Полное доминирование фенилкетонурия
Вприроде наряду с полным доминированием часто встречается неполное (промежуточное), когда гетерозиготы имеют иной фенотип, чем исходные родительские формы, выражение признака у гетерозиготы оказывается промежуточным. Неполное доминирование заключается в ослаблении действия доминантного аллеля в присутствии рецессивного.
Например, у человека анофтальмия (отсутствие глазных яблок) обусловлена аутосомно-рецессивным геном, нормальная величина глазных яблок — доминантным геном. Гетерозиготы имеют микрофтальмию, т.е. уменьшенную величину глазных яблок. У гомозигот по рецессивным аллелям глазные яблоки отсутствуют, у гетерозигот обнаруживается микрофтальмия, у гомозигот по доминантным аллелям яблоки имеют нормальную величину. Задача3
На определение генотипа и фенотипа потомства детей по фенотипу и генотипу родителей при неполном (промежуточном) доминировании
Здоровая женщина с нормальной величиной глазных яблок выходит замуж за мужчину, страдающего анофтальмией (отсутствие глазных яблок). Каков прогноз потомства, если дети от указанной пары вступят в брак с себе подобными (избранные браки).
Решение:
Обозначим гены, определяющие развитие глазных яблок: А— нормальная величина глазных яблок,а — анофтальмия, Аа — микрофтальмия
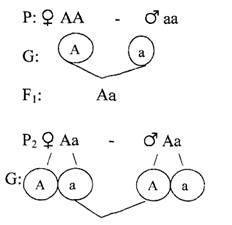
F2: АА : 2Аа : аа
1 : 2 : 1 три фенотипических класса, расщепление по фенотипу.
1 : 2 : 1 три генотипических класса, расщепление по генотипу. Ответ:в первом поколении (F1), когда родители гомозиготны (анофтальмия проявляется только у гомозигот по рецессивным аллелям), все дети имеют уменьшенную величину яблок (микрофтальмия); генотипически они все гетерозиготны, т.е. проявился I закон (правило) единообразия. Г. Менделя, однако, фенотипически дети не имеют сходства с родителями.
Во втором поколении (F2) при неполном доминировании расщепление по фенотипу и генотипу совпадает 1:2:1,т.е. 1/4(25%) детей с нормальной величиной глазных яблок; 1/2(50%) с микрофтальмией 1/4(25%) детей с отсутствием глазных яблок (анофтальмией).
У человека наследуются неполным доминированием признак волнистых волос, заболевания: акаталазия, микрофтальмия, фенилкетонурия, серповидно-клеточная анемия и др. Отклонения от ожидаемого расщепления, связанные с летальными генами
В природе в некоторых случаях во втором поколении расщепление может отличаться от ожидаемого, в связи с тем что гомозиготы по некоторым генам оказываются нежизнеспособными. У человека аналогично наследуется доминантный ген брахидактилии (укороченные фаланги пальцев и редуцированное число самих фаланг). В браках, когда муж и жена страдают брахидактилей, рождаются дети в отношении: две части короткопалых и одна часть с нормальными пальцами (2:1, а не 3:1), доминантные гомозиготные особи погибают еще в эмбриональном развитии.
Задача 4
На определение фенотипа и генотипа потомства по фенотипу и генотипу
родителей при отклонении от ожидаемого расщепления
Одна из пород кур отличается укороченными ногами. Признак этот — доминантный. Контролирующий его ген вызывает одновременно также и укорочение клюва. При этом у гомозиготных цыплят клюв так мал, что они не в состоянии пробить яичную скорлупу и гибнут, не вылупившись из яйца. В инкубаторе хозяйства, разводившего только коротконогих кур, получено 4500 цыплят. Сколько среди них коротконогих?
Решение:
Обозначим гены, определяющие развитие ног и клюва.
А — укороченный клюв и укороченные ноги
а— нормальный клюв,
АА — укороченный клюв, укороченные ноги, летальные гомозиготы.
Поскольку в инкубаторе разводили только коротконогих кур, следовательно все куры в данном хозяйстве по генотипу — гетерозиготны (гомозиготы, как было отмечено, гибнут, не вылупившись).
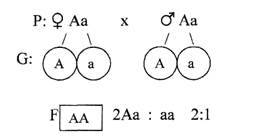
Наблюдается расщепление 2:1, т.е. коротконогие составляют две трети
выводка — в данном случае

Ответ: количество коротконогих с укороченным клювом 3000 цыплят.
