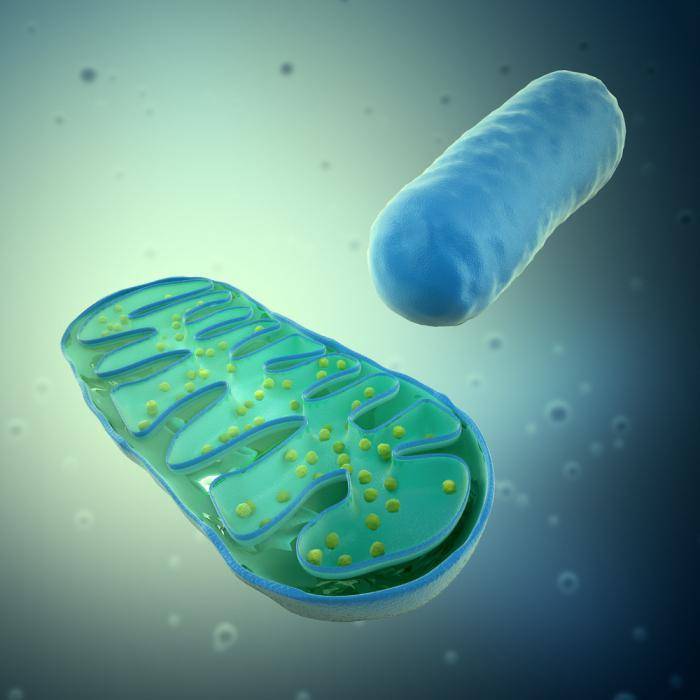
Що таке мітохондрії
У рослин мітохондрії були вперше виявлені в 1904 р. Ф.Мевесом в клітинах пиляків латаття. Вони являють собою округлі чи гантелеподібні тіла, розміри яких надзвичайно мінливі і значною мірою залежать від функціонального стану у клітин, осмотичного тиску і рН середовища. Мітохондрії збільшуються в гіпотонічних речовинах і зменшуються в гіпертонічних, а в кислому середовищі набувають пухирчастої форми. Товщина мітохондрії постійна (близько 1.5 мкм), у той час як довжина помітно коливається, досягаючи 7-10 мкм і більше.
Мітохондрії обмежені двома мембранами (рис. 1.19).

Міжмембранний ростір, чи перимітохондріальний простір
, заповнений основною безструктурною речовиною, що містить глобулярні білки і деякі ферменти. Зовнішня мембрана гладенька, а внутрішня утворює численні гребнеподібні складки — кристи
. Вони істотно збільшують її поверхню, забезпечуючи площу для розміщення мультиферментиних систем
. Мембрани відокремлюють від цитоплазми внутрішній вміст мітохондрій — матрикс
. У матриксі містяться рибосоми і мітохондріальна ДНК, що має кільцеву будову.
Сучасні методи дозволили виявляти присутність особливих «елементарних часток» на внутрішній мітохондріальній мембрані (рис.1.20). Це ферменти АТФ — синтетази, що забезпечують сполучення фосфорилювання АДФ із реакціями в дихальному ланцюзі.
В основі цих часток розташовані компоненти самого дихального ланцюга. Таким чином, основна задача мітохондрій
– це синтез АТФ у результаті циклічного окислювання ди- і три карбонових кислот і аеробних реакцій електронотранспортного ланцюга дихання.
Кількість мітохондрій варіює від десятків до десятків тисяч на клітину, змінюючись в онтогенезі, отже вона визначається рівнем метаболізму. У залежності від ділянки клітини, де необхідні електричні витрати, мітохондрії з течією цитоплазми переміщуються в ту чи іншу ділянку.
Подібні статті
Будова, функції та методи дослідження мітохондрій
Мітохондрії (від греч. mitos — нитка і chondrion — зернятко, крупинка), хондріосоми, які постійно присутні в клітинах тварин і рослин, органоїд, який забезпечує клітинне дихання, у результаті якого енергія вивільнюється або акумулюється в …
Механізми раннього ембріонального розвитку
Керівники тваринницьких господарств звичайно зацікавлені у підвищенні продуктивності сільськогосподарських тварин. Їх кінцевою метою є підвищення якості та кількості продукції без зростання видатків на утримання поголів’я. Біотехнологічні …
Источник: www.grandbiology.com
Что такое митохондрии?
