Как осуществляется фагоцитоз
ФАГОЦИТОЗ (phagocytosis, греческий phagos пожирающий -j- kytos вместилище, здесь — клетка + -osis) — процесс узнавания, активного захвата и поглощения микроорганизмов, разрушенных клеток и инородных частиц специализированными клетками иммунной системы.
Объектом фагоцитоза являются микробы, чужеродные и измененные собственные клетки или их фрагменты, комплексы антиген — антитело и др. Неотъемлемую часть фагоцитоза составляет направленное движение — хемотаксис (см. Таксисы) — фагоцитов к месту локализации чужеродной частицы.
Определение эффективности фагоцитоза проводится для оценки состояния иммунобиологической реактивности организма, а также при различных медико-биологических исследованиях.
Явление фагоцитоза как биологической универсальной реакции одноклеточных, многоклеточных и высших организмов было открыто И. И. Мечниковым, который в 1883 году сформулировал теорию фагоцитоза. И. И. Мечников рассматривал фагоцитоз как одну из форм питания клеток (начиная с простейших).
высокоорганизованных организмов эта форма питания свойственна особым мезенхимальным клеткам-фагоцитам поглощающим и убивающим патогенные микробы и таким образом выполняющим защитную функцию. Именно с функцией этих клеток И. И. Мечников связывал иммунитет к возбудителям инфекционных болезней. Им были описаны фазы фагоцитарного процесса и состояние активации фагоцитов, характеризующееся их новыми свойствами и усиленной способностью поглощать и уничтожать бактерии. Ключевая роль фагоцитов была доказана им в иммунитете, при воспалении, удалении поврежденных клеток, регенерации, атрофии, старении.
К фагоцитам относятся гранулоциты, в основном нейтрофильные лейкоциты (см.), и мононуклеарные фагоцитирующие клетки (см. Система мононуклеарных фагоцитов), например, моноциты, макрофаги и др. В процессе узнавания фагоцитами микробов, веществ и частиц большую роль играют особые компоненты сыворотки крови, которые являются молекулярными посредниками при взаимодействии микробов с фагоцитами и обусловливают усиление фагоцитоза. Эти компоненты называются опсонинами (см.), к ним относятся антитела IgGl, IgG3, IgM, агрегированные IgAl и IgA2 (см. Иммуноглобулины), и термолабильные субкомпоненты комплемента, в основном СЗЬ (см. Комплемент), а также а-1 и р-глобулины, сывороточный а2— HS-гликопротеид. Указывают на опсонизирующие свойства С-реактивного белка (см.) и др. Антитела IgG и IgM специфически связываются с антигенами соответствующих бактерий и через Fc-рецепторы фиксируют их к рецепторам фагоцитов. Фагоциты могут соединяться с объектом фагоцитоза и неспецифически — через гидрофобные связи Ван-дер-Ваальса. Субкомпоненты комплемента, возникающие при классическом или альтернативном пути его активации, сорбируются на объектах фагоцитоза , прикрепление которых к поверхности фагоцита осуществляется через СЗЬ-и С4Ь-рецепторы.
Опсонизированные и неопсонизированные частицы прикрепляются к фагоцитам также с помощью специфических Fc-рецепторов для IgE, гликопротеидов и полисахаридов и неспецифических рецепторов для чужеродных веществ. Большинство нейтрофилов человека содержат Fc-рецепторы для агрегированного IgGl и IgG3, а возможно и для агрегированного I g А; моноциты — рецепторы для IgGl и IgG3. Рецепторы для комплемента высокоаффинны (обладают высокой прочностью соединения), они обеспечивают прилипание опсонизированных частиц к неактивированным макрофагам, поглощают же такие частицы только активированные клетки. На нейтрофилах найдены рецепторы для СЗЬ-, С4Ь- и С5а-субкомпонентов комплемента, на макрофагах — один рецептор для СЗЬ- и С4Ь-, другой — для СЗЬ- и СЗс1-субкомпонентов комплемента. Если частица опсонизирована иммуноглобулином и комплементом, связывание с фагоцитом осуществляется кооперативно через специфические к ним рецепторы, что значительно активирует ее поглощение. Имеются различия между классами рецепторов и опосредуемыми ими реакциями фагоцитоза. Посредством неспецифических и специфических для гликопротеидов и полисахаридов рецепторов осуществляется фагоцитоз бактерий без опсонинов. Известен фагоцитоз инертных частиц — кремнезема, угля и др.
