шишки (рис. 9). Мужские шишки собраны в тесные группы у основания побегов, развивающихся в этом году. Женские шишки сидят поодиночке сначала на верхушке побега, а затем вследствие роста побега оказываются у его основания. Мужская шишка состоит из чешуек, тесно сидящих на ее оси. На нижней поверхности чешуек расположены два спорангия. Внутри спорангия путем редукционного деления развивается огромное количество спор (пылинок). Содержимое каждой пылинки состоит из густой плазмы и ядра. Пылинка одета оболочкой, образующей два пузырчатых сетчатых воздушных мешка (рис. 10). Это приспособление способствует разносу ветром пылинок, высыпавшихся из лопнувшего пыльника. Пылинка прорастает в мужской заросток. При этом ядро ее делится, и образуются две быстро разрушающиеся клетки и две сохраняющиеся дольше клетки — более крупная вегетативная и более мелкая антеридиальная. В таком двуклеточном состоянии пылинка переносится ветром и попадает на поверхность женской шишки, где и происходит процесс оплодотворения.
Рис. 10. Пылинка (спора) и развитие мужского заростка сосны: 1 — зрелая спора, а — воздушные мешки; 2—5 — прорастание споры и образование мужского заростка, б, в — рано исчезающие клетки заростка, г — антеридиальная клетка, д — вегетативное ядро (ядро клетки пыльцевой трубки), е — сестринская клетка, ж — генеративная клетка (при дальнейшем ее делении образуются два спермин — мужские гаметы).
Женская шишка состоит из мелких кроющих чешуек, в пазухах которых развиваются крупные мясистые семенные чешуи. У основания последних на их внутренней (верхней) стороне расположены по две овальные семяпочки (рис. 11). В верху семяпочки имеется маленькое отверстие — пыльцевход. В семяпочке одна из клеток, выделяющаяся крупными размерами, делится редукционно, в результате образуются четыре споры. Три из них отмирают, а четвертая приступает к делению. В результате многократного деления образующихся при этом клеток формируется женский заросток, занимающий середину семяпочки. На заростке образуются два небольших архегония очень упрощенного строения с маленькими шейками, в каждом из которых находится по одной яйцеклетке.
Рис. 11. Семяпочка сосны: 1 — продольный разрез семяпочки, о — женский заросток, б — архегонии, в — нуцеллус, г — покров; 2 — верхняя часть семяпочки в продольном разрезе при большем увеличении, а — женский заросток, б — яйцеклетка архегония, в — нуцеллус, г — покров, д — пыльцевход, е — пыльцевая трубка, проросшая через нуцеллус и достигшая женского заростка. В пыльцевой трубке видны 4 ядра (2 спермин, вегетативное ядро и ядро сестринской клетки).
Если теперь разрезать семяпочку вдоль, то можно увидеть, что заросток окружен содержимым семяпочки (нуцеллусом), который, в свою очередь, одет покровом семяпочки. Наверху осталось всего лишь маленькое отверстие — пыльцевход. Через него и попадает перенесенная ветром на верхушку семяпочки пылинка. Она втягивается внутрь семяпочки, где и прорастает на следующее лето. Пылинка образует пыльцевую трубку, внедряющуюся в нуцеллус и растущую по направлению к шейке одного из архегониев. В это же время антеридиальная клетка делится на две. Одна из образовавшихся клеток в дальнейшем разрушается, а другая (генеративная клетка) увеличивается в размере, делится и образует две половые клетки — мужские гаметы, или спермин, не имеющие жгутиков.
Следует, однако, отметить, что у более древних голосеменных растений (гинкго и саговников) имеются подвижные сперматозоиды. Это свидетельствует об их происхождении от папоротникообразных растений.