Митохондрии – это энергетическая база клеток, их деятельность основана на окислении органических соединений и применении энергии, освободившейся при распаде молекул АТФ. Биологи на простом языке его называют станцией вырабатывания энергии для клеток.В 1850 г. митохондрии выявили в виде гранул в мышцах. Их число менялось в зависимости от условий роста: они скапливаются больше в тех клетках, где большой дефицит кислорода. Это происходит чаще всего при физических нагрузках. В таких тканях появляется острая нехватка энергии, которую восполняют митохондрии.
Появление термина и место в теории симбиогенеза
В 1897 г. Бенд впервые ввел понятие «митохондрия», чтобы обозначить зернистую и нитчатую структуру в цитоплазме клеток. По форме и величине они разнообразны: толщина составляет 0,6 мкм, длина – от 1 до 11 мкм. В редких ситуациях митохондрии могут быть большого размера и разветвленным узлом.
В теории симбиогенеза дается четкое представление о том, что такое митохондрии и как они появились в клетках. В ней говорится, что хондриосома возникла в процессе поражения клетками бактерий, прокариотами. Так как они не могли автономно применять кислород для выработки энергии, это препятствовало полному их развитию, а прогеноты могли развиваться беспрепятственно. В течение эволюции связь между ними дала возможность прогенотам передать свои гены теперь уже эукариотам. Благодаря такому прогрессу митохондрии теперь не являются независимыми организмами. Их генофонд не может быть реализован в полной мере, так как происходит его частичная блокировка ферментами, которые есть в любой клетке.
Где они живут?
Митохондрии сосредотачиваются в тех районах цитоплазмы, где появляется необходимость в АТФ. Например, в мышечной ткани сердца они располагаются неподалеку от миофибрилл, а в сперматозоидах формируют защитную маскировку вокруг оси жгута. Там они вырабатывают очень много энергии для того, чтобы «хвост» крутился. Именно таким образом сперматозоид двигается к яйцеклетке.
В клетках новые митохондрии образуются с помощью простого деления предыдущих органелл. Во время него сохраняется вся наследственная информация.
Митохондрии: как они выглядят
По форме митохондрии напоминает цилиндр. Они часто встречаются в эукариотах, занимая от 10 до 21 % объема клетки. Их размеры и формы во многом разнятся и способны меняться в зависимости от условий, но ширина постоянна: 0,5-1 мкм. Перемещения хондриосом зависят от того, в каких местах клетки совершается быстрая трата энергии. Передвигаются по цитоплазме, применяя для передвижения структуры цитоскелета.Заменой разных по габаритам митохондрий, работающих отдельно друг от друга и снабжающих энергией некоторые зоны цитоплазмы, являются длинные и разветвленные митохондрии. Они способны обеспечить энергией участки клеток, находящиеся далеко друг от друга. Подобная совместная работа хондриосом наблюдается не только у одноклеточных организмов, но и у многоклеточных. Самое сложное строение хондриосом встречается в мышцах скелета млекопитающих, где самые большие разветвленные хондриосомы стыкуются друг с другом, используя межмитохондриальные контакты (ММК).
Они представляют собой узкие просветы между прилегающими друг к другу митохондриальными мембранами. Данное пространство обладает высокой электронной плотностью. ММК больше встречаются в клетках сердечной мышцы, где связываются вместе с работающими хондриосомами.
Чтобы лучше разобраться в вопросе, нужно кратко расписать значимость митохондрии, строение и функции этих удивительных органелл.
Как они устроены?
Для понимания, что такое митохондрии, необходимо узнать их строение. Этот необычный источник энергии имеет форму шара, но чаще вытянут. Две мембраны располагаются близко друг к другу:
- наружная (гладкая);
- внутренняя, которая образует выросты листовидной (кристы) и трубчатой (тубулы) формы.
Если не принимать во внимание размер и форму митохондрии, строение и функции у них одинаковые. Хондриосома разграничена двумя мембранами, размером 6 нм. Наружная мембрана митохондрии напоминает емкость, которая ограждает их от гиалоплазмы. Внутреннюю мембрану от внешней отъединяет участок шириной 11-19 нм. Отличающей чертой внутренней мембраны считается ее способность выпячиваться внутрь митохондрии, принимая форму сплющенных гребней.Внутреннюю полость митохондрии заполняет матрикс, который имеет мелкозернистую структуру, где иногда обнаруживают нити и гранулы (15-20 нм). Нити матрикса создают молекулы ДНК органеллы, а гранулы небольших размеров – рибосомы митохондрии.