Опсонины не только прикрепляют объект фагоцитоза к поверхности фагоцитов, но и активируют их, индуцируя сигналы, идущие от плазматической мембраны, опосредованно вызывают активацию разных гуморальных систем организма, усиливая фагоцитоз.
Процесс поглощения опсонизированной частицы начинается с взаимодействия рецепторов фагоцита с опсонинами, локализованными на поверхности частицы. В дальнейшем происходит взаимодействие соседних свободных рецепторов фагоцита с близлежащими свободными опсонинами частицы до тех пор, пока не будут связаны все опсонины, покрывающие частицу на периферии, и она полностью не погрузится в цитоплазму фагоцита вместе с окружающим участком плазматической мембраны, образуя фагосому. Взаимодействие частицы с плазматической мембраной фагоцита посредством образующихся комплексов опсонин-рецептор запускает сложный механизм фагоцитоза, основная роль в котором принадлежит работе сократительных белков. Процесс поглощения начинается с образования псевдоподии — вытягивания участка цитоплазмы фагоцита в направлении частицы. При формировании псевдоподии находящиеся в ней неориентированные актиновые нити (филаменты) становятся параллельными, что сопровождается преходящим изменением вязкости цитоплазмы. Сформулирована гипотеза жесткости (желатинизации) — сокращения цитоплазмы, изменяющего ее состояние и генерирующего механическую силу движения фагоцита, регулируемого ионами кальция.
и желатинизации актиновые нити перекрестно связываются актинсвязывающим белком, превращающим цитоплазму в гель вследствие образования актиновой решетки. Этот процесс подавляется особЫхМ кальцийзависимым актин-регуляторным белком — гельсолином, являющимся физиол. регулятором желатинизации актина. Далее миозин образует перекрестные мостики с актином и гель начинает сокращаться, особенно в присутствии ионов магния, АТФ и кофактора, являющегося киназой, фосфорилирующей тяжелую цепь миозина. В месте контакта плазматической мембраны и частицы возрастает жесткость цитоплазматических структур (желатинизация участка цитоплазмы). Процесс идет непрерывно; постоянно из плазматической мембраны выделяется растворимый актинсвязывающий белок и мембрана движется по направлению к частице. В области прилипания частицы к плазматической мембране возрастает концентрация ионов кальция, которые «растворяют» актиновую решетку, снижают в этом участке жесткость цитоплазмы, и она движется в сторону повышенной жесткости на конце псевдоподии, т. к. нити миозина натягивают актиновые нити в направлении области наибольшей жесткости решетки.
В процессе фагоцитоза у нейтрофилов потребляется энергия, запасенная в виде АТФ, образованной в результате реакции гликолиза (см.). У альвеолярных макрофагов энергия для фагоцитоза в большей степени (возможно, в основном) извлекается из АТФ, образованной в процессе окислительного фосфорилирования (см. Окисление биологическое).
тановлено, что метаболическим показателем в макрофагах является не абсолютное содержание АТФ, а скорость обновления. Количество АТФ в фагоцитирующих макрофагах частично поддерживается путем фосфорилирования АДФ за счет креатинфосфата (см. Креатин), которого в макрофагах в 3—5 раз больше, чем АТФ, и потребление существенно возрастает при фагоцитозе. Креатинфосфат в макрофагах служит, таким образом, важнейшим резервом и поставщиком химической энергии для фагоцитоза.
Фагоцитоз сопровождается метаболическим, или дыхательным, взрывом, проявляющимся повышением потребления кислорода и окисления глюкозы через гексозомонофосфатный шунт (см. Углеводный обмен). При этом образуются основные продукты восстановления кислорода — супероксидный анион и перекись водорода за счет окисления никотин-амидаденин-динуклеотидов и никотинамидаденин-динуклеотидфосфатов с помощью соответствующих НАДН- и НАДФН-оксидаз; накапливающиеся окисленные коферменты вызывают усиление гексозомонофос-фатного шунта за счет их восстановления с помощью глюкозо-6-фосфат-II 6-фосфоглюконат-дегидрогеназ. Фагоциты имеют сложную систему для разрушения перекиси водорода. Эта система защищает компоненты клетки от разрушения и представлена каталазой, миелопероксидазой, глутатион-пероксидазой, восстановленным глутатионом. Дыхательный взрыв сопровождается усилением метаболизма углеводов, липидов, синтеза РНК, повышением уровня циклического гуанозинмонофосфата, снижением синтеза белка и транспорта аминокислот.