Явление, когда один ген определяет, т.е. детерминирует, несколько признаков, причем один из них в большей степени, чем другие, носит название плейотропии.
Источник: helpiks.org
Доминирование генов
Вы когда-нибудь задумывались, почему у вас есть такой особый цвет глаз или тип волос? Все это связано с передачей генов. Как обнаружил Грегор Мендель, черты наследуются при помощи передачи генов от родителей к их потомкам. Гены представляют собой участки ДНК, расположенные на наших хромосомах. Они передаются от одного поколения к другому через половое размножение. Ген для конкретного признака может существовать в более чем одной форме или аллеле. Для каждой характеристики или признака клетки животных обычно наследуют два аллеля. Парные аллели могут быть гомозиготными (с идентичными аллелями) или гетерозиготными (с разными аллелями) для данного признака.
Когда пары аллелей одинаковы, генотип этого признака идентичен, а фенотип или характеристика, которая наблюдается, определяется гомозиготными аллелями. Когда парные аллели для признака различные или гетерозиготные, возможно несколько вариантов. Гетерозиготные аллели, которые обычно наблюдаются в клетках животных, включают полное доминирование, неполное доминирование и кодоминирование.
Полное доминирование
» data-layzr=»https://natworld.info/wp-content/uploads/2017/07/горох.jpg» alt=»» width=»500″ height=»333″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/07/горох.jpg 500w, https://natworld.info/wp-content/uploads/2017/07/горох-300×200.jpg 300w» sizes=»(max-width: 500px) 100vw, 500px» />
При этом виде доминирования один аллель является доминирующим, а другой рецессивным. Доминантный аллель полностью маскирует рецессивный. Фенотип определяется доминирующим аллелем. Например, гены для формы семени в растениях гороха существуют в двух формах: гладкая (R) и морщинистая (r). В растениях гороха, которые являются гетерозиготными по форме семени, гладкий горох является доминирующим по отношению морщинистому семени, а генотип является (Rr).
Неполное доминирование
» data-layzr=»https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы.jpg» alt=»» width=»500″ height=»332″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы.jpg 500w, https://natworld.info/wp-content/uploads/2017/07/волнистые-волосы-300×199.jpg 300w» sizes=»(max-width: 500px) 100vw, 500px» />
При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов. Пример неполного доминирования проявляется в наследовании типа волос. Кудрявый тип волос (CC) является доминирующим для прямого типа волос (сс). Человек, гетерозиготный по этому признаку, будет иметь волнистые волосы (Cc).
Доминирующая фигурная характеристика не полностью выражена по прямой характеристике, создавая промежуточную характеристику волнистых волос. При неполном доминировании одна характеристика может быть несколько более заметной, чем другая для данного признака. Например, у человека с волнистыми волосами может быть больше или меньше волн, чем у другого с волнистыми волосами. Это указывает на то, что аллель для одного фенотипа выражается немного больше, чем аллель для другого фенотипа.
Кодоминирование
При совместном доминировании ни один из аллелей не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа. Пример кодоминирования наблюдается у индивидов с чертой серповидной клетки.
Наличие серповидной клетки связано с развитием эритроцитов с аномальной формой. Нормальные эритроциты имеют двояковогнутую, дискообразную форму и содержат огромное количество белка, называемого гемоглобином. Гемоглобин помогает красным клеткам крови связываться и переносить кислород в клетки и ткани организма. Серповидная клетка является результатом мутации гена гемоглобина. Этот гемоглобин считается ненормальным и заставляет клетки крови принимать серповидную форму.
Серповидные клетки часто застревают в кровеносных сосудах и блокируют нормальный кровоток. Те, которые несут черту серповидной клетки, гетерозиготные для гена гемоглобина и наследуют один нормальный ген гемоглобина и один серповидный ген гемоглобина. У них нет болезни, потому что аллель гемоглобина серпа и нормальный гемоглобиновый аллель являются кодоминантными по отношению к форме клеток. Это означает, что у носителей серповидных клеток образуются как нормальные эритроциты, так и серповидные.
Неполное доминирование и кодоминирование
Люди часто путают неполное доминирование и кодоминирование. Хотя они и являются примерами наследования, но отличаются выражением генов. Ниже перечислены некоторые различия между ними:
Экспресивность аллеля
- Неполное доминирование: один аллель для определенного признака не полностью выражен над его парным аллелем. Например, цвет тюльпана, аллель для красного цвета (R) не полностью маскирует аллель для белого цвета (r).
