За что отвечают хлоропласты
Строение хлоропластов
Данный органоид присутствует только у растений. Хлоропласты имеют форму двояковыпуклой линзы, в результате чего на листья поступает больше света. Покрыты наружной мембраной. Это мембрана гладкая, по сравнению с внутренней. Внутри находятся тилокоиды.
Благодаря дисковидным тилокоидам образуются граны, которые различимы только под микроскопом, а благодаря трубковидным тилокоидам образуется строма, которая соединяет все образовавшиеся граны в одну систему. Количество гран в хлоропластах составляет приблизительно 40-60 единиц. Граны объединяются между собой с помощью межгранных тяжей.
В строме содержится ДНК, рибосомы, РНК. В мембране тилокоид содержится вещество, от которого зависит цвет листьев. Хлорофилл (зелёный) и каротиноиды (красный, оранжевый, желтый).
Именно благодаря хлорофиллу в клетках растений осуществляется процесс фотосинтеза.
Существует 4 вида холорофилла, в зависимости от строения: a, b, c и d. Тип а и б содержат все растения на суше и зеленые водоросли. А и С- диатомовые водоросли, а и d — красные.
Функции хлоропластов
В хлоропластах происходит фотосинтез — процесс преобразования солнечной энергии в кислород. Хлоропласты способны перемещаться в цитоплазме клеток. За счет этого молекулы хлорофилла получают максимальное количество солнечной энергии для осуществления функции фотосинтеза.
Фотосинтез является основным процессом, вследствие которого на нашей планете образуется кислород и органические вещества.
Без фотосинтеза не было бы растений и кислорода, а без них и животных, в том числе невозможно было бы существование человека.
Еще одной функцией хлоропластов является фиксация углекислоты и встраивание углерода в состав органических веществ. Такой процесс называется реакция Кальвина-Бенсона, в честь ученых, открывших ее.
В конце жизненного цикла органоида, хлорофилл начинает разрушаться, функции растительных клеток нарушаются. Это также может происходить из-за изменения светового дня и резкого понижения температуры окружающей среды. Часть хлоропластов становятся хромопластами — зеленые листья изменяют цвет, и вскоре опадают.
Источник: appteka.ru
Строение хлоропласта
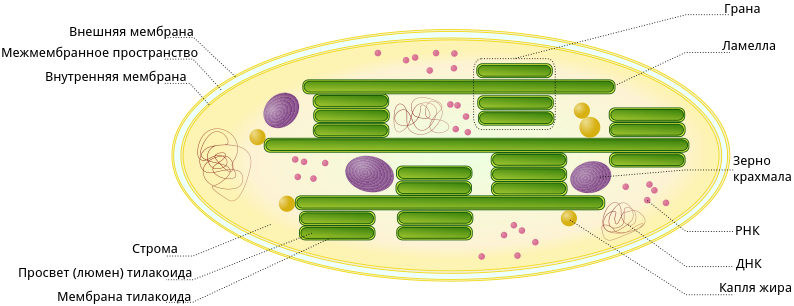
В строении хлоропластов выделяют внешнюю и внутреннюю мембраны, межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.

Тилакоид представляет собой ограниченное мембраной пространство в форме приплюснутого диска. Тилакоиды в хлоропластах объединяются в стопки, которые называют гранами. Граны связаны между собой удлиненными тилакоидами — ламеллами.
Полужидкое содержимое хлоропласта называется стромой. В ней находятся его ДНК и РНК, рибосомы, обеспечивающие полуавтономность органоида (см. Симбиогенез).
Также в строме находятся зерна крахмала. Они образуются при избытке углеводов, образовавшихся при фотосинтетической активности. Жировые капли обычно формируются из мембран разрушающихся тилакоидов.
Функции хлоропластов
Основная функция хлоропластов — это фотосинтез — синтез глюкозы из углекислого газа и воды за счет солнечной энергии, которая улавливается хлорофиллом. В качестве побочного продукта фотосинтеза выделяется кислород. Однако процесс этот сложный и многоступенчатый, при котором синтезируются и побочные продукты, использующиеся как в самом хлоропласте, так и в остальных частях клетки.