После завершения поглощения частицы возникшая фагосома и первичные лизосомы (см.), первичные азурофильные и вторичные специфические гранулы фагоцитов взаимно сближаются и сливаются, образуя фаголизосому. Этот процесс сопровождается исчезновением в фагоцитах изолированных гранул. Из лизосом в фагосому попадает большое количество гидролитических ферментов. Фагоцитоз также связан с секрецией из фагоцитов ряда ферментов — (З-глюкуронидазы, N-ацетил-Р-глюкозаминидазы, кислой и щелочной фосфатазы, катепсина, миелопероксидазы, лактоферрина, плазминогенного активатора. Подобная секреция сопряжена с активацией гексозомоно-фосфатного шунта и длится значительно дольше, чем непосредственно процесс фагоцитоза.
После проникновения бактерий внутрь фагоцитов начинает функционировать сложный микробоцидный механизм, представленный антимикробными системами, как требующими кислорода, так и не зависящими от него. Антимикробная система, требующая кислорода, функционирует в двух вариантах — с участием и без участия миелопероксидазы. Вариант с участием миелопероксидазы высокоактивен в отношении бактерий, грибков, мико-илазм и вирусов. Взаимодействие миелопероксидазы и перекиси водорода сопровождается образованием окислителей, окислением галоидов и галогенизацией, заключающейся в иодировании, хлорировании, бронировании различных бактериальных компонентов, что приводит к гибели бактерий. При описанных реакциях образуются бактерицидные ионы хлора, иода, хлорамины, нитриты, бактерицидные альдегиды, синглетный кислород, которые блокируют многие ферментные системы бактерий. Не зависящий от миелопероксидазы вариант аштшикробной системы фагоцитов вызывает образование токсичных для микробов промежуточных форм восстановленного кислорода — супероксидного аниона, перекиси водорода, гидроксильного радикала и синглетного кислорода. Наиболее активна из них перекись водорода.
К антимикробной системе фагоцитоза, не зависящей от кислорода, относят: лизоцим (см.), расщепляющий пептидогликаны клеточных стенок некоторых грамположительных бактерий до дисахаридов, состоящих из мураминовой кислоты и глюкозамина; лактоферрин, который в ненасыщенной железом форме оказывает микробостатическое действие в фагосомах за счет связывания железа, являющегося ростовым фактором для ряда из них; различные катионные белки. Определенное бактерицидное действие оказывает также формирующееся в фаголизосомах глубокое закисление до pH 6,5—3,75.
Закисление, кроме того, активирует лизосомальные гидролазы первичных лизосом, неактивные при слабощелочном pH.
Микробоцидные системы фагоцитов функционируют в кооперации. Они обладают различной потенцией, но все вместе оказывают взаимоперекрывающее действие, поэтому обладают высокой надежностью и эффективностью даже при дефектах фагоцитоза.
При нарушении хемотаксиса фагоцитоз бактерий подавлен, что способствует развитию и злокачественному течению ряда инфекционных болезней. Вещества, индуцирующие хемотаксис, называются хемоаттрактантами и подразделяются на несколько групп: 1) продукты специфических, в основном иммунологических реакций,— СЗа-, С5а-субкомпоненты комплемента, активированный комплекс G567, СЗ-конвертаза альтернативного пути активации комплемента, лимфокины (см.
диаторы клеточного иммунитета), трансферфактор лимфоцитов, цитофильные антитела; 2) неспецифические эндогенные хемо-аттрактанты — продукты поврежденных клеток, калликреин (см. Кинины), плазминогенный активатор, фибринопептид В, гидролизованные или агрегированные IgG, коллаген, а- и Р-казеин молока, циклический аденозинмонофосфат и др.; 3) экзогенные хемоаттрактанты — фрагменты белка бактерий, содержащие N-формилметионин, пептиды, липиды или липопротеиды, выделяющиеся в процессе жизнедеятельности бактерий в организме.