Достигнув архегония, пыльцевая трубка лопается, и один из спермиев попадает в архегонии и сливается с яйцеклеткой. Происходит оплодотворение, и образуется зигота. Другой спермий вскоре отмирает. Из зиготы формируется зародыш нового растения, питающийся за счет запасных веществ клеток женского заростка (эндосперма). Семяпочка становится теперь семенем. Семя одето плотной кожурой, в которую превратился покров семяпочки.
Рис. 12. Строение и прорастание семени сосны: 1 — строение семени, а — кожура (покров) семени, б — остаток нуцеллуса, в — эндосперм, в середине которого находится зародыш, г — семядоли зародыша, д — подсемядольное колено (зачаточный стебель) е — корешок, ж — подвесок; 2 — прорастание семени сосны, а — кожура семени, б — семядоли, в — подсемядольное колено, г — корень, д — побег с первыми листьями.
Семена созревают к осени. Они сидят в основании чешуи шишки. К осени на второй год своего существования шишка разрастается. Из зеленой она становится бурой, чешуи подсыхают, расходятся, семена выпадают и рассеиваются. Попав в благоприятные условия, семена прорастают и развиваются в новые растения (рис. 12).
В цикле развития сосны, так же как и у папоротникообразных, преобладает бесполое поколение. Половое поколение здесь еще более просто устроено. При этом оно потеряло способность к самостоятельной жизни и развивается внутри тканей бесполого поколения (женский заросток — внутри семяпочки, а мужской заросток — внутри пылинки).
Особенность размножения покрытосеменных (или цветковых) растений — образование цветка как специализированного органа, приспособленного к половому размножению (рис. 13). Внешняя часть цветка состоит из околоцветника, обычно в виде лепестков и чашелистиков. Но главную часть цветка составляют находящиеся в его центре пестик (или пестики) и тычинки, расположенные вокруг пестика. Тычинки состоят из тычиночных нитей и пыльников, а пестик — из одного или нескольких сросшихся между собой завернутыми внутрь краями плодолистиков. В образующейся при этом срастании полости скрыты расположенные обычно по краям плодолистиков одна или несколько семяпочек.
Рис. 13. Размножение у покрытосеменных (цветковых) растений. Продольный разрез цветка (чашелистики и лепестки удалены): а — тычинка, на ее поперечном разрезе видны гнезда пыльника; б — пыльник в продольном разрезе, видна пыльца; в — нить тычинки; г — завязь; д — столбик; е — рыльце пестика; ж — прорастающая на рыльце пылинка; з — пыльцевая трубка, проросшая через ткани рыльца, столбика и достигшая зародышевого мешка семяпочки; и — зародышевый мешок.
В нижней части пестик расширен. Это завязь. Кверху пестик утончается и образует столбик, который заканчивается по-разному устроенным рыльцем, служащим для улавливания и восприятия пыльцы. Плодолистики затем изменяются и принимают большое участие в образовании плода.
Как и у голосеменных, здесь центральную часть семяпочки занимает однородная ткань из живых клеток — нуцеллус. С внешней стороны нуцеллус прикрыт двумя, реже одним покровом. Наверху покровы не смыкаются. Здесь имеется отверстие — пыльцевход. Вскоре после образования нуцеллуса одна из его верхних клеток путем редукционного деления образует четыре споры. Одна из них сильно разрастается и приступает к делению, в результате образуется женский заросток — зародышевый мешок. Остальные три споры отмирают.
Рис. 14. Пыльца покрытосеменного (цветкового) растения и ее прорастание: 1 — пылинка, внутри видны округлое вегетативное ядро — ядро клетки пыльцевой трубки (а) и изогнутая генеративная клетка (б); 2 — через пору в наружной оболочке пылинки вытягивается пыльцевая трубка; 3 — вегетативное ядро опустилось в пыльцевую трубку; 4, 5 — генеративная клетка поделилась, образовались два спермия (в); 6 — зрелые спермин (в).