Основным фотосинтетическим пигментом является хлорофилл. Он существует в нескольких разных формах. Кроме хлорофилла в фотосинтезе принимают участие пигменты каротиноиды.
Пигменты локализованы в мембранах тилакоидов, здесь протекают световые реакции фотосинтеза. Кроме пигментов здесь присутствуют ферменты и переносчики электронов. Хлоропласты стараются расположиться в клетке так, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету.
Хлорофилл состоит из длинного углеводного кольца и порфириновой головки. Хвост гидрофобен и погружен в липидный слой мембран тилакоидов. Головка гидрофильна и обращена к строме. Энергия света поглощается именно головкой, что приводит к возбуждению электронов.
Электрон отделяется от молекулы хлорофилла, который после этого становится электроположительным, т. е. оказывается в окисленной форме. Электрон принимается переносчиком, которые передает его на другое вещество.
Разные виды хлорофилла отличаются между собой несколько различным спектром поглощения солнечного света. Больше всего в растениях хлорофилла А.
В строме хлоропласта происходят темновые реакции фотосинтеза. Здесь находятся ферменты цикла Кальвина и другие.
Источник: biology.su
Происхождение
В настоящее время общепризнано[2] происхождение хлоропластов путём симбиогенеза. Предполагают, что хлоропласты возникли из цианобактерий, так как являются двухмембранным органоидом, имеют собственную замкнутую кольцевую ДНК и РНК, полноценный аппарат синтеза белка (причем рибосомы прокариотического типа — 70S), размножаются бинарным делением, а мембраны тилакоидов похожи на мембраны прокариот (наличием кислых липидов) и напоминают соответствующие органеллы у цианобактерий.
глаукофитовых водорослей вместо типичных хлоропластов в клетках содержатся цианеллы — цианобактерии, потерявшие в результате эндосимбиоза способность к самостоятельному существованию, но отчасти сохранившие цианобактериальную клеточную стенку[3].
Давность этого события оценивают в 1 — 1,5 млрд лет[4].
Часть групп организмов получала хлоропласты в результате эндосимбиоза не с прокариотными клетками, а с другими эукариотами, уже имеющими хлоропласты[5]. Этим объясняется наличие в оболочке хлоропластов некоторых организмов более чем двух мембран[Пр. 2]. Самая внутренняя из этих мембран трактуется как потерявшая клеточную стенку оболочка цианобактерии, внешняя — как стенка симбионтофорной вакуоли хозяина. Промежуточные мембраны — принадлежат вошедшему в симбиоз редуцированному эукариотному организму. У некоторых[Пр. 3] групп в перипластидном пространстве между второй и третьей мембраной располагается нуклеоморф, сильно редуцированное эукариотное ядро[6].
Строение
| Этот раздел не завершён. |
У различных групп организмов хлоропласты значительно различаются по размерам,строению и количеству в клетке. Особенности строения хлоропластов имеют большое таксономическое значение[7]. В основном хлоропласты имеют форму двояковыпуклой линзы, размер их около 4-6 мкм.
Оболочка хлоропластов
У различных групп организмов оболочка хлоропластов отличается по строению.
У глаукоцистофитовых, красных, зелёных водорослей[8] и у высших растений оболочка состоит из двух мембран. У других эукариотных водорослей хлоропласт дополнительно окружён одной или двумя мембранами. У водорослей, обладающих четырёхмембранными хлоропластами, наружная мембрана обычно переходит в наружную мембрану ядра.
Перипластидное пространство
Ламеллы и тилакоиды
Ламеллы соединяют полости тилакоидов
Пиреноиды
Пиреноиды — центры синтеза полисахаридов в хлоропластах[9]. Строение пиреноидов разнообразно, и не всегда они морфологически выражены. Могут быть внутрипластидными и стебельчатыми, выступающими в цитоплазму. У зелёных водорослей и растений пиреноиды располагаются внутри хлоропласта, что связано с внутрипластидным запасанием крахмала.
Стигма
Стигмы или глазки встречаются в хлоропластах подвижных клеток водорослей. Располагаются вблизи основания жгутика. Стигмы содержат каротиноиды и способны работать как фоторецепторы[10].