На поверхности фагоцитов обнаружены специфические рецепторы для хемоаттрактантов — эйкозатетраеновой кислоты, синтетических формил-метионил-пептидов, С5а-субком-понента кохмплемента. По-видимому, число этих рецепторов неодинаково у разных типов фагоцитов, напр, циркулирующие нейтрофилы кролика в 8 раз слабее связывали хемотаксические пептиды, чем перитонеальные нейтрофилы. Доказана реакция сократительной системы клетки на действие хемоаттрактантов. Ее ориентация на градиент хемоаттрактантов обусловлена работой микротрубочек, выполняющих роль цитоскелета клетки,— они поддерживают поляризованную вытянутую на градиент хемоаттрактантов форму клетки. Однако непосредственно движение фагоцита осуществляет система микрофиламентов. Предполагают, что белки крови — альбумин и IgG являются регуляторами локомоторной функции фагоцитов. Активация фагоцитов хемоаттрактантами во многом сопровождается теми же изменениями, которые происходят при фагоцитозе — метаболическим взрывом, секрецией из клеток ферментов и др. Определенная регулирующая роль принадлежит циклическим нуклеотидам: циклический аденозинмонофосфат подавляет, а циклический гуанозинмоно-фосфат стимулирует хемотаксис.
Способы и методические подходы к оценке фагоцитоза разнообразны и зависят от конкретных задач исследования. Они позволяют определить эффективность процессов поглощения частиц, гибели и переваривания живых микроорганизмов и метаболические изменения фагоцитов. Важные данные о фагоцитозе могут быть также получены при исследовании хемотаксиса и опсонизации.
Для оценки фагоцитоза используют различные микроорганизмы — стафилококки (см.), эшерихии (см.), сальмонеллы (см. Сальмонелла) и др. Используют как живые, так и убитые микробы, но поскольку живые бактерии нередко выделяют токсические продукты, подавляющие фагоцитоз, лучше использовать убитые.
Фагоцитоз усиливается в присутствии сыворотки, опсонизирующей бактерии. Для усиления и стандартизации фагоцитоза используют предопсонизацию, то есть предварительную (до фагоцитоза) обработку микроба опсонинами — специфическими антителами — либо свежей сывороткой, в которой микробы активируют систему комплемента и адсорбируют появляющиеся субкомпоненты комплемента, облегчающие фагоцитоз.
нако в экспериментах с живыми микробами применяют лишь те, которые не убиваются опсонизирующей сывороткой. Скорость фагоцитоза анализируют при совместном инкубировании фагоцитов и живых бактерий. Через разные промежутки времени забирают пробы, с помощью дифференциального центрифугирования освобождаются от фагоцитов и надосадочную жидкость сеют на чашки с агаром, что позволяет определить уменьшение числа живых бактерий в процессе фагоцитоза. При работе с грибками рода Candida препарат просчитывают в камере Горяева, определяя при этом число внеклеточно расположенных грибков.
Для анализа фагоцитоза путем определения процента фагоцитов, поглотивших бактерии (фагоцитарный индекс Гамбургера), или среднего числа бактерий, поглощенных одним фагоцитом (фагоцитарное число Райга), скорости фагоцитоза используют частицы латекса, крахмала, зимозана, кармина, угля и др. Предложен метод исследования фагоцитоза, при котором используют капельки парафинового масла, содержащего специальный краситель и стабилизированного белком. Поглощенный материал определяют спектрофотометрически (см. С пектрофотометрия). Также используют частицы или микробы, меченные радиоактивными изотопами (см. Меченые соединения). Метод характеризуется быстротой выполнения, однако не позволяет полностью избавиться от прилипших бактерий, что завышает показатели фагоцитоза. Другой вариант состоит в добавлении к среде с фагоцитами и частицами меченых сывороточных белков, которые при фагоцитозе попадают в фагосому, что позволяет оценить количественно интенсивность фагоцитоза. Применяют также ксеногенные интактные или сингенные поврежденные или опсонизированные эритроциты, анализируя их поглощение визуально или по выходу гемоглобина.
При исследовании поглощения живых бактерий, особенно с последующим учетом количества убитых бактерий необходимо удалить с поверхности фагоцитов прилипшие микробы. Для этого применяют различные антибиотики, убивающие внеклеточные бактерии, но не проникающие в фагоциты, специальные препараты (фенилбутазан), прерывающие в определенные моменты фагоцитоза и внутриклеточную инактивацию микробов. Разработан метод, позволяющий различать прилипшие и поглощенные убитые грибки рода Candida по окраске препарата трипановым синим.