Синтез АТФ на первой стадии проходит в гиалоплазме. На данной ступени идет начальное окисление субстратов или глюкозы до пировиноградной кислоты. Данные процедуры проходят без кислорода — анаэробное окисление. Следующая стадия образования энергии заключается в аэробном окислении и распада АТФ, данный процесс происходит в митохондриях клеток.
Что делают митохондрии?
Основными функциями этой органеллы являются:
-
выработка энергии для клеток;
-
хранение наследственной информации в виде собственной ДНК.

Наличие в митохондриях своей дезоксирибонуклеиновой кислоты еще раз подтверждает симбиотическую теорию появления этих органелл. Также, помимо основной работы, они участвуют в синтезе гормонов и аминокислот.
Митохондриальная патология
Мутации, происходящие в геноме митохондрии, приводят к удручающим последствиям. Носителем наследственной информации человека является ДНК, которая передается потомкам от родителей, а митохондриальный геном передается только от матери. Объясняется данный факт очень просто: цитоплазму с заключенными в ней хондриосомами дети получают вместе с женской яйцеклеткой, в сперматозоидах они отсутствуют. Женщины с данным отклонением могут передать потомству митохондриальное заболевание, больной мужчина – нет.
В обычных условиях хондриосомы располагают одинаковой копией ДНК — гомоплазмия. В геноме митохондрии могут происходить мутации, вследствие совместного существования здоровых и мутированных клеток возникает гетероплазмия.
Благодаря современной медицине на сегодняшний день выявлены более 200 заболеваний, поводом возникновения чего послужила мутация митохондрии ДНК. Не во всех случаях, но терапевтическому поддержанию и лечению митохондриальные болезни поддаются хорошо.
Вот мы и разобрались с вопросом о том, что такое митохондрии. Как и все остальные органеллы, они очень важны для клетки. Они косвенно принимают участие во всех процессах, для которых нужна энергия.
Источник: fb.ru
- Друкувати
- Запитати
- Надіслати другу
- Поділитись
- Google+
- Підписатись на новини
Система синтезу білка в мітохондріях. Продукти мітохондріального білкового синтезу. Синтез мітохондріальних білків у цитоплазмі. Транспорт у мітохондрії білків, синтезованих у цитоплазмі. Формування окремих компонентів мембран мітохондрій
Роль мітохондрії у розвитку хребетних переоцінити важко. Адже всі процеси в клітині, так чи інакше пов’язані з енергетичним обміном, залежать від клітинного дихання, важливою складовою якого є окислювальне фосфорилювання. Останнє не можна уявити без мітохондріальних мембран. У процесі онтогенезу роль окислювального фосфорилювання, а отже і мітохондрій, проявляється особливо яскраво і відкриває цілий ряд питань, що потребують дослідження.
По-перше, на даний час ведеться жвава наукова дискусія щодо впливу енергетичного обміну на вік клітини. Відомо, що при розвитку і старінні організму сильно змінюється поглинання кисню клітиною, і це не може не відображатися на структурі та кількості мітохондрій. Тобто, вивчаючи процеси формування мітохондрій у розвитку, ми зможемо впливати на процеси клітинного старіння та омолоджування.
По-друге, широкі перспективи відкриває дослідження мітохондрії як носія власного геному. Геном мітохондрії має значно менші розміри, ніж ядерний геном, і тому є зручним об’єктом для вивчення. Крім цього, відкритим залишається питання про взаємодію мітохондріального та ядерного геномів, про перенесення генів з мітохондрії в ядро у процесі еволюційного розвитку. Дослідження цих проблем може допомогти у вирішенні багатьох питань еволюції і генетики.
По-третє, досі незрозумілим є процес відтворення мітохондрій в оогенезі, і це питання також активно досліджується.
Синтез мітохондріальних білків і особливості формування мітохондрій
Формування мітохондрій є багатоступінчатим і багатокомпонентним процесом Особливістю їх утворення є те, що мітохондріальні білки є продуктами діяльності двох генетичних систем: мітохондріальної та ядерної. Просторове розділення цих двох генетичних систем є одною із причин складності їх системи взаєморегуляції як на рівні синтезу окремих білків, так і на рівні інтактної мітохондрії. Розглянемо процеси, що в сукупності утворюють єдиний механізм формування мітохондрій
Система синтезу білків в мітохондрія
Апарат білкового синтезу, локалізований у мітохондріях, призначення для утворення дуже малої кількості білків, що становить не більше 7 -10% білків внутрішньої мембрани мітохондрій. Основним продуктом мітохондріального білкового синтезу є поліпептиди з сильно вираженими гідрофобними властивостями [Пинус, Рабинович, 1977].