См. также
- Фотосинтез
- Триозофосфатный транслокатор
- Хромопласты
- Цианеллы
Примечания
Комментарии
- ↑ Хлоропласты организмов, относящихся к группе хромистов, имеют четырёхслойную оболочку. Предполагается, что в истории их возникновения включение одной клетки в состав другой происходило дважды.
- ↑ Например, у динофитовых и эвгленовых имеется 3 мембраны, а у охрофитов — 4.
- ↑ У криптофитовых, хлорарахниофитовых и некоторых динофитовых.
Литература
- Белякова Г. А. Водоросли и грибы // Ботаника: в 4 т. / Белякова Г. А., Дьяков Ю. Т., Тарасов К. Л. — М.: Издательский центр «Академия», 2006. — Т. 1. — 320 с. — 3000 экз. — ISBN 5-7695-2731-5.
- Карпов С.А. Строение клетки протистов. — СПб.: ТЕССА, 2001. — 384 с. — 1000 экз. — ISBN 5-94086-010-9.
- Lee, R. E. Phycology, 4th edition. — Cambridge: Cambridge University Press, 2008. — 547 с. — ISBN 9780521682770.
|
||||||||||||||||||||||||||||
|
||||||||||||||||||||||
Отрывок, характеризующий Хлоропласты
– Нет. Она живёт там, куда ни я, ни ты не можем пойти. Её земная жизнь здесь с нами, кончилась, и она теперь живёт в другом, очень красивом мире, из которого может тебя наблюдать. Но она видит, как ты страдаешь, и не может отсюда уйти. А здесь она уже находиться дольше тоже не может. Поэтому ей нужна твоя помощь. Ты хотела бы ей помочь?
– А откуда ты всё это знаешь? Почему она разговаривает с тобой?!.
Я чувствовала, что пока ещё она мне не верит и не хочет признавать во мне друга. И я никак не могла придумать, как же объяснить этой маленькой, нахохлившейся, несчастной девчушке, что существует «другой», далёкий мир, из которого, к сожалению, нет возврата сюда. И что её любимая мама говорит со мной не потому, что у неё есть выбор, а потому, что мне просто «посчастливилось» быть немножечко «другой», чем все остальные…
– Все люди разные, Алинушка, – начала я. – Одни имеют талант к рисованию, другие к пению, а вот у меня такой особый талант к разговору с теми, которые ушли из нашего с тобой мира уже навсегда. И твоя мама говорит со мной совсем не потому, что я ей нравлюсь, а потому, что я её услышала, когда больше никто её услышать не мог. И я очень рада, что хоть в чём-то могу ей помочь. Она тебя очень любит и очень страдает оттого, что ей пришлось уйти… Ей очень больно тебя оставлять, но это не её выбор. Ты помнишь, она тяжело и долго болела? – девочка кивнула. – Вот эта болезнь и заставила её покинуть вас. А теперь она должна уйти в свой новый мир, в котором она будет жить. И для этого она должна быть уверена, что ты знаешь, как она тебя любит.
Девочка грустно на меня посмотрела и тихо спросила:
– Она живёт теперь с ангелами?.. Папа мне говорил, что она теперь живёт в таком месте, где всё, как на открытках, что мне дарят на рождество. И там такие красивые крылатые ангелы… Почему она не взяла меня с собой?..
– Потому, что ты должна прожить свою жизнь здесь, милая, а потом ты тоже пойдёшь в тот же мир, где сейчас твоя мама.
Девочка засияла.
– Значит, там я её увижу? – радостно пролепетала она.
– Конечно, Алинушка. Поэтому ты должна быть всего лишь терпеливой девочкой и помочь твоей маме сейчас, если ты её так сильно любишь.
– Что я должна делать? – очень серьёзно спросила малышка.
– Всего лишь думать о ней и помнить её, потому, что она видит тебя. И если ты не будешь грустить, твоя мама наконец-то обретёт покой.
– Она и теперь видит меня?– спросила девочка и её губки начали предательски дёргаться.
