Рибосомы участвуют в синтезе
Строение
Важнейшей органеллой клетки является ядро. Оно содержит генетическую информацию и ядрышко, где образуются рибосомы. Синтезированные рибосомы через поры ядерной мембраны попадают либо на эндоплазматическую сеть, либо в цитоплазму. В зависимости от расположения в эукариотической клетке выделяют два вида рибосом:
- связанные – располагаются на эндоплазматической сети (шероховатый вид);
- свободные – располагаются в цитозоле.
Гладкая ЭПС образуется после освобождения от рибосом. В растительных клетках гладкая ЭПС формирует провакуоли, из которых затем образуются вакуоли.

Рис. 1. Расположение рибосом в клетке.
Рибосомы – немембранные органеллы, имеющие округлую форму и состоящие из двух частей – субъединиц (большой и малой), каждая из которых представляет собой смесь рибосомальной РНК (рРНК) и белков. С химической точки зрения рибосома – нуклеопротеид, состоящий из нуклеиновых кислот и протеинов.
Рис. 2. Строение рибосом.
Различают четыре разновидности молекул РНК рибосомы:
- 18S-РНК – содержит 1900 нуклеотидов;
- 5S-РНК – содержит 120 нуклеотидов;
- 5,8S-РНК – состоит из 160 нуклеотидов;
- 28S-РНК – состоит из 4800 нуклеотидов.
Малая частица рибосомы образована 30-35 белками и 18S-РНК. В большую субчастицу входит 45-50 белков и 5S-, 5,8S-, 28S-РНК.
В нерабочем состоянии части рибосом разъединены. Они соединяются с помощью информационной (матричной) РНК, обхватывая её с двух сторон. При синтезе белка рибосомы объединяются, образуя комплексы – полисомы или полирибосомы, связанные мРНК и напоминающие бусины на нитке.
Синтез белка
Главная функция рРНК – синтез белка и аминокислот.
Биосинтез белков включает два процесса:
- транскрипцию;
- трансляцию.
Транскрипция происходит с участием ДНК. Генетическую информацию считывает фермент РНК-полимераза, образуя мРНК. Далее начинается процесс трансляции, происходящий на рибосомах.
Этот процесс разделяется на три этапа:
- инициацию – начало синтеза;
- элонгацию – биосинтез;
- терминацию – завершение синтеза, отделение рибосомы.
При инициации происходит сборка рибосомы. Контактные части субъединиц называются активными центрами, между которыми располагается:
- мРНК в качестве «шаблона» синтеза;
- тРНК, осуществляющая перенос аминокислот на синтезируемую цепь;
- синтезируемый пептид, состоящий из аминокислот.
В процессе элонгации происходит удлинение полипептидной цепи за счёт присоединения аминокислот. Цепь отсоединяется от рибосомы на стадии терминации благодаря стоп-кодону – единицы генетического кода, шифрующего прекращение синтеза белка.
Рис. 3. Общая схема синтеза белка на рибосоме.
Биосинтез требует энергетических затрат. При присоединении одной аминокислоты расходуется по две молекулы АТФ (аденозинтрифосфата) и ГТФ (гуанозинтрифосфата). Кроме того, ГТФ тратится на процессы инициации и терминации.
Источник: obrazovaka.ru
Ядро — обязательная составная часть клетки эукариот. Оно контролирует и управляет деятельностью клетки, хранит и передаёт генетическую информацию.
Строение ядра одинаково для всех клеток. Ядроклетки обычно имеет размеры от 3 до 10 мкм в диаметре. В нем содержится ДНК, которая вместе с белками — гистонами образует комплексы — хромосомы, видимые в световом микроскопе при делении клетки. Хромосомы (греч. “хрома” — краска, “сома” — тело) несут генетическую информацию о структуре клетки и ее физиологической активности.
Рисунок 12 — Структура ядра клетки
Содержимое ядра отделено от цитоплазмы ядерной оболочкой, состоящей из двух близко расположенных друг к другу мембран, между которыми имеется узкая щель, заполненная полужидким веществом. Время от времени обе мембраны сливаются друг с другом, образуя ядерные поры, через которые происходит обмен различными веществами между ядром и цитоплазмой: из ядра выходят молекулы иРНК и тРНК, участвующие в синтезе различных белков, а входят белки, синтезирующиеся в цитоплазме. Внутреннее содержание ядра составляет ядерный сок — кариоплазма (греч. "karyon" — орех, ядро ореха), в ней находится одно или несколько ядрышек и значительное количество белка, РНК и ДНК (99% всей ДНК клетки), из которых образуются хромосомы.
Ядрышко — это место сборки рибосом из рибосомных белков и рибосомных ДНК, синтезируемых в цитоплазме (их может быть одно или несколько). Оно находится внутри ядра, и не имеет собственной мембранной оболочки. Основной функцией является синтез рибосом. В ядрышке локализуются белки, принимающие участие в этих процессах.
Рибосомы — немембранные органеллы. Это — важнейший органоид живой клетки, сферической или слегка эллипсоидной формы, диаметром 15-20 нм, состоящий из большой и малой субъединиц.