- Кодоминирование: оба аллеля для конкретного признака полностью выражены. Аллель для красного цвета (R) и аллель для белого цвета (r) выражены и видны в гибриде.
Зависимость аллелей
- Неполное доминирование: влияние одного аллеля зависит от его парного аллеля для данного признака.
- Кодоминирование: влияние одного аллеля не зависит от его парного аллеля для данного признака.
Фенотип
- Неполное доминирование: Гибридный фенотип представляет собой смесь обеих аллелей, что приводит к третьему промежуточному фенотипу. Пример: Красный цветок (RR) X Белый цветок (rr) = Розовый цветок (Rr)
- Кодоминирование: гибридный фенотип представляет собой комбинацию выраженных аллелей, что приводит к третьему фенотипу, который включает оба фенотипа. Пример: Красный цветок (RR) X Белый цветок (rr) = Красно-белый цветок (Rr)
Наблюдаемые характеристики
- Неполное доминирование: фенотип может быть выражен в разной степени в гибриде. (Пример: розовый цветок может иметь более легкую или более темную окраску в зависимости от количественной выраженности одного аллеля по сравнению с другим).
- Кодоминирование: оба фенотипа полностью выражены в гибридном генотипе.
Краткий вывод
При неполном доминировании один аллель для конкретного признака не является полностью доминирующим над другим аллелем. Это приводит к третьему фенотипу, в котором наблюдаемые характеристики представляют собой смесь доминирующих и рецессивных фенотипов.
При кодоминировании ни один аллель не является доминирующим, но оба аллеля для конкретного признака полностью выражены. Это приводит к третьему фенотипу, в котором наблюдается более одного фенотипа.
Понравилась статья? Поделись с друзьями:
Источник: NatWorld.info
Полное доминирование
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы по данному аллелю. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный аллель), имеют половинное количество нормального фактора свертывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминирование
При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный — дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную — нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят к промежуточному характеру наследования, то есть отсутствию доминирования[1].
Кодоминирование
При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно и в полной мере. Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвёртая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов — это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Другие варианты доминирования
- Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. На этом типе аллельного взаимодействия основано явление гетерозиса (превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности)[1].
- Доминирование, связанное с полом, происходит тогда, когда одна и та же аллель у самцов проявляется как доминантная, а у самок — как рецессивная. Например, у овцематок доминирует комолость (R), а у баранов — рогатость (R1)[1].
Относительный характер доминирования
Как уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии. Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2,5-3 тыс. м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в равных количествах присутствуют оба варианта бета-глобиновых цепей — нормальный А и мутантный S (то есть наблюдается кодоминирование).
Молекулярные механизмы
Молекулярные основы доминирования были неизвестны Менделю. В настоящее время ясно, что локус, соответствующий определённому гену, состоит из длинных последовательностей, включающих сотни и тысячи нуклеотидов ДНК. Центральная догма молекулярной биологии гласит, что ДНК → РНК → белок, то есть ДНК транскрибируется в мРНК, а мРНК транслируется в белок. В этом процессе различные аллели могут транскрибироваться или не транскрибироваться, а будучи затранскрибированными, транслироваться в различные формы одного и того же белка — изоформы. Часто белки функционируют как ферменты, катализирующие химические реакции в клетке, которые прямо или косвенно определяют фенотип. У любого диплоидного организма аллели, соответствующие одному локусу, являются либо одинаковыми (у гомозигот), либо разными (у гетерозигот). Даже если на уровне последовательностей ДНК аллели различны, то их белки могут быть идентичными. В отсутствие различий между белковыми продуктами невозможно сказать, какой из аллелей доминирует (в этом случае имеет место кодоминирование). Даже если два белковых продукта слегка отличны друг от друга, они, вероятно, дают одинаковый фенотип и могут осуществлять одинаковые ферментативные реакции (если они являются ферментами). В этом случае также невозможно сказать, какой из аллелей доминирует.