– Да милая.
Она на какой-то миг замолчала, как бы собираясь внутри, а потом крепко сжала кулачки и тихо прошептала:
– Я буду очень хорошей, милая мамочка… ты иди… иди пожалуйста… Я тебя так люблю!..
Слёзы большими горошинами катились по её бледным щёчкам, но лицо было очень серьёзным и сосредоточенным… Жизнь впервые наносила ей свой жестокий удар и, казалось, будто эта маленькая, так глубоко раненная, девчушка вдруг совершенно по-взрослому что-то для себя осознала и теперь пыталась серьёзно и открыто это принять. Моё сердце разрывалось от жалости к этим двум несчастным и таким милым существам, но я, к сожалению, ничем больше не могла им помочь… Окружающий их мир был таким невероятно светлым и красивым, но для обоих это уже не мог больше быть их общий мир…
Жизнь порой бывает очень жестокой, и мы никогда не знаем, в чём заключается смысл приготовленной нам боли или потери. Видимо, это правда, что без потерь невозможно осмыслить того, что по праву или по счастливой случайности, дарит нам судьба. Только вот, что же могла осмыслить эта несчастная, съёжившаяся, как раненный зверёк, девчушка, когда мир вдруг обрушился на неё всей своей жестокостью и болью самой страшной в жизни потери?..
Я ещё долго сидела с ними и старалась, как могла, помочь им обеим обрести хоть какой-то душевный покой. Я вспомнила своего дедушку и ту жуткую боль, которую принесла мне его смерть… Как же должно было быть страшно этой хрупкой, ничем не защищённой малышке потерять самое дорогое на свете – свою мать?..
Мы никогда не задумываемся о том, что те, которых по той или иной причине отнимает у нас судьба, переживают намного глубже нас последствия своей смерти. Мы чувствуем боль потери и страдаем (иногда даже злясь), что они так безжалостно нас покинули. Но, каково же им, когда их страдание умножается в тысячи раз, видя то, как страдаем от этого мы?!. И каким беспомощным должен себя чувствовать человек, не имея возможности ничего больше сказать и ничего изменить?..
Я бы многое тогда отдала, чтобы найти хоть какую-то возможность предупредить об этом людей. Но, к сожалению, у меня таковой возможности не было… Поэтому, после печального визита Вероники, я стала с нетерпением ждать, когда же ещё кому-то смогу помочь. И жизнь, как это всегда обычно бывало, не заставила себя долго ждать.
Сущности приходили ко мне днём и ночью, молодые и старые, мужские и женские, и все просили помочь им говорить с их дочерью, сыном, мужем, женой, отцом, матерью, сестрой… Это продолжалось нескончаемым потоком, пока, под конец, я не почувствовала, что у меня нет больше сил. Я не знала, что, входя с ними в контакт, я должна была обязательно закрываться своей (к тому же, очень сильной!) защитой, а не открываться эмоционально, как водопад, постепенно отдавая им всю свою жизненную силу, которую тогда ещё, к сожалению, я не знала, как восполнять.