Апарат трансляції мітохондрій характеризується рядом унікальних властивостей. Перш за все, це стосується розміру рибосом. Мітохондріальні рибосоми тварин — найменші серед усіх відомих типів рибосом, що зумовлено відносно невеликими розмірами рРНК, відсутністю 5S рРНК, малим розміром рибосомальних білків [Минченко, 1987].
У порівнянні з цитоплазматичними та бактеріальними мітохондріальні рибосоми тварин характеризуються більш низьким співвідношенням РНК та білка. В клітинах жаби у великій субчастинці мітохондріальних рибосом вміст РНК становить 32%, в малій субчастинці — 20%, тоді як в цитоплазматичних рибосомах ця величина дорівнює 58% РНК для великої субчастинки і 50% РНК для малої субчастинки. Для мітохондрій ссавців встановлено, що вміст РНК у їхніх рибосомах приблизно вдвічі нижчий в порівнянні з іншими рибосомами [Озернюк, 1978].
Рибосоми тварин містять не менш ніж 87 білків: 52 білки у великій субодиниці (молекулярна маса — 8,8 — 49 тис. дальтон) і 33 білки у меншій субодиниці (молекулярна маса — 10 — 48 тис. дальтон), причому всі вони різняться за електрофоретичною рухливістю від білків цитоплазматичних рибосом. Унікальними є і мітохондріальні мРНК. Вони не містять довгих фланкіруючих ділянок, що не транслюються, які присутні в мРНК з цитоплазми, і відповідальні за зв’язування з рибосомами, характеризуються вкороченими полі-А-послідовностями та відсутністю "кепів", необхідних для ефективної трансляції цитоплазматичних мРНК [Гаузе, 1977].
Що стосується мітохондріальної ДНК, то відомо, що вона успадковується лише по материнській лінії. Сцолозі [Szollozi, 1965; цит. за: Билл, Ноулз, 1981] виявив, що у щурів при заплідненні мітохондрії сперміїв входять в яйцеклітину, а потім набухають і руйнуються. Існують докази того, що після запліднення мітохондріальна ДНК елімінується [Hutchison et al. 1974, цит. за Билл, Ноулз, 1981]. Висловлено припущення, що по батьківській лінії може передаватися не більш ніж одна молекула ДНК мітохондрій на 25 тисяч материнських молекул. [Минченко, Дударева, 1990]
У ссавців мітохондріальний геном представлений кільцевою мітохондріальною ДНК розміром близько 5 мкм (молекулярна маса приблизно 10 МД), що присутня в мітохондріях у вигляді ковалентно замкнутих форм, що перебувають у надспіралізованому стані [Минченко, 1987].
Комплементарні ланцюги в мітохондріальної ДНК більшості тварин суттєво різняться за плавучою щільністю в градієнтах лужного хлориду цезію, оскільки мають неоднаковий середній нуклеотидний склад, що коливається у тварин різних видів у значних межах. Частина молекул ДНК в мітохондріях клітин більшості організмів присутня у вигляді олігомерних форм, що поділяються на два класи: кільцеві олігомери, тобто молекули з контурною довжиною, кратною довжині мономерних кілець і ланцюгові олігомери (катенани) що складаються із зв’язаних топологічним зв’язком мономерних кілець [Минченко, Дударева, 1990].
Особливістю мітохондріальної системи трансляції є також те, що синтез білків в мітохондріях людини і тварин здійснюється за участю лише 22 — 23 тРНК. Цієї кількості тРНК достатньо завдяки тому, що система генетичного коду мітохондрій змінена в порівнянні із універсальним генетичним кодом. (табл. 1). Більшість мітохондріальних тРНК здатні зчитувати по чотири кодони, які різняться за третім нуклеотидом, тобто при наявності чотирьох варіантів кодування однієї амінокислоти. Слід також відмітити, що розмір мітохондріальних тРНК клітин тварин менший у порівнянні з цитоплазматичними тРНК. [Anderson et al., 1981].