Доминирование, как правило, возникает, когда один из аллелей является нефункциональным на молекулярном уровне, то есть не транскрибируется или даёт нефункциональный белковый продукт. Это может быть результатом мутации, изменяющей последовательность ДНК аллеля. У гомозиготы по нефункциональным аллелям, как правило, проявляется характерный фенотип из-за отсутствия определённого белка. Например, у людей и других животных непигментированная кожа альбиносов проявляются из-за гомозиготности по аллелю, препятствующему синтезу кожного пигмента меланина. Важно понимать, что рецессивность определяется у аллеля не по отсутствию какой-либо функции: у гетерозигот это является результатом взаимодействия с альтернативным аллелем. Возможны три основных типа таких взаимодействий:
- В типичном случае единичный функциональный аллель дает достаточно белка, чтобы получить фенотип, идентичный фенотипу гомозиготы по функциональному аллелю. Это называется гаплодостаточностью (англ. haplosufficiency). Например, если принять количество фермента, производимого функциональной гетерозиготой, за 100%, то каждый из функциональных аллелей будет ответственна за выработку 50% общего количества фермента. Единственный функциональный аллель гетерозиготы даёт 50% фермента, и этого достаточно для поддержания нормального фенотипа. Если гетерозигота и гомозигота по функциональному аллелю имеют одинаковый фенотип, то функциональный аллель доминирует над нефункциональным. Так происходит с геном альбинизма: гетерозигота производит количество фермента, которого достаточно для образования предшественника меланина, и особь имеет нормальную пигментацию.
- Реже наличие единственного функционального аллеля не обеспечивает нормальный фенотип, однако его дефектность выражена не так ярко, как у гомозиготы по нефункциональным аллелям. Это происходит тогда, когда функциональный аллель не является гаплодостаточным. Обычно к этим случаям относят понятия гаплонедостаточности и неполного доминирования. Промежуточный вариант этого взаимодействия имеет место тогда, когда гетерозигота имеет фенотип, промежуточный между двумя гомозиготами. В зависимости от того, к какой из гомозигот ближе вариант признака гетерозиготы, говорят о неполном доминировании одной аллели над другой. Примером такого взаимодействия может служить описанный выше случай с гемоглобином человека.
- Редко единственная функциональная аллель гетерозиготы даёт неполноценный генный продукт, и её фенотип схож с фенотипом гомозиготы по нефункциональным аллелям. Такие случаи гаплонедостаточности крайне необычны. В этих случаях нефункциональная аллель доминирует над функциональной. Такая ситуация может происходить тогда, когда нефункциональная аллель даёт дефектный белок, который подавляет функцию белка, образуемого нормальной аллелью. Дефектный белок «доминирует» над стандартным, и фенотип гетерозиготы более походит на фенотип гомозиготы по дефектным аллелям. Следует обратить внимание на то, что доминантными часто некорректно называют дефектные аллели, вызываемый которыми в гомозиготном состоянии фенотип не изучен, однако в сочетании с нормальным аллелем они дают характерный фенотип. Этот феномен происходит при некоторых генетических заболеваниях, вызванных тринуклеотидными повторами, например, болезни Хантингтона.
Эволюция доминантности
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путём отбора малых мутаций как структурных генов, так и генов-модификаторов.
Так или иначе, степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое, по некоторым данным, произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название «индустриальный меланизм») (Н. Н. Иорданский «Эволюция жизни»)
См. также
- Аллель
- Гетерозигота
- Гомозигота
- Доминантный признак
- Рецессивный признак
- Законы Менделя
- Взаимодействие неаллельных генов
Источник: wikiredia.ru
Здравствуйте, уважаемые читатели блога репетитора биологии по Скайпу biorepet-ufa.ru.
В предыдущей статье говорилось о том, что прежде чем приступить к решению задачи по генетике, надо иметь четкое представление к какому типу задач она относится. То есть к взаимодействиям аллельных или неаллельных генов.
В этой статье разберем, что означают названия разных вариантов заданий на взаимодействие аллельных генов: 1) полное доминирование, 2) неполное доминирование, 3) кодоминирование и 4) множественный аллелизм.
1. Полное доминирование
Это когда доминантный аллельный ген (А) в гетерозиготном организме (Аа) полностью скрывает присутствие рецессивного аллеля гена (а). Как раз явление полного доминирования отражено в 1 законе Менделя — законе единообразия гибридов первого поколения.
«>
Случайно так получилось у Менделя или не случайно, но самые известные его опыты по изучению наследования у гороха формы семян (1), цвета семян (2), цвета цветков (3), расположения цветков — пазушные или верхушечные (4), высоты растений (5), цвета бобов (6) и их формы (7) оказались именно примерами взаимодействия аллельных генов с полным доминированием одного аллеля над другим.