Гибель и переваривание поглощенных микробов выявляют путем инкубирования суспензии фагоцитов с микробами, последующего отмывания фагоцитов of прилипших микробных клеток, подсчета живых микробов, оставшихся в пробах фагоцитов, забираемых в различные сроки инкубации. Число живых бактерий определяют серийными посевами из проб фагоцитов на чашки Петри с агаром. Число живых грибков подсчитывают в лизате фагоцитов после инкубации с помощью окрашивания метиленовым синим. Внутриклеточное переваривание бактерий изучают также с помощью включения в них 3Н-уридина. Для этого культуру фагоцитов, поглотивших бактерии, обрабатывают актиномицином D, добавляя в среду 3Н-уридин. Метка, включаясь в живые внутриклеточные бактерии, не попадает в убитые и фагоциты.
Анализ повреждающего действия фагоцитов на микробы можно проводить по степени окрашивания поглощенных микробов красителями или по окраске метиленовым синим фаголизосом фагоцитов. Завершенность фагоцитоза оценивают по отношению среднего числа убитых микробов к живым или числа фагоцитов с переваренными микробами к общему числу фагоцитирующих фагоцитов, а также по проценту разрушенных микробов от числа фагоцитированных или по среднему числу убитых микробов на один фагоцит. Выраженность метаболических изменений при фагоцитозе анализируют по потреблению кислорода, хемилюминесценции, окислению глюкозы, иодированию и др.
Фагоциты играют ключевую роль в формировании противомикробного иммунитета (см. Иммунитет), обусловленного как специфическими, так и неспецифическими факторами защиты. Несмотря на то, что специфический иммунитет опосредуется специфическими Т-клетками, а также специфическими антителами, опсонизирующими бактерии и усиливающими фагоцитоз, элиминация патогенных бактерий осуществляется неспецифически — фагоцитами, активированными лимфокинами специфических Т-лимфоцитов. Активированные фагоциты значительно эффективнее убивают бактерии, что показал еще И. И. Мечников. Естественная невосприимчивость к возбудителям инфекционных болезней также обусловлена в основном фагоцитарными клетками. Ключевая роль принадлежит им и в детоксикации бактериальных токсинов, нейтрализованных антителами.
Макрофаги, перерабатывая антиген и представляя его лимфоцитам, участвуя в межклеточной кооперации, активации и супрессии пролиферации лимфоцитов, являются необходимым звеном в формировании иммунологической толерантности (см. Толерантность иммунологическая) и трансплантационного иммунитета (см. Иммунитет трансплантационный). Макрофаги участвуют в противоопухолевом иммунитете (см. Иммунитет противоопухолевый), оказывая цитостати-ческое и цитотоксическое действие на опухолевые клетки.
Повреждения фагоцитов различными иммуносупрессорами, бло-каторами (см. Иммунитет, Йммуподепрессивные вещества), ионизирующим излучением (см.) вызывают резкое подавление противомикробной устойчивости организма. При воздействии на животных большими дозами ионизирующего излучения фагоцитарная активность может практически исчезнуть. Нормализуется фагоцитарная активность у животных, как правило, после 20-го дня. У кроликов, облученных в дозе 600 рад (6 Гр), она восстанавливается только через 40 дней. Между дозой ионизирующего излучения и степенью подавления фагоцитоза существует корреляция. Дозы 10—75 рад (0,1 — 0,75 Гр) усиливают фагоцитоз гранулоцитов, а 350—600 рад (3,5—6 Гр)—резко его угнетают, причем снижается завершенность фагоцитоз, в 3—4 раза подавляется подвижность фагоцитов, а также уменьшается абсолютное их число. Эти же закономерности характерны для макрофагов, число и переваривающая способность которых при облучении также резко снижаются.
Выявлены болезни, сопровождающиеся первичными (врожденными) или вторичными (приобретенными) дефектами фагоцитоза. К ним относится так называемая хроническая гранулематозная болезнь, возникающая у детей, в фагоцитах которых из-за дефекта оксидаз нарушено образование перекисей и надперекпсей и, следовательно, процесс инактивации микробов. Сниженная способность к уничтожению бактерий выявлена у людей, нейтрофилы которых синтезируют недостаточное количество миелопероксидазы, глюкозо-б-фосфат-дегидрогеназы, пируваткиназы. Замедленная гибель микробов обнаружена у больных с синдромом Чедиака — Хигаси (см. Тромбоцитопатии), в нейтрофилах которых нарушено выделение в фагосому лизосомальных ферментов из-за дефекта системы микротрубочек. Описано нарушение процесса полимеризации актина, ведущее к замедлению поглощения частиц нейтрофилами и их подвижности. Больные с указанными дефектами фагоцитов часто страдают тяжелыми бактериальными и грибковыми инфекциями.