Женский заросток у покрытосеменных еще более упрощен по сравнению с голосеменными и состоит всего из 8 клеток. Образуется он так. Ядро споры делится на два. Расходясь к полюсам зародышевого мешка, они снова двукратно делятся. Теперь на полюсах уже по четыре ядра. Вскоре от каждой из этих четверок по направлению к центру мешка отделяется по одному ядру. Это полярные ядра. Здесь они сближаются, затем, сливаясь, образуют вторичное (центральное) ядро зародышевого мешка.
Ядра, оставшиеся на полюсах, облекаются цитоплазмой. Образуется по три клетки на каждом из полюсов. Противоположные от семявхода клетки называются антиподами. Три клетки, расположенные близ верхнего конца зародышевого мешка, не одинаковы. Средняя из них представляет собой яйцеклетку, а расположенные по бокам возле нее две меньшие клетки называются вспомогательными. Середина зародышевого мешка заполнена цитоплазмой и вакуолями с вторичным ядром в центре.
Рис. 15. Зародышевый мешок (женский заросток) покрытосеменного (цветкового) растения и двойное оплодотворение: 1 — яйцеклетка; 2 — вспомогательные клетки; 3 — вскрывшаяся пыльцевая трубка. Один из спермиев (4а) сливается с яйцеклеткой; 5 — полярные ядра, сливающиеся со вторым спермием (4б); 6 — три клетки в нижней части зародышевого мешка (антиподы).
В пыльнике тычинки, в каждом из четырех его гнезд образуются споры (пылинки). Они происходят из особых материнских клеток пыльцы в результате редукционного деления их. Содержимое пылинки состоит из крупного ядра и цитоплазмы (рис. 14). Пылинка окружена двумя оболочками: внутренней и внешней. Во внешней оболочке имеются отверстия или утонченные места. Еще в гнезде пыльника в каждой пылинке начинается формирование мужского заростка. Он еще более упрощен в сравнении с голосеменными. Ядро пылинки делится, и образуются две клетки: более крупная — вегетативная и более мелкая — генеративная. После этого пыльник вскрывается, пыльца из него высыпается и с помощью ветра, насекомых или воды, а у некоторых тропических растений при помощи птиц попадает на рыльце пестика. Этот процесс называется опылением.
Около 10% цветковых растений опыляется ветром. Цветки ветроопыляемых растений невзрачны. Они имеют околоцветник в виде пленок, чешуек; нередко он совсем отсутствует, например у злаков, осок, дуба, березы, осины, ольхи. Пыльца этих растений очень мелкая, с гладкой наружной оболочкой. Пыльцы образуется очень много, ведь ветер — ненадежный опылитель. Лишь небольшая часть пылинок попадает на рыльце пестика.
Большинство цветковых растений опыляется насекомыми: пчелами, осами, шмелями, бабочками, мухами. Насекомые посещают цветки из-за сладкого сока (нектара), который выделяется особыми нектарными желёзками, расположенными на лепестках, тычинках или на цветоложе. Венчики цветков насекомоопыляемых растений ярко окрашены и хорошо заметны издали. Пыльца у них более крупная, наружная оболочка пылинок имеет выросты в виде шипов, бугорков, и потому пыльца легко задерживается на рыльце пестика.
Рис. 16. Строение и прорастание семени у клещевины: 1 — проросток (2 стадии развития), а — оболочка семени, б — эндосперм, в середине которого находится зародыш, в — семядоли, г — подсемядольное колено — зачаточный стебель, д — корень; 2 — разрез семени.