«>
Всегда при полном доминировании у генетических гибридов первого поколения, то есть гетерозигот — Аа, полученных от скрещивания генетически чистых гомозиготных родительских линий АА (пурпурные цветки) с аа (белые цветки), полностью исчезали фенотипически растения с белыми цветками. Все гибридные гетерозиготные растения Аа по фенотипу получались как и растения с генотипом АА — то есть с пурпурными цветками.
2. Неполное доминирование
Этим термином обозначается такой тип взаимодействия аллельных генов, когда в первом поколении от скрещивания двух чистых линий АА и аа тоже проявляется как генотипическое, так и фенотипическое единообразие гибридов первого поколения: все Аа. Но все эти гибриды фенотипически не похожи ни на одну родительскую форму, а имеют новое третье промежуточное проявление признака.
«>
Например, от скрещивания растений ночной красавицы в красными цветками (ВВ) с растениями с белыми цветками (bb), в первом поколении образуются растения с розовыми цветками (Bb).
Такой тип взаимодействия аллельных генов с неполным доминированием часто встречается в природе, в том числе и у человека. Например, у отца нос большой, а у матери — маленький, у ребенка нос может быть среднего размера.
3. Кодоминирование
Явление кодоминирования можно рассмотреть на примере проявления групп крови у человека по системе АВО. Известно, что группы крови по системе АВО в популяции человека контролируются не двумя аллелями, а тремя IО, IА, IВ Выделяю слово в «популяции», так как и в этом случае у любого конкретного человека аллельных генов, ответственных за группу крови может быть всегда только два из этих трех.
Чтобы это было понятнее, разберем как наследуются группы крови у человека, которых насчитывается 4 фенотипических и 6 генотипических.
«>Группы крови в популяции человека контролируются не двумя аллельными генами, а тремя, названными IО, IА, IВ (аллельные гены, ответственные за формирование группы крови человека обозначаются буквой I c индексами 0, А или В, приписанными сверху).
Понятно, что в организме каждого конкретного человека могут присутствовать лишь какие-то два аллеля из этих трех существующих в популяции людей.
Стало известно, что аллельные гены IА и IВ, ответственные за синтез определенных белков агглютиногенов, доминируют над третьим рецессивным аллелем IО, контролирующим отсутствие кодирования агглютиногенов.
Если в организме человека присутствуют оба рецессивных аллельных гена, то есть если его генотип IОIО, то это будет человек фенотипически с I или, так называемой, нулевой группой крови.
II и III группы крови формируются по правилу полного доминирования: генетически гомозиготный организм IАIА и гетерозиготный IАIО формируют II фенотипическую группу крови, а гомозиготный IВIВ и гетерозиготный IВIО формируют III группу крови.
Но при совместном присутствии в генотипе какого-либо человека аллелей IА и IВ ни один из них не доминирует над другим. Говорят — они кодоминантны.
Что это значит фенотипически? Эти два доминантных аллеля IА и IВ, отвечая за образование в организме человека двух разных белков агглютиногенов, обеспечат у него проявление нового свойства — IV группу крови.
4. Множественный аллелизм
Когда за определенный признак отвечает не одна пара аллельных генов, а несколько, то кроме основных (доминантного и рецессивного) генов могут присутствовать аллельные гены, находящиеся по силе влияния между этими двумя основными генами.
Эти аллельные гены обеспечивают промежуточный характер наследования: по отношению к доминантному они ведут себя как рецессивные, а по отношению к рецессивному — как доминантные.
«>Например, у кроликов сплошная черная окраска меха обусловлена доминантным геном А, гомозиготные рецессивные животные (аа) — белые. Но существуют еще два состояния этого гена, имеющих собственный фенотип в гомозиготном состоянии: шиншилловой аш и гималайской аг окрасок. Шиншилла фенотипически — сплошная серая масть, а гималайский фенотипически — белый, но кончики ушей, хвоста, ног и носа окрашены в черный цвет.
Все возможные генотипы данных четырех фенотипов окрасок представлены в табличке ниже:
«>
Вот мы разобрали основные типы взаимодействий аллельных генов.
Для тех, кто хочет быстро разобраться как надо решать такие задачи по генетике, могу предложить мою платную книжицу.
В следующей статье опишем основные типы взаимодействий неаллельных генов и вы будете готовы к решению генетических задач на все 100%.
*****************************************************************
Кто захочет быстро научиться решать задачи по генетике, предлагаю свою платную книгу.