Первичные нарушения фагоцитоза наблюдаются и на уровне опсонинов, например, при врожденном дефиците СЗ- и С5-компонентов комплемента, который может привести к развитию рецидивирующих инфекций с поражением легких, костей, кожи.
Вторичные дефекты фагоцитоза описаны при заболеваниях соединительной ткани, почек, нарушении питания, вирусных и рецидивирующих бактериальных инфекциях.
Библиогр.: Берман В. М. и Слав-с к а я E. М, Завершенный фагоцитоз, Журн. микр., эпид. и иммун., № 3, с. 8, 1958; П о д о п р и г о р а Г. И. и Андреев В. Н. Современные методы изучения фагоцитарной активности лейкоцитов in vitro, там же, № 1, е. 19, 1976; X р а м-цов А. В. и Земсков В. М. Роль плазматической мембраны в активации лизосомальных ферментов, Докл. АН СССР, т. 271, № 1, с. 241, 1983; Handbook of experimental immunology, ed. by D. M. Weir, v. 2—3, Oxford a. o., 1979; Handbook of experimental pharmacology, ed. by J. R. Vane a. S. H. Ferreira, v. 50, pt 1, В. a. o., 1978; KlebanoffS. J. a. Clark R. A. The neutrophil, function and clinical disorders, Amsterdam a. o., 1978; Mononuclear phagocytes, Functional aspects, ed. by R. van Furth, pt 1 — 2, Hague a. o., 1980; The reticuloendothelial system, a comprehensive treatise, v. 1 — Morphology, ed. by H. Friedman a. o., N. Y.— L., 1980.
В. М. Земсков.
Источник: xn--90aw5c.xn--c1avg
Клеточные факторы системы видовой резистентности- фагоциты, поглощающие и разрушающие патогенные микроорганизмы и другой генетически чужеродный материал. Представлены полиморфоядерными лейкоцитами или гранулоцитами— нейтрофилами, эозинофилами и базофилами (клетками миелопоэтического ряда), а также моноцитами и тканевыми макрофагами (клетками макрофагально- моноцитарной системы).
Значение фагоцитирующих клеток для защиты организма впервые доказал И.И.Мечников, разработавший фагоцитарную теорию иммунитета.
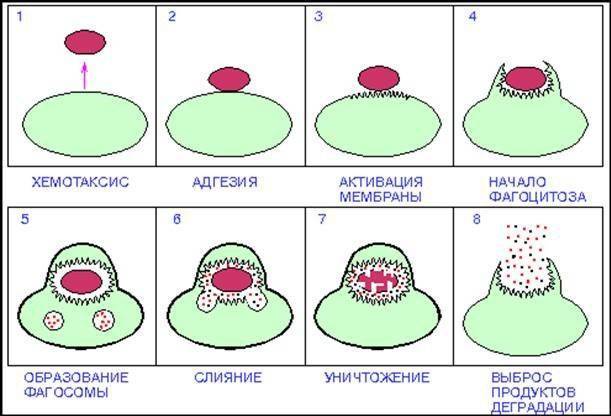
Стадии фагоцитоза.
Процесс фагоцитоза (поглощения твердофазного объекта) состоит из пяти стадий.
1.Активация (усиление энергетического метаболизма). Факторами активации и хемотаксиса являются бактериальные продукды (ЛПС, пептиды), компоненты комплемента (С3 и С5), цитокины и антитела.
2.Хемотаксис.
3.Адгезия.
4.Поглощение.
5. Внутриклеточный киллинг и переваривании (либо исход фагоцитоза)
Адгезия связана с наличием ряда рецепторов на поверхности фагоцитов ( к Fc- фрагментам антител, компонентам комплемента, фибронектину), обеспечивающих прочность рецептор- опосредованных взаимодействий опсонинов, обволакивающих микроорганизмы и ограничивающих их подвижность (антитела, С3в, фибронектин).
Фагоциты обладают амебоподобными псевдоподиями. При поглощении образуется фагосома с поглощенным объектом (бактерией), к ней присоединяется и сливается содержащая литические ферменты лизосома, образуется фаголизосома.
Возможно три исхода фагоцитоза:
— завершенный фагоцитоз;
— незавершенный фагоцитоз;
— процессинг антигенов.