—
Рисунок 13 — Структура и схема рибосомы
Рибосомы обнаружены в клетках всех организмов. В их состав входят белки и РНК. Каждая субъединица состоит из нескольких десятков белков. Белки в рибосоме держатся на каркасе, состоящем из рибосомной РНК.
Рибосомы служат для биосинтеза белка из аминокислот по заданной матрице на основе генетической информации, предоставляемой матричной РНК или мРНК. Также существуют транспортные РНК – тРНК, которые поставляют необходимые аминокислоты для составления пептидной цепи. Транспортная РНК входит в рибосому, комплементарно связываясь с кодоном мРНК, затем происходит реакция при которой аминокислотные остатки связываются друг с другом, а тРНК удаляется. "Словарь" для перевода с языка нуклеотидов на язык аминокислот называется генетическим кодом.
Рисунок 14 — Схема биосинтеза белка
Иногда этот процесс осуществляется не одной рибосомой, а целой группой рибосом (такую группу называют полисомой).
Рисунок 15 -Полисома
Клеточный центр играет исключительную роль в организации цитоскелета: многочисленные цитоплазматические микроклубочки расходятся от него во все стороны. В центре клеточного центра находятся две центриоли. Каждая центриоль представляет собой цилиндр (длиной 0,3 мкм и диаметром 0,1 мкм), по окружности которого располагается девять триплетов микротрубочек. Центриоли образуют пары, члены которых расположены под прямым углом друг к другу.
ред делением клетки члены пары расходятся к противоположным полюсам и возле каждой из них возникает дочерняя центриоль. От центриолей, расположенных на разных полюсах клетки, протягиваются друг к другу параллельные микротрубочки, образуя митотическое веретено, способствующее равномерному распределению генетического материала между дочерними клетками. Часть нитей веретена прикрепляется к хромосомам. Однако центриоли обнаружены не у всех клеток, имеющих клеточный центр. Нет их и у высших растений.
а) б)
Рисунок 24 — а) центриоль с 9 триплетами микротрубочек;
б) пара центриолей: 1 — материнская ; 2 — дочерняя
7 Мембранные органоиды клетки
Рибосомы из ядрышка поступают (через поры в ядерной оболочке) на мембраны эндоплазматической сети (ЭПС) — системы соединенных между собой канальцев и полостей различной формы и величины, контактирующей со всеми органоидами клетки.
ЭПС бывают двух видов — шероховатая и гладкая: на шероховатой ЭПС (или гранулярной) располагается множество рибосом, которые осуществляют синтез белков. Рибосомы придают мембранам шероховатый вид. Мембраны гладкой ЭПС не несут рибосом на своей поверхности, в них располагаются ферменты синтеза и расщепления углеводов и липидов. Гладкая ЭПС выглядит как система тонких трубочек и цистерн.
рибосомы
а) б)
Рисунок 16 – Эндоплазматическая сеть а) шероховатая;
б) вверху шероховатая, ниже гладкая ЭПС
Продукты синтеза (белки, жиры и углеводы), образовавшиеся в каналах и полостях ЭПС, транспортируются к аппарату Гольджи.
Комплекс Гольджи – это органоид клетки, основой которого является гладкая мембрана, образующая пакеты уплощённых цистерн,
уложенных в стопку, и крупных и мелких пузырьков, расположенных на концах полостей.
Рисунок 17 – Аппарат Гольджи
Все поступившие в аппарат Гольджи вещества накапливаются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму, к органоидам клетки, где потребляются или выделяются из клетки.
Рисунок 18 – Микрофотография аппарата Гольджи.
Наряду с образованием белков, жиров и углеводов и т.д., ЭПС клетки вырабатывает специфические вещества белковой природы – ферменты, которые, накапливаясь в аппарате Гольджи выделяются в виде лизосом — небольших округлых телец. Лизосомы (греч. “лизео” — растворяю, “сома” — тело) — самые мелкие мембранные образования, представляющие собой пузырьки, диаметром 0,5 мкм, содержат ферменты, расщепляющие белки, углеводы, жиры и нуклеиновые кислоты. Лизосомы участвуют в расщеплении старых “частей” клетки, целых клеток и отдельных органов. Например, исчезновение хвоста у головастика лягушек происходит под действием ферментов лизосом.
Выделяющиеся из аппарата Гольджи пузырьки с водой движутся к вакуолям.
Вакуоли — мембранные органеллы, являющиеся резервуарами воды с растворенными в ней соединениями. В растительных клетках на долю вакуолей приходится до 90% объема, а животные клетки имеют временные вакуоли, занимающие не более 5% их объема.
Вакуоли
Рисунок 19 – Вакуоли в клетке
Вакуоли растительных клеток поддерживают тургорное давление и поставляют воду, используемую при фотосинтезе.
ЭПС, аппарат Гольджи, лизосомы и вакуоли составляют систему, отдельные элементы которой могут переходить друг в друга при перестройке и изменении функций мембран.
Рисунок 20 — Система образования и выделения веществ через ЭПС и аппарат Гольджи.