Табл. 1 Генетичний код мітохондрій [Anderson et al., 1981].
|
Phe |
UUU |
77 |
Ser |
UCU |
32 |
Tyr |
UAU |
46 |
Cys |
UGU |
5 |
|
|
UUC |
141 |
|
UCC |
99 |
|
UAC |
89 |
|
UGC |
17 |
|
Leu |
UUA |
73 |
|
UCA |
83 |
Ter |
UAA |
— |
Trp |
UGA |
93 |
|
|
UUG |
16 |
|
UCG |
7 |
|
AAG |
— |
|
UGG |
11 |
|
Leu |
CUU |
65 |
Pro |
CCU |
41 |
His |
CAU |
18 |
Arg |
CGU |
7 |
|
|
CUC |
167 |
|
CCC |
119 |
|
CAC |
79 |
|
CGC |
25 |
|
|
CUA |
276 |
|
CCA |
52 |
Gln |
CAA |
81 |
|
CGA |
29 |
|
|
CUG |
45 |
|
CCG |
7 |
|
CAG |
9 |
|
CGG |
2 |
|
Ile |
AUU |
125 |
Thr |
ACU |
51 |
Asn |
AAU |
33 |
Ser |
AGU |
14 |
|
|
AUC |
196 |
|
ACC |
155 |
|
AAC |
131 |
|
AGC |
39 |
|
Met |
AUA |
167 |
|
ACA |
133 |
Lys |
AAA |
85 |
Ter |
AGA |
— |
|
|
AUG |
40 |
|
ACG |
10 |
|
AAG |
10 |
|
AGG |
— |
|
Val |
GUU |
30 |
Ala |
GCU |
43 |
Asp |
GAU |
15 |
Gry |
CGU |
24 |
|
|
GUC |
49 |
|
GCC |
124 |
|
GAC |
51 |
|
CGC |
88 |
|
|
GUA |
70 |
|
GCA |
80 |
Glu |
GAA |
64 |
|
GGA |
67 |
|
|
GUG |
18 |
|
GCG |
8 |
|
GAG |
24 |
|
GGG |
34 |
У таблиці показано бокси, що зчитуються за допомогою єдиної тРНК. Вказано також загальну кількість амінокислотних кодонів, знайдених у генах та послідовностях, які можуть бути генами ДНК мітохондрій.
Ще однією особливістю синтезу мітохондріальних білків є те, що він стійкий до дії специфічного інгібітора цитоплазматичної трансляції циклогексиміду і специфічно пригнічується хлорамфеніколом і бромідом етидію. Синтез білків в ізольованих мітохондріях протікає з низькою швидкістю і нелінійно. Це, можливо, зумовлено затримкою просування полірибосоми по мРНК в її середній ділянці і частково присутністю в мітохондріях інгібітора трансляції, який знижує загальну кількість полірибосом шляхом порушення цілісності мРНК [Шугалий и др., 1974].
Продукти мітохондріального білкового синтезу
Мітохондріальний геном тварин кодує синтез відносно невеликої кількості поліпептидів. В мітохондріальній ДНК клітин людини та тварин ідентифіковано 13 рамок зчитування для п’яти відомих і восьми ще не вивчених білків. В генах, що кодують синтез мітохондріальних білків у клітинах тварин інтрони не виявлені, на відміну від клітин вищих рослин та грибів [Anderson et al, 1981].
Цікавим є той факт, що кількість білків — продуктів мітохондріальної трансляції, виявлених за допомогою електрофорезу, значно перевищує кількість рамок зчитування, а отже і кількість потенціальних генів, ідентифікованих в Н-тяжі мітохондріальної ДНК. Це може пояснюватися тим, що серед білків, які синтезуються в мітохондрії, є такі, що кодуються L — тяжем мітохондріальної ДНК, хоча на ньому рамки зчитування ще не виявлені. Крім цього, можна припустити, що деякі з ідентифікованих рамок зчитування можуть кодувати синтез більш ніж одного білка.