Завершенный фагоцитоз- полное переваривание микроорганизмов в клетке- фагоците.
Незавершенный фагоцитоз- выживание и даже размножение микроорганизмов в фагоците. Это характерно для факультативных и особенно — облигатных внутриклеточных паразитов. Механизмы персистирования в фагоцитах связаны с блокадой фагосомо- лизосомального слияния (вирус гриппа, микобактерии, токсоплазмы), резистентностью к действию лизосомальных ферментов (гонококки, стафилококки), способностью микробов быстро покидать фагосомы после поглощения и длительно пребывать в цитоплазме (риккетсии).
В процессе фагоцитоза происходит “окислительный взрыв” с образованием активных форм кислорода, что обеспечивает бактерицидный эффект.
К одной из важнейших функций макрофагов (наряду с хемотаксисом, фагоцитозом, секрецией биологически активных веществ) является переработка (процессинг) антигена и представление его иммунокомпетентным клеткам с участием белков главной системы гистосовместимости (МНС) класса 2.
Фагоцитоз — не только уничтожение чужеродного, но и представление антигена для запуска иммунных реакций и секреции медиаторов иммунных и воспалительных реакций. Система макрофагов- центральное звено не только естественной резистентности (видового иммунитета), но и играет важную роль в приобретенном иммунитете, кооперации клеток в иммунном ответе.
2.11 Комплемент, его структура, функции, пути активации, роль в иммунитете.
Природа и характеристика комплемента. Комплемент является одним из важных факторов гуморального иммунитета, играющим роль в защите организма от антигенов. Комплемент представляет собой сложный комплекс белков сыворотки крови, находящийся обычно в неактивном состоянии и активирующийся при соединении антигена с антителом или при агрегации антигена. В состав комплемента входят 20 взаимодействующих между собой белков, девять из которых являются основными компонентами комплемента; их обозначают цифрами: С1, С2, СЗ, С4… С9. Важную роль играют также факторы В, D и Р (пропердин). Белки комплемента относятся к глобулинам и отличаются между собой по ряду физико-химических свойств. В частности, они существенно различаются по молекулярной массе, а также имеют сложный субъединичный состав: Cl-Clq, Clr, Cls; СЗ-СЗа, СЗЬ; С5-С5а, С5b и т. д. Компоненты комплемента синтезируются в большом количестве (составляют 5—10% от всех белков крови), часть из них образуют фагоциты.
Функции комплемента многообразны: а) участвует в лизисе микробных и других клеток (цитотоксическое действие); б) обладает хемотаксической активностью; в) принимает участие в анафилаксии; г) участвует в фагоцитозе. Следовательно, комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов(например, опухолевых клеток, трансплантата).
Механизм активации комплемента очень сложен и представляет собой каскад ферментативных протеолитических реакций, в результате которого образуется активный цитолитический комплекс, разрушающий стенку бактерии и других клеток. Известны три пути активации комплемента: классический, альтернативный и лектиновый.
По классическому пути комплемент активируется комплексом антиген-антитело. Для этого достаточно участия в связывании антигена одной молекулы IgM или двух молекул IgG. Процесс начинается с присоединения к комплексу АГ+АТ компонента С1, который распадается на субъединицы Clq, Clr и С Is. Далее в реакции участвуют последовательно активированные «ранние» компоненты комплемента в такой последовательности: С4, С2, СЗ. Эта реакция имеет характер усиливающегося каскада, т. е. когда одна молекула предыдущего компонента активирует несколько молекул последующего. «Ранний» компонент комплемента С3 активирует компонент С5, который обладает свойством прикрепляться к мембране клетки. На компоненте С5 путем последовательного присоединения «поздних» компонентов С6, С7, С8, С9 образуется литический или мембраноатакующий комплекс который нарушает целостность мембраны (образует в ней отверстие), и клетка погибает в результате осмотического лизиса.
Альтернативный путь активации комплемента проходит без участия антител. Этот путь характерен для защиты от грамотрицательных микробов. Каскадная цепная реакция при альтернативном пути начинается с взаимодействия антигена (например, полисахарида) с протеинами В, D и пропердином (Р) с последующей активацией компонента СЗ. Далее реакция идет так же, как и при классическом пути — образуется мембраноатакующий комплекс.