Очень важно, чтобы пыльца не попадала на рыльце того же цветка. В случае самоопыления, как заметил еще Ч. Дарвин, получается более слабое потомство. У растений имеются различные приспособления, обеспечивающие перекрестное опыление, при котором пыльца попадает на рыльце другого цветка. Так, у ветроопыляемых растений цветки большей частью раздельнополые: одни цветки содержат только тычинки (тычиночные цветки), другие — только пестики (пестичные цветки). У насекомоопыляемых растений цветки, как правило, обоеполые, имеющие тычинки и пестики. Очень часто тычинки созревают и начинают высыпать пыльцу значительно раньше, чем полностью сформируется пестик. У многих растений пестики созревают раньше тычинок. У некоторых растений, например у примулы, медуницы, незабудки, тычинки и пестики не одинаковы по длине. Наиболее здоровое и сильное потомство вырастает из семян, образовавшихся в результате перенесения пыльцы из цветков с длинными тычинками на рыльце пестиков с длинными столбиками.
Попавшая на рыльце пестика пыльца прорастает (рис. 14). Вегетативная клетка, находящаяся внутри пылинки, разрастается и вытягивается в пыльцевую трубку, которая выходит через отверстие во внешней оболочке пылинки и продвигается в виде тонкой нити сквозь рыхлую ткань рыльца и стенок пестика к семяпочке. Через пыльцевход она направляется к зародышевому мешку.
Во время роста пыльцевой трубки в нее проникает генеративная клетка. Здесь она делится и образует две мужские гаметы (спермин). Достигнув зародышевого мешка, пыльцевая трубка, в которой находятся вегетативное ядро и два спермин, лопается, и содержимое ее изливается в зародышевый мешок (рис. 15). Один из спермиев сливается с яйцеклеткой. Образуется зигота. Второй спермий направляется в середину зародышевого мешка и сливается там со вторичным ядром. Происходит так называемое двойное оплодотворение, составляющее особенность цветковых растений. Честь его открытия в конце XIX столетия принадлежит нашему русскому ученому С. Г. Навашину.
Оплодотворенное вторичное ядро начинает быстро делиться. В результате зародышевый мешок заполняется массой клеток, содержащих питательные вещества (крахмал, масло). Эту используемую для питания зародыша ткань называют эндоспермом. Оплодотворенная яйцеклетка — зигота начинает расти и делиться, в результате чего формируется зародыш, представляющий собой маленькое растение, состоящее из семядолей (двух или одной), подсемя-дольного колена и корня.
Семяпочка тем временем превращается в семя, ее покровы твердеют и образуют кожуру семени (рис. 16). Стенки завязи (плодолистики) разрастаются, становятся сочными или твердыми, кожистыми или деревянистыми. Теперь завязь превращается в плод, надежно защищающий семена. Плоды разносятся животными или ветром, и после разрушения стенок (околоплодника) семена освобождаются. Семя в благоприятных условиях прорастает и дает новое бесполое поколение цветкового растения. Таким образом, в цикле развития покрытосеменных растений также преобладает бесполое поколение.
Как мы уже говорили, низшим растениям, а также мхам и папоротникообразным для полового процесса необходима вода, в которой сперматозоиды активно движутся к яйцеклеткам. Эти растения произрастают или в воде (водоросли), или во влажных, тенистых местах (мхи, папоротники, хвощи, плауны). Половой процесс у семенных и особенно у цветковых растений не связан с водой, недостаток которой так остро ощущается при жизни на суше. Мужские гаметы (спермин) у них доставляются к яйцеклеткам при помощи пыльцевой трубки. Кроме того, семя надежно защищает зародыш. Благодаря этим особенностям семенные и особенно покрытосеменные растения смогли завоевать сушу. Они в настоящее время господствуют на Земле.
перейти к началу страницы
Источник: 2i.su
Тема нашего следующего рассказа это отношения полов. Проследим, как они складываются у растений, принадлежащих к разным систематическим группам.
Водоросли. Для большинства водорослей, которые дали начало высшим растениям, характерно половое размножение. Более того, у названных ниже групп водорослей размножение бесполое отсутствует вообще. Таковы сцеплянки (конъюгаты), большинство зеленых водорослей, имеющих сифоновое строение, харовые, фукусовые и диатомовые водоросли.