Цитоплазма большинства растительных и животных клеток содержит “энергетические станции” — митохондрии.
Митохондрииимеют палочковидную, нитевидную или шаровидную форму диаметром около 1 мкм и длиной около 7 мкм. Митохондрии (греч. “митос” — нить, “хондрион” — зерно, гранула) хорошо видны в световой микроскоп, имеют наружную гладкую мембрану и внутреннюю мембрану, имеющую многочисленные складки — кристы, в которые встроены ферменты, участвующие в преобразовании энергии питательных веществ, поступающих в клетку, в энергию молекул АТФ. Число крист (лат. “криста” — гребень, вырост) неодинаково в разных митохондриях клеток. Их может быть от нескольких десятков до нескольких сотен и даже тысяч: чем больше энергетических затрат осуществляет данная клетка, тем больше она содержит митохондрий. Внутреннее пространство митохондрий заполнено гомогенным веществом, называемым матриксом. Вещество матрикса более плотное, чем то, которое окружает митохондрию.
В матриксе присутствуют нити ДНК и РНК, а также рибосомы, что обеспечивает митохондриям самовозобновление путем деления. Митохондрии тесно связаны с мембранами эндоплазматической сети, каналы которой часто открываются непосредственно в митохондрии.
Количество митохондрий меняется в процессе индивидуального развития организма (онтогенеза): в молодых растущих и делящихся клетках их значительно больше, чем в стареющих.
Рисунок 21 — Митохондрия
Цитоплазма растительных клеток содержит пластиды, животные клетки их не имеют. Различают три основных типа пластид: лейкопласты, хромопласты и хлоропласты. Они имеют разную окраску. Бесцветные лейкопласты находятся в цитоплазме клеток неокрашенных частей растений: стеблях, корнях, клубнях. Например, их много в клубнях картофеля, в которых накапливаются зерна крахмала. Хромопласты находятся в цитоплазме цветков, плодов, стеблей, листьев. Хромопласты обеспечивают желтую, красную, оранжевую окраску растений. Зеленые хлоропласты содержатся в клетках листьев, стеблей и других частях растения, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, они часто имеют овальную форму. У высших растений в одной клетке содержится несколько десятков хлоропластов.
Рисунок 22 — Пластиды
Зеленые хлоропласты способны переходить в хромопласты — поэтому осенью листья желтеют, а зеленые помидоры краснеют при созревании. Лейкопласты могут переходить в хлоропласты (позеленение клубней картофеля на свету). Таким образом, хлоропласты, хромопласты и лейкопласты способны к взаимному переходу.
Основная функция хлоропластов — фотосинтез, т.е. в хлоропластах на свету осуществляется синтез органических веществ из неорганических за счет преобразования солнечной энергии в энергию молекул АТФ. Хлоропласты высших растений имеют размеры 5-10 мкм и по форме напоминают двояковыпуклую линзу. Каждый хлоропласт окружен двойной мембраной, обладающей избирательной проницаемостью. Снаружи располагается гладкая мембрана, а внутренняя имеет складчатую структуру. Основная структурная единица хлоропласта – тилакоид, плоский двумембранный мешочек, ирающий ведущую роль в процессе фотосинтеза. В мембране тилакоида расположены белки, аналогичные белкам митохондрий, которые участвуют в цепи переноса электоронов. Тилакоиды расположены стопками, напоминающие стопки монет (от 10 до 150) и называемыми гранами. Грана имеет сложное строение: в центре располагается хлорофилл, окруженный слоем белка; затем располагается слой липоидов, снова белок и хлорофилл.
В каждом хлоропласте примерно по 50 гран, расположенных в шахматном порядке. В мембранах, формирующих тилакоиды, содержатся ферменты, улавливающие солнечный свет и синтезирующие АТФ. Внутренняя среда хлоропласта содержит ферменты, синтезирующие органические вещества с использованием энергии АТФ. . Каждый хлоропласт содержит ДНК и рибосомы и способен к автономному делению, как и митохондрии. Зеленый цвет хлоропластов обусловлен содержанием в них пигмента хлорофилла, имеющего сложное химическое строение. В живом и функционирующем хлоропласте содержится до 75 % воды.
Рисунок 23 — Хлоропласт
Размеры, форма митохондрий и хлоропластов, наличие в них двуцепочечной ДНК и собственных рибосом делают их похожими на клетки бактерий. На основании этого сходства существует теория симбиотического происхождения эукариотической клетки, в соответствии с которой полагают, что предки современных митохондрий и хлоропластов были когда-то самостоятельными прокариотическими организмами.
Помимо различных органоидов клетка имеет включения — непостоянные образования, которые то возникают, то исчезают. Включения являются продуктами метаболизма и локализуются в основном в цитоплазме клетки в виде гранул, зерен, капель и кристаллов. Липоиды откладываются в виде мелких капель, полисахариды — в виде гранул (зерна крахмала, гранулы гликогена); белковые соединения откладываются реже (тоже в виде гранул, есть шарики, палочки, пластинки), они есть в яйцеклетках, печени, в цитоплазме простейших и многих других животных. К клеточным включениям относятся некоторые пигменты (липофуцин, образующийся главным образом при старении организма; липохромы находящиеся в яичниках и надпочечниках; ретинин, входящий в состав зрительного пурпура; гемоглобин крови; меланин кожи и другие пигменты). Еще встречаются секреторные включения, чаще располагающиеся в железистых клетках: они могут быть белковыми, сахарами, липопротеидами и т.д.