Лектиновыи путь активации комплемента также происходит без участия антител. Он инициируется особым маннозосвязывающим белком сыворотки крови, который после взаимодействия с остатками маннозы на поверхности микробных клеток катализирует С4. Дальнейший каскад реакций сходен с классическим путем.
В процессе активации комплемента образуются продукты протеолиза его компонентов — субъединицы СЗа и СЗb, С5а и С5b и другие, которые обладают высокой биологической активностью. Например, СЗа и С5а принимают участие в анафилактических реакциях, являются хемоаттрактантами, СЗb — играет роль в опсонизации объектов фагоцитоза, и т. д. Сложная каскадная реакция комплемента происходит с участием ионов Са2+ и Mg2+.
Источник: megaobuchalka.ru
Этапы фагоцитоза:
Хемотаксис
Вредоносный объект проникает в организм, и он недолго остается там незамеченным. Этот объект, будь то бактерия, инородное тело или что-то еще, выделяет особые вещества (хемоаттрактанты) и прямо контактирует с кровью или тканями. Все это ставит организм в известность о присутствии внутри него агрессора.
Возникает каскад биохимических реакций. На первой  стадии фагоцитоза тучные клетки выбрасывают в кровь специальные соединения, вызывающие реакцию воспаления. Начало воспалительного процесса «пробуждает» от состояния покоя макрофаги и другие клетки-фагоциты. Нейтрофилы, уловив присутствие хемоаттрактантов, быстро выходят из крови в ткани и спешат мигрировать к воспалительному очагу.
стадии фагоцитоза тучные клетки выбрасывают в кровь специальные соединения, вызывающие реакцию воспаления. Начало воспалительного процесса «пробуждает» от состояния покоя макрофаги и другие клетки-фагоциты. Нейтрофилы, уловив присутствие хемоаттрактантов, быстро выходят из крови в ткани и спешат мигрировать к воспалительному очагу.
Сложно это описать, а еще сложнее себе это представить, но проникновение патогена в организм ведет к запуску настоящего эффекта домино, включающего сотни (!) различных физиологических явлений, проходящих на клеточном и субклеточном уровнях. Состояние иммунной системы на этом этапе фагоцитоза можно сравнить с состоянием потревоженного пчелиного улья, когда его многочисленные обитатели готовятся атаковать обидчика.
Адгезия
Последовательность фагоцитоза продолжается второй стадией – реакцией адгезии. Подошедшие к нужному месту фагоциты протягивают к патогену свои отростки, вступают с ним в контакт и распознают его. Они не спешат сразу нападать и вначале предпочитают убедиться, не ошибаются ли они на счет «чужака». Распознание вредоносного агента происходит при помощи особых рецепторов на поверхности мембран фагоцитов.
Активация мембраны
На третьей стадии фагоцитоза в клетках-защитниках происходят невидимые реакции, которые подготавливают их к захвату и уничтожению патогена.
Погружение
Мембрана фагоцита – это текучая, пластичная субстанция, которая может менять форму. Что она и делает, когда клетка сталкивается с вредоносным объектом. На фото видно, что фагоцит протягивает к чужеродной частице свои «щупальца». Потом он постепенно растекается вокруг нее, наползает на нее и полностью ее захватывает.
Образование фагосомы
Когда фагоцит охватывает частицу со всех сторон, его мембрана замыкается снаружи, а внутри клетки остается закрытый пузырек с атакованным объектом внутри. Таким образом, клетка как будто проглатывает частицу. Этот пузырек носит название фагосомы.
Формирование фаголизосомы (слияние)
Пока проходили другие этапы фагоцитоза, внутри фагоцита готовилось к использованию его оружие – органеллы-лизосомы, содержащие «пищеварительные» ферменты клетки. Как только бактерия или другой вредный объект оказался пленен клеткой-защитником, к ней приближаются лизосомы. Их мембраны сливаются с оболочкой, обволакивающей частицу, и их содержимое изливается внутрь этого «мешка».
Киллинг
Это самый драматичный момент во всем механизме фагоцитоза. Захваченный объект переваривается и расщепляется фагоцитом.
Удаление продуктов расщепления
Все, что осталось от убитой бактерии или другой переваренной частицы, удаляется из клетки. Бывшая фаголизосома, представляющая собой мешочек с продуктами деградации, подходит к наружной мембране фагоцита и сливается с ней. Так из клетки удаляются остатки поглощенного объекта. Последовательность фагоцитоза завершается.
Источник: www.transferfaktory.ru