Половое размножение, в процессе которого происходит слияние двух голых (лишенных жесткой клеточной стенки) гамет, — широко распространено во всех отделах водорослей. При наиболее примитивной форме полового процесса — изогамии — сливаются гаметы, морфологически не различимые, у большинства водорослей они подвижные, активно передвигающиеся в воде.
Ho иногда неподвижные гаметы могут приводиться в контакт движением родительских талломов. Такой половой процесс называется конъюгацией. Например, у нитчатых водорослей класса Конъюгаты (Сцеплянки) две нити располагаются параллельно, их клетки развивают навстречу друг другу выступы — конъюгационные отростки. Эти отростки затем приходят в соприкосновение, их оболочки растворяются, и образуется канал, по которому содержимое одной клетки перетекает в другую, где оба протопласта сливаются с образованием зиготы. Конъюгация наблюдается и у одноклеточных водорослей-сцеплянок (рис. 17).
При анизогамии (гетерогамии) сливающиеся гаметы различаются своими размерами. Оогамный половой процесс заключается в слиянии крупной неподвижной, лишенной жгутиков яйцеклетки с мелкими, снабженными жгутиками сперматозоидами. Иногда яйцеклетка оплодотворяется мужскими гаметами, лишенными жгутиков. Такие гаметы получили название спермациев, они характерны для представителей отдела Красные водоросли.
Возникающая в результате полового процесса зигота прорастает либо непосредственно после образования, либо после более или менее продолжительного периода покоя. В одних случаях прорастание зиготы сопровождается редукционным делением, в результате чего развиваются гаплоидные растения. Например, у некоторых водорослей отдела Зеленые зигота является единственной диплоидной стадией развития, а вся их вегетативная жизнь проходит в гаплоидном состоянии. Эти водоросли-гаплонты.
У большинства других водорослей, напротив, вся вегетативная жизнь проходит в диплоидном состоянии, а гаплоидная фаза представлена лишь гаметами, перед образованием которых происходит редукционное деление. Эти водоросли (большинство видов отдела Зеленые, имеющие сифоновое строение, все диатомовые, порядок фукусовые из отдела Бурые водоросли), следовательно, являются диплонтами.
Наконец, у ряда водорослей (из отделов Бурые и Красные водоросли, отдела Зеленые водоросли-кладофора, морские виды ульвы) наблюдается правильное чередование гаплоидной и диплоидной фаз (генераций, или поколений). Эти фазы, как мы помним, называют спорофитом и гаметофитом. Причем спорофит и гаметофит могут различаться морфологически (гетероморфная смена генераций), а могут не иметь внешних различий (изоморфная смена генераций).
Мы сознательно так подробно остановились на особенностях полового размножения низших растений для того, чтобы читатель смог проследить описанные тенденции у высших растений. Как же протекает половое размножение у них?
Все современные споровые растения размножаются половым способом. Как мы помним, оплодотворение яйцеклеток, которые формируются в архегониях, осуществляют подвижные сперматозоиды, выплывающие из антеридиев.
Моховидные — это растения, в жизненном цикле которых доминирует гаметофит. Гаметангии развиваются на вершинах побегов (кукушкин лен обыкновенный) или на боковых веточках (мох сфагнум). Двужгутиковые сперматозоиды обеспечивают оплодотворение яйцеклеток только в дождливую погоду, при обильных росах или затоплении участка водой.
Плауновидные имеют явно доминирующий спорофит. А гаметофиты этих растений либо обоеполые и ведут подземный сапротрофный образ жизни (виды рода Ликоподиум), либо они раздельнополые и сильно редуцированы (рис. 18). Оплодотворение 2-3 — жгутиковыми сперматозоидами происходит только при наличии капельной влаги. Поэтому распространение плаунов ограничено влажными тропическими лесами, а в зонах с умеренным климатом — заболоченными участками леса, берегами озер и т. п.