8 Питание клетки. Фагоцитоз и пиноцитоз.
Любая живая клетка питается, т.е. захватывает из внешней среды питательные вещества (в виде отдельных молекул или больших групп молекул — пищевых частиц, иногда даже целых клеток меньшего размера), и так или иначе использует эти вещества.
Есть всего два принципиально различных варианта использования питательных веществ.
1. Молекулы питательных веществ можно использовать для построения других молекул, выполняющих в жизни клетки какие-то функции, например, молекул, входящих в состав клеточной мембраны. Этот вариант использования клеткой питательных веществ называется ассимиляцией.
2. Другой вариант – получение энергии, которая при этом выделяется и используется клеткой, например, для передвижения или для захвата новых пищевых частиц. Такой вариант использования веществ называется диссимиляцией.
Для переноса воды и различных ионов в клеточной мембране существуют поры, через которые они пассивно поступают в клетку. Кроме того, существует активный перенос веществ в клетку с помощью специальных белков, входящих в состав плазматической мембраны. Он осуществляется также на основе процессов фагоцитоза и пиноцитоза
Фагоцитоз ("фагос" — "пожиратель", "цитос" — "клетка") — питание клетки сравнительно большими пищевыми частицами (в том числе другими клетками). Общая картина фагоцитоза показана на рис. 9.
Рисунок 9- Фагоцитоз. Пиноцитоз. Рецепторный эндоцитоз
Проплывающая мимо клетки пищевая частица касается мембраны и прилипает к ней. Мембрана под ней прогибается, охватывая частицу со всех сторон. В результате образуется мембранный пузырек с частицей внутри — пищеварительная вакуоль. Она отрывается от мембраны и уплывает вглубь цитоплазмы. Там она сливается с другим пузырьком (первичной лизосомой, отделившимся от комплекса Гольджи. Пузырек — результат этого слияния — называют вторичной лизосомой. После этого пищевая частица начинает растворяться. Минут через 20 внутри вторичной лизосомы виднеются только несколько маленьких бесформенных кусочков, почему-то "не захотевших" растворяться. Затем вторичная лизосома подплывает к мембране клетки и сливается с ней, выбрасывая из клетки наружу эти "кусочки" (рисунок 20).
Другой вариант, гораздо более приемлемый для многоклеточных животных – вторичная лизосома выбрасывает непереваренные остатки в специальную вакуоль накопления на «вечное хранение».
Все эти удивительные превращения происходят благодаря деятельности специальных молекул. Специальные молекулы мембраны клетки (рецепторы), обеспечивают прилипание пищевой частицы к мембране и образование пищеварительной вакуоли. Рецепторы — это молекулы мембраны клетки, которые могут узнавать другие молекулы (лиганды), и прочно к ним прилипать. Коснувшаяся мембраны частица прилипает в том случае, если на ее поверхности имеются лиганды к каким-нибудь рецепторам, имеющимся на поверхности клетки (на мембране обычно имеется около 100 различных разновидностей рецепторов, и каждый из них "узнает" определенный лиганд).
В случае, когда клетка захватила с помощью фагоцитоза другую маленькую клетку, первичная лизосома приносит из комплекса Гольджи специальные молекулы (пищеварительные ферменты), умеющие "разрезать" большие молекулы (полимеры) на части. Из-за этого органоиды захваченной клетки "разваливаются" на отдельные мелкие молекулы. В мембране вторичной лизосомы имеются также белки-переносчики, которые умеют переносить эти мелкие молекулы через мембрану в цитоплазму клетки.
Пиноцитоз (греч. "pino" — пить) — процесс захвата и поглощения капелек жидкости с растворенными в ней веществами. Пиноцитоз напоминает фагоцитоз, но фагоцитоз широко распространен у животных, а пиноцитоз осуществляется как растительными, так и животными организмами.
Клеточная стенка растений и бактерий препятствует фагоцитозу и поэтому фагоцитоз у них практически отсутствует.
ТЕМА: ТКАНЕВЫЙ УРОВЕНЬ
Тканевый уровень представлен тканями, объединяющими клетки определенного строения, размеров, расположения и сходных функций. Ткани возникли в ходе исторического развития вместе с многоклеточностью. У многоклеточных организмов они образуются в процессе онтогенеза как следствие дифференциации клеток. У животных различают несколько типов тканей (эпителиальная, соединительная, мышечная, нервная, а также кровь и лимфа). У растений различают меристематическую, защитную, основную и проводящую ткани. На этом уровне происходит специализация клеток.