Очень скоро я буквально не имела сил двигаться и слегла в постель… Когда мама пригласила нашего врача, Дану, проверить, что же такое снова со мной стряслось, та сказала, что это у меня «временная потеря сил от физического переутомления»… Я не сказала никому ничего, хотя прекрасно знала настоящую причину этого «переутомления». И как делала уже давно, просто честно глотала любое лекарство, которое прописала мне моя двоюродная сестра, и, отлежавшись в постели около недели, опять была готова на свои очередные «подвиги»…
Я давно поняла, что искренние попытки объяснений того, что по-настоящему со мной происходило, не давали мне ничего, кроме головной боли и усиления постоянного наблюдения за мной моих бабушки и мамы. А в этом, честно говоря, я не находила никакого удовольствия…
Источник: o-ili-v.ru
(от греч. chlorós ≈ зелёный и plastós ≈ вылепленный, образованный), внутриклеточные органеллы растительной клетки ≈ пластиды , в которых осуществляется фотосинтез. Окрашены в зелёный цвет благодаря присутствию в них основного пигмента фотосинтеза ≈ хлорофилла . Основная функция Х., состоящая в улавливании и преобразовании световой энергии, нашла отражение и в особенностях их строения. У высших растений Х. ≈ тельца линзообразной формы диаметром 3≈10 мкм и толщиной 2≈5 мкм, представляют собой систему белково-липидных мембран, погруженных в основное вещество ≈ матрикс, или строму, и отграничены от цитоплазмы наружной мембраной (оболочкой). Внутренние мембраны образуют единую (непрерывную) пластинчатую, или ламеллярную, систему, состоящую из замкнутых уплощённых мешочков (цистерн) ≈ т. н. тилакоидов, которые группируются по 10≈30 (стопками) в граны (до 150 в Х.), соединяющиеся между собой крупными тилакоидами. При таком строении значительно увеличивается фотоактивная поверхность Х. и обеспечивается максимальное использование световой энергии. В мембране тилакоидов, состоящей из двух слоев белка, разделённых слоем липидов, осуществляется первичная световая стадия фотосинтеза, ведущая к образованию двух необходимых для ассимиляции CO2 соединений ≈ восстановленного никотинамид-адениндинуклеотидфосфата (НАДФ×Н) и богатого энергией соединения аденозинтрифосфата (АТФ). Источником энергии для образования молекул АТФ является разность потенциалов, которая образуется на мембране в результате векторного (направленного) переноса заряда. Разделение заряда по обеим сторонам мембраны обеспечивается особым расположением компонентов электронно-транспортной цепи в мембране, перешнуровывающих её толщу. Благодаря мембранам, играющим роль «перегородок», осуществляется пространственное разобщение продуктов фотосинтеза, например O2 и восстановителей, без которых эти продукты взаимодействовали бы друг с другом. Наружная поверхность тилакоида покрыта частицами диаметром 14≈15 нм, которые представляют собой «факторы сопряжения», участвуют в синтезе АТФ. В строме же сосредоточены ферменты фиксации CO2; (темновая стадия фотосинтеза).
У растений, способных к «кооперативному» фотосинтезу, существует 2 типа Х., различающихся по строению и функциям. Одни из них, находящиеся в клетках мезофилла, мелкие с гранами, другие, более крупные, содержатся в клетках обкладки проводящих сосудистых пучков, граны в них лишь зачаточные или совсем отсутствуют. В Х. второго типа функционирует фотосистема 1, которая образует АТФ в ходе циклического фосфорилирования, а НАДФ×Н ≈ за счёт реакции декарбоксилирования яблочной кислоты. Х. клеток обкладки фиксируют CO2 на рибулозодифосфате, т. е. с помощью цикла Калвина, а Х. клеток мезофилла ≈ на фосфоенолпирувате (путь Хетча ≈ Слэка); т. о. взаимодействие Х. обоих типов обеспечивает высокую эффективность фотосинтеза у растений. В строму Х., наряду с ферментами фиксации CO2, включены нити ДНК, рибосомы, крахмальные зёрна, осмиофильные гранулы.
Наличие в Х. собственного генетического аппарата и специфической белоксинтезирующей системы обусловливает определённую, хотя и относительную, автономию Х. в клетке. При развитии и размножении растения в новых генерациях клеток Х. возникают только путём деления. Происхождение Х. связывают с симбиогенезом , полагая, что современные Х. ≈ потомки сине-зелёных водорослей, вступившие в симбиоз с древними ядерными гетеротрофными клетками бесцветных водорослей или простейших.
Х. занимают 20≈30% объёма растительной клетки. У водорослей, например хламидомонады, имеется один Х., в клетке высших растений содержится от 10 до 70 Х. Развиваются Х. из т. н. инициальных частиц, или пропластид, ≈ небольших пузырьков, отделяющихся от ядра. В конце вегетации растения Х. в результате разрушения хлорофилла утрачивают зелёную окраску и превращаются в хромопласты . См. также Фотосинтез .