На даний час серед мітохондріальних продуктів трансляції ідентифіковані три субодиниці (І, ІІ і ІІІ) цитохромоксидази, апоцитохром b і дві субодиниці АТФ-ази, причому гени, що кодують синтез обох субодиниць (6-ї та 8-ї) виявлені лише в геномі мітохондрій дріжджів, а в клітинах тварин однозначно виявлений лише ген 6-ї субодиниці. Детально вивчено функціональне значення цілого ряду білків, які синтезуються в мітохондріях. Існують дані, що деякі із синтезованих в мітохондріях поліпептидів можуть виходити з мітохондрій і грати певну роль в ядерно-мітохондріальних взаємовідношеннях, зокрема в регуляції експресії ядерних генів, що кодують синтез мітохондріальних білків [Bibb et al., 1981; цит. за: Минченко, 1987].
Доказом цитоплазматичного походження основної маси мітохондріальних білків є такі ознаки:
- Мітохондрії цитоплазматичних мутантів petite дріжджів мають дефективний дихальний ланцюг (відсутні цитохроми аа3, В і С), в них редукована або повністю відсутня мітохондріальна ДНК, а також відсутня, очевидно, система синтезу білка. Отже, всі білки, присутні у таких дефектних мітохондріях, мають цитоплазматичне походження;
- Мітохондрії клітин, що ростуть у присутності інгібіторів мітохондріального білкового синтезу (хлорамфеніколу або еритроміцину), мають дефектну дихальну систему. При повному інгібуванні білкового синтезу в мітохондріях всі білки, що залишилися в органелі, мають цитоплазматичне походження. Природно, що мітохондрії клітин, які ростуть в присутності інгібіторів синтезу мітохондріальних білків, за своєю будовою та набором ферментів схожі на мітохондрії цитоплазматичних мутантів petite дріжджів [Озернюк, 1985].
Отже, значення мітохондріального білкового синтезу полягає в забезпеченні мітохондрій невеликою кількістю гідрофобних білків, які входять у склад ряду ферментних комплексів, локалізованих на внутрішній мембрані мітохондрій, а також у синтезі поліпептидів, які беруть участь в регуляції ядерно-цитоплазматичних взаємовідношеннях.
Синтез мітохондріальних білків у цитоплазмі
Мітохондрії містять у зовнішніх та внутрішніх мембранах, міжмембранному просторі і матриксі кілька сотень білків, синтез яких протікає на матрицях, що кодуються ядерним геномом. Для багатьох мітохондріальних ферментів відомі функціонуючі в цитоплазмі аналоги, але цитоплазматичні та мітохондріальні форми цих білків кодуються різними генами. Синтез цих білків відбувається як на вільних, так і на зв’язаних із мембранами ендоплазматичного ретикулуму цитоплазматичних полісомах. [Сэджер, 1975].
На даний час детально вивчено синтез великої кількості мітохондріальних білків зовнішніх та внутрішніх мембран, міжмембранного простору та матриксу і їх транспорт в мітохондрії. Всі вивчені білки зовнішніх мембран мітохондрій синтезуються в цитоплазмі і в такому вигляді вбудовуються у мембрану, а всі інші білки (принаймні, частина з них) синтезуються у вигляді попередників з N-кінцевою лідерною послідовністю розміром від 1 до 6 тис. дальтон. Білки чи їх попередники, що поступають з цитоплазми, розпізнаються певними структурами (рецепторами), локалізованими на зовнішній мембрані мітохондрій.
У білків-попередників в N-кінцевій лідерній послідовності міститься не лише інформація, необхідна для розпізнавання рецептором, а й інформація, що забезпечує подальший транспорт білка і окремі стадії його процесингу [Adoutte, 1983, Benson et al., 1984; цит. за:Минченко, 1987].
Транспорт у мітохондрії білків, синтезованих у цитоплазмі
Очевидно, існує не менш ніж три механізми транспорту білків та їх попередників у мітохондрії. Перший механізм характерний для білків зовнішніх мембран мітохондрій, які синтезуються на цитоплазматичних полісомах у вигляді зрілих форм. Ці білки мають сигнальні послідовності, за допомогою яких впізнаються певними структурами зовнішньої мембрани і вбудовуються в неї.
Другий та третій механізми транспорту характерні для тих мітохондріальних білків, які синтезуються в цитоплазмі у вигляді попередників. Ці білки своїми розпізнавальними структурами взаємодіють з певними рецепторами на зовнішній мембрані і проникають крізь неї. Білки, що транспортуються за другим механізмом, проходять через внутрішню мембрану за рахунок процесу, який вимагає наявності електрохімічного градієнта на внутрішній мембрані; потім відбувається відщеплення лідерної послідовності.