Функции, выполняемые животным организмом, очень разнообразны, поэтому и клетки в нем построены неодинаково. По внешним, или морфологическим, признакам можно выделить однородные группы клеток, из которых как бы соткан организм; отсюда произошло название ткани, т. е. различные группы клеток. Каждая группа однородных клеток выполняет определенную функцию и обладает особыми, только ей присущими качествами.
Ни одна из тканей не является независимой, изолированной группой однородных клеток. Только при самой тесной работе всех клеток как частей целого организма возможна их жизнь.
На основании особенностей строения и функции клеток различают следующие ткани: эпителиальную, соединительную, мышечную и нервную.
1. Эпителиальная ткань.
Эпителиальная ткань, или эпителий, характеризуется тем, что клетки располагаются в нем целыми рядами, одна возле другой. Эпителий очень распространен в сложном организме. Он покрывает поверхность тела животного, полости и органы, выполняющие различную физиологическую роль в организме. Эпителий защищает внутренние ткани, и проникнуть к этим тканям можно, только нарушив эпителий.
Функциональное значение эпителия разнообразно, и построен он в различных местах тела неодинаково. Там, где клетки эпителия располагаются в один ряд, он называется однослойным; там, где ряды клеток наслаиваются один на другой, – многослойным.
Различают однослойный цилиндричский эпителий, который, в свою очередь, делится на мерцательный, каемчатый и железистый, а также многослойный эпителий.
Мерцательный эпителий покрывает дыхательные пути, яйцеводы и характеризуется наличием тонких подвижных нитей на свободном конце клеток, называемых ресничками. Они постоянно двигаются в одну сторону, вследствие чего из дыхательных путей выделяются мокрота, различные посторонние частицы, а в яйцеводах происходит перемещение яйцевой клетки в матку.
Каемчатый, или кишечный, эпителий покрывает внутреннюю поверхность кишечника. На свободном конце клеток этого эпителия имеется особое приспособление – кайма, или кутикула, при помощи которой в стенки кишечника всасываются растворенные в воде питательные вещества.
Железистый эпителий находится главным образом в железах. Клетки железистого эпителия выделяют специальную жидкость, называемую секретом. Форма и строение железистых клеток очень разнообразны, как и выделяемый ими секрет.
Многослойный эпителий в зависимости от формы клеток подразделяют на: 1) многослойный цилиндрический, встречающийся редко, главным образом в выводных протоках желез; 2) многослойный переходный, отличающийся большой растяжимостью и выстилающий полости, сильно изменяющие свой объем (например, полость мочевого пузыря); 3) многослойный плоский, состоящий из плоских клеток, которые ороговевают. Он покрывает снаружи тело животного, выстилает внутри ряд органов (полость рта, глотку, пищевод и др.), являясь защитным эпителием.
2. Соединительные ткани
Соединительные ткани распространены по всему организму. Они связывают различные части тела между собой. Соединительные ткани подразделяются на две основные группы: ткани питающие (трофические) и опорные (механические).
Кровь и лимфа по своему происхождению относятся к трофической группе соединительной ткани. В состав крови входят плазма и форменные элементы.
Плазма представляет собой жидкую часть крови и состоит из воды, неорганических и органических веществ. Одни из них являются питательным материалом для клеток, другие – продуктами обмена веществ, подлежащими удалению из организма.
В крови, находящейся вне организма, плазма свертывается, причем выпадает белковое вещество – фибрин, образующий тромб. Способность крови образовывать тромб предохраняет от кровотечений при нарушении целостности кровеносного сосуда.
Жидкость, остающаяся после удаления фибрина, называется кровяной сывороткой.
К группе механических соединительных тканей относятся хрящевая и костная ткани. Хрящевая ткань встречается там, где требуется большая упругость (остов дыхательного аппарата), или там, где необходимо смягчать толчки и сотрясения (на концах костей в суставах).
Рисунок 28 — Строение хрящевой ткани: 1 — межклеточное вещество; 2 — клетка; 3 – ядро
Костная ткань – самая прочная в организме. В ней, кроме органических соединений, много минеральных веществ, а именно фосфорно-кальциевых солей. Это придает костной ткани большую крепость, а наличие органических веществ – упругость. Кость пронизана каналами, через которые проходят кровеносные и лимфатические сосуды, а также нервные волокна. Стенки костей состоят из сплошного компактного вещества, а внутри кость построена из губчатого вещества, пустые пространства которого заполнены костным мозгом.
Рисунок 27- Строение костной ткани: 1 — костная клетка (остеоцит); 2 — ядро; 3 — межклеточное вещество
Кроме того, существует волокнистая соединительная ткань, выполняющая, помимо опорной, и трофическую функцию, так как в ее межклеточных щелях циркулируют питательные вещества.
Волокнистая соединительная ткань бывает рыхлой, плотной и эластической. Рыхлая соединительная ткань залегает под кожей между мускулами и служит для соединения и образования остова отдельных органов. Плотная соединительная ткань встречается в сухожилиях, связках и других органах и отличается от рыхлой плотностью и прочностью.