Хвощевидные — это споровые растения, в жизненном цикле которых доминирует спорофит. Гаметофиты хотя и многоклеточные, но сильно редуцированы (мужские варьируют от 1 до 10 мм в диаметре, женские — от 3 до 30 мм), иногда развиваются обоеполые гаметангии (с антеридиями и архегониями), (рис. 19). Сперматозоиды у хвощей имеют многочисленные жгутики (около сотни), они осуществляют оплодотворение яйцеклеток только при наличии капельной воды. Хвощи встречаются в разных растительных зонах, но всегда вблизи воды или в местах с достаточным содержанием влаги в почве.
Папоротниковидные обладают мощно развитым спорофитом, тогда как их гаметофит сильно редуцирован. Как и у плаунов, он может либо вести подземный сапротрофный образ жизни, либо представлять собой миниатюрное зеленое растеньице (заросток), (рис. 20). Например, гаметофит щитовника мужского имеет вид зеленой сердцевидной пластинки площадью около 1 см2, на нижней стороны которой образуются ризоиды, антеридии, а позднее архегонии. Сперматозоиды несут пучок жгутиков, с помощью которых во время дождя или обильной росы активно передвигаются, проникают в архегоний и оплодотворяют яйцеклетку.
Так осуществляется половой процесс у современных споровых растений. К сожалению, гаметофиты растений, обитавших на Земле в минувшие геологические эпохи, сохранились в ископаемом состоянии несравнимо хуже, чем спорофиты. Поэтому их реконструкция весьма затруднена, и мы можем лишь строить догадки о том, как же протекала эволюция полового размножения у растений после их выхода из Мирового океана на сушу. Можно предполагать, что с появлением растений на суше в конце силурийского периода и на протяжении девона и карбона гаметофиты споровых растений «чувствовали себя» достаточно комфортно.
Как предполагают ученые-палеоботаники, споровые растения достигли пышного расцвета в каменоугольном периоде (карбоне), 360-286 млн. лет назад. В это время на Земле преобладали мелководья и болотистые низменности. Климат Евразии и Северной Америки был тропическим и субтропическим. В обширных заболоченных лесах, под пологом гигантских спорофитов папоротников, древовидных хвощей и плаунов — сиггилярий было влажно и сумеречно. С появлением богатейшей растительности каменоугольного периода связано образование перегноя, давшего вместе с песком и глиной начало плодородным влагоемким почвам. Ну, чем не рай для гаметофитов!
В этих условиях гаметофиты споровых растений, унаследовавшие от водорослей подвижные гаметы (сперматозоиды), по-видимому, не испытывали затруднений с осуществлением полового процесса. Действительно, капельной влаги было вдоволь, а в полумраке напочвенного яруса тропического леса их нежные ткани не страдали от перегрева и обезвоживания. Оплодотворенные яйцеклетки древних споровых растений также находили благоприятные условия для своего развития на влажных и богатых перегноем почвах каменоугольного периода.
Однако в конце палеозойской — в начале мезозойской эры климат Земли стал более сухим. Это привело к резкому сокращению представителей флоры, существование которых было сопряжено с условиями высокой влажности. Поэтому с начала пермского периода началось вымирание большинства древесных споровых растений.
Семенные растения, которые появились на Земле еще в конце девонского периода палеозойской эры, в мезозое стали занимать господствующее положение. Какие же адаптации позволили семенным растениям стать доминирующей группой в растительном мире Земли?
Прежде всего, заметим, что на фоне стремительных изменений климата Земли, которые происходили в начале мезозойской эры, прогресса смогли достичь лишь те группы растений, чья наследственная изменчивость протекала столь же интенсивно. В свою очередь наследственная изменчивость организма во многом зависит от того, насколько эффективно осуществляется его половое размножение.