Транспорт таких білків не спряжений облігатно з процесингом, оскільки попередники білків ефективно накопичуються всередині мітохондрій (матриксі чи внутрішніх мембранах) навіть при сильному інгібуванні мітохондріальних протеїназ. За третім механізмом надходять білки з цитоплазми в міжмембранний простір. Ці білки транслокуються крізь зовнішню мембрану мітохондрій, а потім N-кінцева частина попередника білка проходить через внутрішню мембрану в матрикс, де за допомогою матриксних протеїназ відбувається відщеплення частини лідерної послідовності.
Після цього утворена форма попередника повертається у міжмембранний простір, де зв’язується із зовнішньою поверхнею внутрішньої мембрани, і лише після цього відбувається завершальний процесинг під дією зв’язаної з внутрішньою мембраною протеїнази. Транспортовані в мітохондрії поліпептиди зазнають посттрансляційної модифікації. В мітохондріях виявлені протеїнкінази, які фосфорилюють білки зовнішньої та внутрішньої мембран мітохондрій [Минченко, Дударева, 1990].
Формування окремих компонентів мембран мітохондрій. Розмноження мітохондрій
Збирання компонентів мітохондрій, що синтезуються у різних частинах клітини, є досить складним процесом, що регулюється як на рівні взаємної координації мітохондріального та цитоплазматичного білкового синтезу, так і на рівні властивостей самих компонентів. Вважають, що гідрофобні продукти синтезу білка в мітохондріях зв’язуються з ліпідами. Утворені протеоліпіди здатні до взаємної агрегації і формування більш крупних комплексів, які можуть бути "матрицями" для зв’язування білків, які синтезуються у цитоплазмі. Такий висновок для хребетних вперше був зроблений у роботах Дроза та Бержерона [Droz, Bergeron, 1965; цит. за: Озернюк, 1978].
При вивченні особливостей ультраструктури та властивостей внутрішньої мембрани мітохондрій було зроблено висновок, що в мітохондріях існують зони росту, в яких переважно і відбувається вбудовування нових компонентів. Зони росту, на думку Вернера та Нейперта, розташовані в ділянках, де внутрішня мембрана переходить у кристи. [Werner, Neupert, 1972; цит. за: Озернюк, 1978].
Наскільки складним є питання про механізм розмноження мітохондрій, свідчить той факт, що досі не зрозуміло, як відбувається процес збільшення кількості мітохондрій. На думку Озернюка [1978], мітохондрії розмножуються шляхом поділу. Проте в роботах останніх років автори притримуються думки, що мітохондрії виникають в результаті формування мітохондріальних мембран на структурах-попередниках[Лузиков, 1980; цит. за: Зотин, Зотина, 1993].
Оскільки у багатьох випадках мітохондрії здатні утворювати неперервний мітохондріальний ретикулум, або ж виникають гігантські мітохондрії [Айзенштадт, 1984], то не виключено, що остання точка зору більше відповідає реальності. Все це показує, що висловлювати які-небудь припущення щодо механізму збільшення мітохондрій у період малого росту ооцитів поки що рано.
Література
1. Айзенштадт Т. В. Цитология оогенеза — М., Наука, 1993, 364с.
2. Билл Дж., Ноулз Дж. Внеядерная наследственность. М., Мир, 1981, 168с.
3. Бурнашева С. А., Габаева Н. С. и др. Современные проблемы сперматогенеза. М., Наука, 1982, 259с.
4. Гаузе Г. Г. Митохондриальная ДНК. М., Наука, 1977, 286с.
5. Данилова Л. В. Ультраструктурные исследования сперматогенеза. М., Наука, 1978, 206с.
6. Зотин А. И., Зотина Р. С. Феноменологическая теория развития, роста и старения организма. М., Наука, 1993, 364с.
7. Кауфман З. С. Эмбриология рыб. М., Агропромиздат, 1990, 272с.
8. Минченко А. Г., Дударева Н. А. Митохондриальный геном. Новосибирск, Наука, 1990, 194с.
9. Озернюк Н. Д. Рост и воспроизведение митохондрий. М., Наука, 1978, 264с.
10. Озернюк Н. Д. Энергетический обмен в раннем эмбриогенезе рыб. М., Наука,!985, 196с.
11. Сэджер Р. Цитоплазматические гены и органеллы. М., Мир, 1975, 424с.
Источник: osvita.ua