Источник: www.biorepet-ufa.ru
Полное доминирование
При полном доминировании фенотип гетерозиготы не отличается от фенотипа доминантной гомозиготы. Видимо, в чистом виде полное доминирование встречается крайне редко или не встречается вовсе. Например, люди, гетерозиготные по гену гемофилии А (сцепленный с Х-хромосомой рецессивный ген), имеют половинное количество нормального фактора свертывания по сравнению с гомозиготными по нормальному аллелю людьми, и активность фактора свертывания VIII у них в среднем вдвое ниже, чем у здоровых людей. В то же время у здоровых людей активность этого фактора варьирует от 40 до 300 % по сравнению со средней для популяции. Поэтому наблюдается значительное перекрывание признаков у здоровых и носителей-гетерозигот. При фенилкетонурии (аутосомно-рецессивный признак) гетерозиготы обычно считаются здоровыми, однако активность печёночного фермента фенилаланин-4-гидроксилазы у них вдвое ниже нормы, а содержание фенилаланина в клетках повышено, что, по некоторым данным, приводит к снижению IQ и повышенному риску развития некоторых психотических расстройств.
Неполное доминирование
При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный — дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную — нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
При неполном доминировании во втором поколении моногибридного скрещивания наблюдается одинаковое расщепление по генотипу и фенотипу в соотношении 1:2:1.
Кодоминирование
При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов — это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Относительный характер доминирования
Как уже отмечалось выше, характер доминирования зависит от уровня анализа признака. Рассмотрим это на примере серповидно-клеточной анемии. Гетерозиготные носители гена гемоглобина S (AS) на уровне моря имеют нормальную форму эритроцитов и нормальную концентрацию гемоглобина в крови (полное доминирование А над S). На больших высотах (более 2,5-3 тыс. м) у гетерозигот концентрация гемоглобина понижена (хотя и намного выше, чем у больных), появляются эритроциты серповидной формы (неполное доминирование А над S). Этот пример показывает, что доминантность может зависеть от условий. Гетерозиготы AS и гомозиготы SS обладают примерно одинаковой устойчивостью к малярии, гомозиготы АА подвержены малярии в большей степени. По данному проявлению ген S доминирует над А. Наконец, в эритроцитах носителей АS в равных количествах присутствуют оба варианта бета-глобиновых цепей — нормальный А и мутантный S (то есть наблюдается кодоминирование).
Эволюция доминантности
Новые мутации могут, конечно, сразу обладать доминантным проявлением в фенотипе диплоидных особей, но вероятность выживания мутантов вообще невелика, и поэтому преимущественно сохраняются именно рецессивные мутации. Впоследствии, если при каких-либо изменениях внешних условий новый признак окажется благоприятным, обусловливающий его мутантный аллель может вторично приобрести доминантное фенотипическое выражение (следует подчеркнуть, что доминантны и рецессивны, собственно говоря, не сами аллели, а их проявления в фенотипе). Переход аллеля от рецессивного к доминантному состоянию может быть обусловлен различными механизмами, действующими на разных уровнях преобразований наследственной информации в онтогенезе. Генетически такой переход может быть достигнут через отбор особых генов-модификаторов, влияющих на фенотипическое проявление мутантного аллеля (гипотеза Р.Фишера), или же через отбор аллелей с большей физиологической активностью (обеспечивающих более интенсивный синтез ферментов), чем первоначальный рецессивный вариант (гипотезы С.Райта и Д.Холдейна). В сущности, эти гипотезы не исключают, а взаимно дополняют друг друга, и эволюция доминантности может происходить путем отбора малых мутаций как структурных генов, так и генов- модификаторов.
Так или иначе, степень доминантности фенотипического проявления аллелей может эволюционировать, повышаясь под контролем отбора, если данный аллель становится благоприятным для его носителя при изменениях внешних условий. Примером этого может служить повышение доминантности аллеля, контролирующего темную окраску бабочек березовой пяденицы (Biston betu-laria), которое, по некоторым данным, произошло в течение последних ста лет в индустриальных районах Европы (явление, получившее название «индустриальный меланизм») (Н. Н. Иорданский «Эволюция жизни»)
См. также
- Аллель
- Гетерозигота
- Гомозигота
- Доминантный признак
- Рецессивный признак
- Законы Менделя
- Взаимодействие неаллельных генов
Источник: dic.academic.ru