Рисунок 26 — Строение плотной соединительной ткани: 1 — коллагеновые волокна; 2 — ядро; 3 — клетки: 4 — эластиновые волокна
Эластическая соединительная ткань характеризуется большим количеством эластических волокон, прочностью, достаточной упругостью; встречается она в различных связках и крупных кровеносных сосудах.
5. Мышечная ткань
Мышечная ткань имеет своеобразные клетки, сильно вытянутые в длину, почему они и получили название мышечных волокон. Различают гладкую и поперечнополосатую мышечную ткань
Рисунок 29- Строение мышечной ткани: 1 — мышечная клетка (мышечное волокна); 2 — ядра; 3 — межклеточное вещество; 4 — волокно межклеточного вещества
Гладкая мышечная ткань сокращается независимо от воли животного. Она распространена во внутренних органах тела: пищеварительных, дыхательных и мочеполовых; в сосудах, в селезенке и т. д.
Поперечнополосатая мышечная ткань подразделяется на скелетную и сердечную. Скелетная мышечная ткань находится на тех частях скелета, которые участвуют в движении; она сокращается по произволу, почему ее и называют мышечной тканью произвольного движения. Сердечная мышечная ткань имеется в сердце и функционирует независимо от воли животного. Ее особенностью являются правильно чередующиеся сокращения, т. е. ритм.
6. Нервная ткань
Нервная ткань предназначена в организме для восприятия и передачи раздражений как внутри организма, так и при общении его с внешней средой. Через нервную ткань, животные воспринимают самые разнообразные ощущения: свет, цвет, запах, вкус, звук и пр.
ТЕМА: Организменный уровень развития живого
Онтогенез (от греч. ontos – существо, geneses – развитие) – это цикл развития индивидуального организма (животного или растения), начинающийся с образования давших ему начало половых клеток и заканчивающийся его смертью.
Онтогенез – индивидуальное развитие организма
Филогенез.- история возникновения и развития вида (животных или растений).
В Х1Х веке немецкими учеными Фрицем Мюллером и Эрнестом Геккелем был сформулирован биогенетический закон:
«Онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития ) вида , к которому эта особь относится »
Онтогенез в зависимости от характера развития организмов типируют на прямой и непрямой
Прямое развитие организмов в природе встречается в виде неличиночного и внутриутробного развития, тогда как непрямое развитие наблюдается в форме личиночного развития.
1.Сущность роста и развития организмов.
Итак, после оплодотворения яйцеклетки начинается рост и развитие нового живого организма, который повторяет путь развития родителей – отца и матери. Это – очень сложный процесс и заключается во взаимодействии наследственности, полученной от родителей, и условий среды, окружающей растущий организм.
Рост организма это постепенное увеличение его массы в результате увеличения количества клеток.
Рост можно измерить, построив на основе результатов измерений кривые размеров организма, массы, сухой массы, количества клеток, содержания азота и других показателей. При этом иногда одни клетки становятся морфологически, биохимически и функционально отличными от других клеток.
Размножение и дифференцировка одних клеток всегда координированы с ростом и дифференцировкой других. Оба эти процесса происходят на протяжении всего жизненного цикла организма. Поскольку дифференцирующиеся клетки изменяют свою форму, а в изменения формы вовлекаются группы клеток, то это сопровождается морфогенезом, который определяет структурную организацию клеток и тканей, а также общую морфологию организмов.
Таким образом, рост является результатом количественных изменений в виде увеличения количества клеток (массы тела) и качественных – в виде дифференцировки клеток и морфогенеза. Рост может быть неопределенным – продолжающимся всю жизнь (у растений), и определенным, ограниченным каким–либо сроком (у многих животных рост прекращается вскоре после достижения полового созревания).
Источник: studopedia.su
О генетическом коде
Программа, задающая последовательность аминокислотных остатков в белке, записана в геноме клетки: около полувека назад было установлено, что аминокислотные последовательности всех белков непосредственно закодированы в ДНК с помощью так называемого генетического кода. Согласно этому коду, универсальному для всех живых организмов, каждой из двадцати существующих аминокислот соответствует свой кодон — тройка нуклеотидов, представляющих собой элементарные единицы цепочки ДНК. Любой белок закодирован в ДНК определенной последовательностью кодонов. Эта последовательность называется геном.
Одна клетка может содержать до 10 тыс. рибосом — белковых минифабрик, составляющих до 30% сухой клеточной массы
Как же эта генетическая информация доходит до рибосомы? На отдельном гене, как на матрице, синтезируется цепь еще одной информационной молекулы — рибонуклеиновой кислоты (РНК). Этот процесс копирования гена, называемый транскрипцией, осуществляется специальными ферментами — РНК-полимеразами.
Но РНК, полученная таким образом, еще не является матрицей для синтеза белка: из нее, вырезаются определенные «некодирующие» куски нуклеотидной последовательности (процесс сплайсинга).
Точность белкового синтеза рибосомой исключительно высока — у человека ошибка составляет один на три тысячи «неправильный» аминокислотный остаток
В результате получается матричная РНК (мРНК), которая и используется рибосомами в качестве программы для синтеза белка. Сам синтез, т.е. перевод генетической информации с языка нуклеотидной последовательности мРНК на язык аминокислотной последовательности белка, называется трансляцией.