Выше мы рассмотрели особенности полового размножения у споровых растений и убедились, что его эффективность существенно ограничена слабым развитием гаметофита, ведущим к тому же гигрофитный образ жизни. Зависимость гаметофита споровых растений от жидкой среды, как мы видели, обусловлена, с одной стороны, низким уровнем общей организации, а с другой стороны, продуцированием подвижных мужских гамет сперматозоидов, распространение которых возможно только в воде.
Поэтому на фоне уменьшения влажности климата Земли, дальнейший прогресс в царстве растений потребовал от них кардинального совершенствования полового процесса, в частности его «отрыва» от внешней капельной влаги.
Ну, а поскольку гаметофиты семенных растений оказались (благодаря спорофиту) высоко над землей — в воздушной среде, то перед ними встала принципиально новая задача. Предстояло «доверить» многообещающую встречу полов не водной стихии, а воздуху атмосферы.
И вот тут очень кстати оказались миниатюрные размеры мужского гаметофита. Как мы помним, у семенных растений он уменьшился буквально до нескольких клеток (пыльцевого зерна, или пылинки). Благодаря своей микроскопичности, мужской гаметофит приобрел летучесть способность переноситься воздушным потоком на женский спорангий, производить здесь гаметы и оплодотворять яйцеклетку. Миниатюрность пыльцевых зерен помогла растениям «заключить союз» с их переносчиками, в первую очередь с летающими насекомыми, для которых этот груз не стал обременительным.
Невольно напрашивается сравнение парящего в небесах мужского гаметофита (пылинки) с Амуром, несущим на своих крыльях радость встречи полов. Попутно заметим, что «готовясь к свиданию», мужской гаметофит сумел приобрести неплохую защиту: подобно средневековому рыцарю он оделся в «доспехи» — спородерму, вместе с которой обрел стойкость к внешним воздействиям.
Как мы помним, спородерма первоначально защищала бесполые споры растений. Ту же роль она играет теперь и в жизни мужского гаметофита: с первого клеточного деления он развивается под надежной защитой этой прочной оболочки.
В воздушно-сухом состоянии пыльца у многих растений может сохранять жизнедеятельность в течение длительного времени. У тюльпана она сохраняется 38-108 дней, у вишни — 30-100 дней, у груши — 70-210 дней, у сливы — до 180-220 дней, а у финиковой пальмы — до 10 лет. Гораздо легче пыльца переносит низкие температуры (до -20 °С), чем высокие, которые действуют губительно на ее дальнейшее развитие.
Пыльца растений, опыляемых ветром, может распространяться на очень большие расстояния. Специальными опытами установлено, что пыльца березы может переноситься на расстояние до 30-35 км. Пыльца финиковой пальмы переносится воздушными течениями на расстояние до 60 км, а пыльца ольхи и орешника до 400 км. Ho все же наиболее эффективное распространение пыльцы ветром осуществляется на небольшие расстояния между растениями, удаленными друг от друга на несколько десятков или сотен метров.
He всегда пыльца растений сохраняет способность прорастать. Дело в том, что у большинства растений пыльца очень гигроскопична. Она легко впитывает влагу воздуха, набухает, лопается и теряет способность прорастать. Полет пыльцы в сильной степени задерживается лесными массивами, горными вершинами, водоемами. Поэтому огромное количество пылинок погибает, не достигнув цели своего путешествия.
Вот почему одним из приспособлений растений к жизни на суше является образование очень большого количества пыльцы. Например, у ржи на 1 семязачаток (семяпочку) приходится 52 310 пыльцевых зерен, у липы — 43 500, у груши — 60 778. А вот продуктивность одного растения саговника рода энцефаляртос оценивается в 7 млрд. пыльцевых зерен.
Таким образом, у семенных растений половой процесс «оторвался» от водной среды. Благодаря этому революционному событию, растениям удалось успешно противостоять климатическим изменениям в начале мезозойской эры. В значительной степени этому помогло совершенствование механизма оплодотворения и появление качественно нового органа размножения и расселения — семени.
Источник: agro-portal.su