Лит.: Хлоропласты и митохондрии. Вопросы мембранной биологии, Сб., М., 1969; Лёви А., Сикевиц Ф., Структура и функция клетки, пер. с англ., М., 1971; Хит О., Фотосинтез, пер. с англ., М., 1972; Баславская С. С., Фотосинтез, М., 1974; Насыров Ю. С., Фотосинтез и генетика хлоропластов, М., 1975; Structure and function orchloroplasts, ed. М. Gibbs, B., 1971.
Р. М. Бекина.
Источник: xn--b1algemdcsb.xn--p1ai
Федеральное Агентство науки и образования.
Сибирский Федеральный Университет.
Институт Фундаментальной Биологии и Биотехнологии.
Кафедра биотехнологии.
РЕФЕРАТ
На тему: Строение и функции хлоропластов.
Выполнила: студентка
31гр.Шестопалова Н.С.
Проверила:
доцент кафедры
биотехнологии
д.б.н. Голованова Т.И.
Красноярск
2008г.
Содержание:
1. Введение………………………………………………………………………3
2. Обзор литературы……………………………………………………….4
2.1 Происхождение хлоропласта…………………………………………4
2.2 Развитие хлоропласта из пропластиды…………………………….5
2.3 Строение хлоропластов……………………………………………..7
2.4 Генетический аппарат хлоропластов……………………………….9
3. Функции хлоропластов………………………………………………11
4. Вывод…………………………………………………………………16
5. Список используемой литературы………………………………….17
Введение:
Пластиды –это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов(высшие растения, низшие водоросли, некоторые одноклеточные организмы). У высших растений найден целый набор различных пластид( хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой которая осуществляет фотосинтетические процессы, является хлоропласт.
2.Обзор литературы:
2.1Происхождение хлоропласта.
Общепринятым в настоящее время является представление об эндосимбиотическом происхождении хлоропластов в клетках растений. Хорошо известно, что лишайники представляют собой форму сожительства (симбиоза) гриба и водоросли, при котором зеленые одноклеточные водоросли живут внутри клеток гриба. Предполагают, что таким же путем несколько миллиардов лет назад фотосинтезирующие цианобактерии (синезеленые водоросли) проникли в эукариотические клетки и затем в ходе эволюции потеряли свою автономность, передав большое число важнейших генов в ядерный геном. В результате независимая бактериальная клетка превратилась в полуавтономную органеллу, сохранившую главную исходную функцию — способность к фотосинтезу, однако формирование фотосинтетического аппарата оказалось под двойным ядерно-хлоропластным контролем. Под ядерный контроль перешли деление хлоропластов и сам процесс реализации его генетической информации, которая осуществляется в цепи событий ДНК РНК белок.
Неоспоримые доказательства прокариотического происхождения хлоропластов получены при анализе нуклеотидных последовательностей их ДНК. ДНК рибосомальных генов имеет высокую степень сродства (гомологию) у хлоропластов и бактерий. Сходная нуклеотидная последовательность обнаружена для цианобактерий и хлоропластов в генах АТФсинтазного комплекса, а также в генах аппарата транскрипции (гены субъединиц РНК-полимеразы) и трансляции. Регуляторные элементы хлоропластных генов — промоторы, локализованные в области 35-10 пар нуклеотидов до начала транскрипции, определяющие считку генетической информации, и терминальные нуклеотидные последовательности, определяющие ее прекращение, организованы в хлоропласте, как упоминалось выше, по бактериальному типу. И хотя миллиарды лет эволюции внесли массу изменений в хлоропласт, они не изменили нуклеотидную последовательность хлоропластных генов, и это является неоспоримым доказательством происхождения хлоропласта в зеленом растении от прокариотического предка, древнего предшественника современных цианобактерий.
2.2Развитие хлоропласта из пропластиды.
Хлоропласт развивается из пропластиды — маленькой бесцветной органеллы (несколько микрон в поперечнике), окруженной двойной мембраной и содержащей характерную для хлоропласта кольцевую молекулу ДНК. Пропластиды не имеют внутренней мембранной системы. Они плохо изучены ввиду их крайне малых размеров. Несколько пропластид содержится в цитоплазме яйцеклетки. Они делятся и передаются от клетки к клетке в ходе развития зародыша. Этим объясняется то обстоятельство, что генетические признаки, связанные с ДНК пластид, передаются только по материнской линии (так называемая цитоплазматическая наследственность).