Декодирование и синтез
В клетках эукариот одну мРНК обычно транслирует сразу множество рибосом, образуя так называемые полисомы, которые можно отчетливо видеть с помощью электронной микроскопии, позволяющей получать увеличение в десятки тысяч раз.
Как поступают в рибосому аминокислоты, являющиеся строительными кирпичиками для синтеза белка? Еще в 50-х годах прошлого столетия были открыты особые «перевозчики», доставляющие аминокислоты в рибосому, — короткие (длиной менее 80 нуклеотидов) транспортные РНК (тРНК). Специальный фермент присоединяет аминокислоту к одному из концов тРНК, причем каждой аминокислоте соответствует строго определенная тРНК. Синтез белка на рибосоме включает три основные стадии: начало, удлинение полипептидной цепи и окончание.
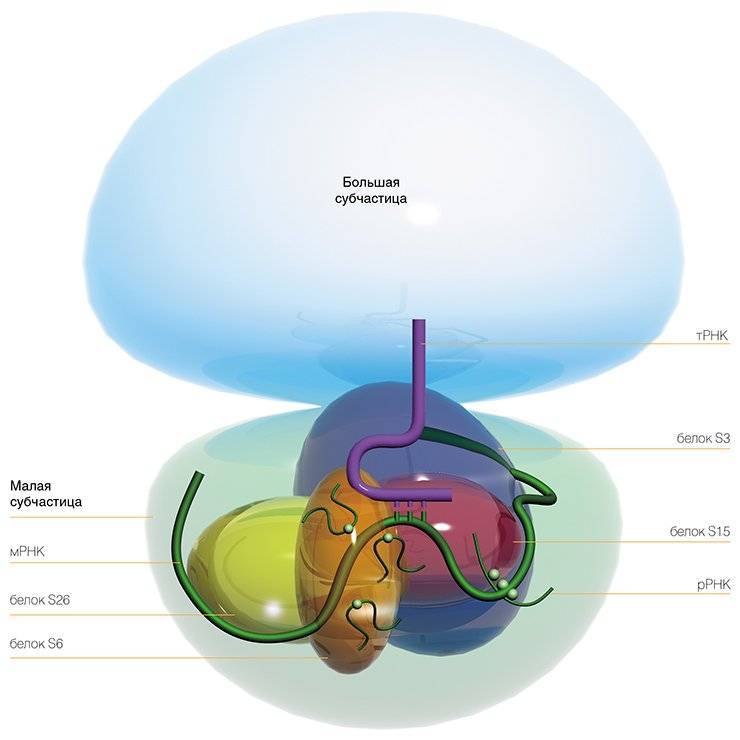
Сама рибосома — одна из самых сложно организованных молекулярных машин клетки — состоит из двух неравных частей, так называемых субчастиц (малой и большой). Ее можно легко разделить на части центрифугированием при сверхвысоких скоростях в специальных пробирках с раствором сахарозы, концентрация которой увеличивается сверху вниз. Поскольку малая субчастица в два раза легче большой, они движутся от верха пробирки к дну с разными скоростями.
Малая субчастица отвечает за декодирование генетической информации. Она состоит из высокомолекулярной рибосомной РНК (рРНК) и нескольких десятков белков (около 20 у прокариот и более 30 — у эукариот).
В раковых клетках резко повышается уровень некоторых рибосомных белков. Возможная причина — сбои в механизмах авторегуляции их производства
Большая субчастица, ответственная за образование пептидной связи между аминокислотными остатками, состоит из нескольких рРНК: одной высокомолекулярной и одной (или двух в случае эукариот) низкомолекулярной, а также нескольких десятков белков (более 30 у прокариот и до 50 у эукариот). О масштабе деятельности рибосом можно судить хотя бы по тому факту, что рибосомная РНК составляет около 80 % всей РНК клетки, тРНК, транспортирующая аминокислоты, — около 15 %, тогда как матричная РНК, несущая информацию о белковой последовательности, — лишь 5 %!
Нужно отметить, что рибосомные белки наделены множеством других, дополнительных функций, которые могут проявляться на разных этапах жизнедеятельности клетки. Например, рибосомный белок S3 человека — один из ключевых белков центра связывания мРНК на рибосоме — принимает также участие в «ремонте» повреждений в ДНК (Kim et al., 1995), участвует в апоптозе (запрограммированной гибели клетки) (Jung et al., 2004), а также защищает от разрушения белок теплового шока (Kim et al., 2006).
Кроме того, чересчур интенсивный синтез некоторых рибосомных белков может свидетельствовать о развитии злокачественной трансформации клетки. Например, значительное повышение уровня пяти рибосомных белков было обнаружено в опухолевых клетках толстого кишечника (Zhang et al., 1999). Недавно сотрудниками лаборатории структуры и функции рибосом ИХБФМ СО РАН был открыт новый механизм авторегуляции биосинтеза рибосомных белков у человека, основанный на принципе обратной связи. Неуправляемый синтез рибосомных белков, характерный для опухолевых клеток, вероятно, вызван сбоями именно в этом механизме. Дальнейшие исследования в этой области представляют особый инте¬рес не только для ученых, но и для медиков.
Работает как «рибозим»
Удивительно, но, несмотря на миллиарды лет эволюции, разделяющие бактерии и человека, вторичная структура рибосомальных РНК у них мало различается.
О том, как уложена рРНК в субчастицах и каким образом она взаимодействует с рибосомными белками, до недавнего времени было известно не много. Революционный сдвиг в понимании устройства рибосомы на молекулярном уровне произошел на рубеже нового тысячелетия, когда с помощью рентгеноструктурного анализа удалось расшифровать на уровне отдельных атомов структуру рибосом простейших организмов и их модельных комплексов с мРНК и тРНК. Это позволило понять молекулярные механизмы декодирования генетической информации и образования связей в молекуле белка.
Оказалось, что оба важнейших функциональных центра рибосомы — как декодирующий на малой субчастице, так и отвечающий за синтез белковой цепочки на большой субчастице — сформированы не белками, а рибосомной РНК. То есть, рибосома работает подобно рибозимам — необычным ферментам, состоящим не из белков, а из РНК.
Рибосомные белки, тем не менее, также играют важную роль в работе рибосомы. В отсутствие этих белков рибосомные РНК совершенно неспособны ни декодировать генетическую информацию, ни катализировать образование пептидных связей. Белки обеспечивают необходимую для работы рибосомы сложную «укладку» рРНК в функциональных центрах, служат «передатчиками» изменений пространственной структуры рибосомы, необходимых в процессе работы, а также связывают различные молекулы, влияющие на скорость и точность процесса белкового синтеза.
Сама рабочая схема белкового цикла в принципе одинакова для рибосом всех живых существ. Однако до сих пор неизвестно, до какой степени схожи молекулярные механизмы работы рибосом у разных организмов. Особенно не хватает информации об устройстве функциональных центров рибосом высших организмов, которые изучены гораздо хуже, чем рибосомы простейших.
Это связано с тем, что многие из методов, успешно использованных для исследования рибосом прокариот, оказались для эукариот неприменимыми. Так, из рибосом высших организмов не удается получить кристаллы, пригодные для рентгеноструктурного анализа, а их субчастицы невозможно «собрать» в пробирке из смеси рибосомных белков и рРНК, как это делается у простейших.
От низших — к высшим
И все-таки способы получения сведений о строении функциональных центров рибосом высших организмов существуют. Одним из таких методов является метод химического аффинного сшивания, разработанный 35 лет назад в отделе биохимии НИОХ СО АН СССР (ныне ИХБФМ СО РАН) под руководством академика Д. Г. Кнорре.
Метод основан на использовании коротких синтетических мРНК, несущих в выбранном положении химически активные («сшивающие») группы, которые в нужный момент можно активировать (например, облучая мягким ультрафиолетовым светом).
Метод аффинного химического сшивания был разработан 35 лет назад в отделе биохимии НИОХ СО АН СССР (ныне ИХБФМ СО РАН) под руководством академика Д. Г. Кнорре.До появления рентгеноструктурного анализа рибосом он использовался во всем мире для изучения рибосом у прокариот.
Этот метод и сегодня является основным для изучения структурно-функциональной организации рибосом высших организмов
Достоинство этого метода в том, что сшивающую группу можно присоединить практически к любому нуклеотидному остатку мРНК и в результате получить детальную информацию о его окружении на рибосоме. Используя набор коротких мРНК с разным расположением сшивающей группы, нам удалось определить рибосомные белки и нуклеотиды рРНК рибосомы человека, образующие канал для считывания генетической информации в процессе трансляции.
Впервые экспериментально удалось показать, что все нуклеотиды рРНК малой рибосомной частицы человека, соседствующие с кодонами мРНК, расположены в консервативных районах вторичной структуры молекулы рРНК. Более того, их расположение совпадает с положением соответствующих нуклеотидов во вторичной структуре рРНК рибосом низших организмов. Это позволило сделать вывод, что эта часть рибосомной РНК малой субчастицы составляет эволюционно консервативный «кор» (сердцевину) рибосомы, структурно идентичный у всех организмов.
С другой стороны, в устройстве мРНК-связывающего канала рибосом у человека и низших организмов обнаружен ряд принципиальных различий. Оказалось, что у высших организмов рибосомные белки играют намного большую роль в формировании этого канала, чем у прокариот, кроме того, в этом участвуют также белки, не имеющие «двойников» (гомологов) у низших организмов.
Почему же, несмотря на то, что функция рибосомы практически не изменилась в процессе эволюции, в организации декодирующего центра рибосом у высших организмов появились специфичные черты? Вероятно, это связано с более сложной и многостадийной регуляцией белкового синтеза у эукариот по сравнению с прокариотами, в ходе которой рибосомные белки мРНК-связывающего канала могут взаимодействовать не только с мРНК, но и с различными факторами, влияющими на эффективность и точность трансляции. Так ли это, покажут дальнейшие исследования.
Источник: scfh.ru