В ходе развития хлоропласта из пропластиды внутренняя мембрана ее оболочки образует "впячивания" внутрь пластиды. Из них развиваются мембраны тилакоидов, которые создают стопки — граны и ламеллы стромы. В темноте пропластиды дают начало формированию предшественника хлоропласта (этиопласта), который содержит структуру, напоминающую кристаллическую решетку. При освещении эта структура разрушается и происходит формирование характерной для хлоропласта внутренней структуры, состоящей из тилакоидов гран и ламелл стромы.
В клетках меристемы содержится несколько пропластид. При формировании зеленого листа они делятся и превращаются в хлоропласты. Например, в клетке закончившего рост листа пшеницы содержится около 150 хлоропластов. В органах растений, запасающих крахмал, например в клубнях картофеля, крахмальные зерна формируются и накапливаются в пластидах, называемых амилопластами. Как выяснилось, амилопласты, как и хлоропласты, образуются из тех же пропластид и содержат такую же ДНК, как хлоропласты. Они формируются в результате дифференцировки пропластид по другому пути, чем у хлоропластов. Известны случаи превращения хлоропластов в амилопласты и наоборот. Например, часть амилопластов превращается в хлоропласты при позеленении клубней картофеля на свету.В ходе созревания плодов томатов и некоторых других растений, а также в лепестках цветков и осенних красных листьях хлоропласты превращаются в хромопласты — органеллы, содержащие оранжевые пигменты каротиноиды. Такое превращение связано с разрушением структуры тилакоидов гран и приобретением органеллой совершенно иной внутренней организации. Эту перестройку пластиде диктует ядро, и она осуществляется с помощью особых белков, кодируемых в ядре и синтезируемых в цитоплазме. Например, кодируемый в ядре 58 кДа полипептид, образующий комплекс с каротиноидами, составляет половину всего белка мембранных структур хромопласта. Так, на основе одной и той же собственной ДНК в результате ядерно-цитоплазматического влияния пропластида может развиваться в зеленый фотосинтезирующий хлоропласт, белый, содержащий крахмал амилопласт или оранжевый, заполненный каротиноидами хромопласт. Между ними возможны превращения. Это интересный пример различных путей дифференцировки органелл на основе одной и той же собственной ДНК, но под влиянием ядерно-цитоплазматического "диктата".
2.3Строение хлоропласта.
Хлоропласты — пластиды высших растений, в которых идет процесс фотосинтеза, т. е. использование энергии световых лучей для образования из неорганических веществ (углекислого газа и воды) органических веществ с одновременным выделением в атмосферу кислорода. Хлоропласты имеют форму двояковыпуклой линзы, размер их около 4-6 мкм. Находятся они в паренхимных клетках листьев и других зеленых частей высших растений. Число их в клетке варьирует в пределах 25-50.
Снаружи хлоропласт покрыт оболочкой, состоящей из двух липопротеиновых мембран, внешней и внутренней. Обе мембраны имеют толщину около 7нм, они отделены друг от друга межмембранным пространством около 20-30нм. Внутренняя мембрана хлоропластов, как и других пластид образует складчатые впячивания внутрь матрикса или стромы. В зрелом хлоропласте высших растений видны два типа внутренних мембран. Это- мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Связь внутренней мембраны хлоропласта с мембранными структурами внутри него хорошо прослеживается на примере мембран ламелл стромы. В этом случае внутренняя мембрана хлоропласта образует узкую (шириной около 20нм.) складку, которая может простираться почти через всю пластиду. Таким образом, ламелла стромы может представлять собой плоский полый мешок или же иметь вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно и не образуют связей между собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами. Число тилакоидов на одну грану варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом как бы связывают между собой отдельные граны хлоропластов. Однако полости камер тилакоидов всегда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы.
В матриксе ( строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
В хлоропластах содержатся различные пигменты. В зависимости от вида растений это:
хлорофилл:
— хлорофилл А (сине-зеленый) — 70 % (у высших растений и зеленых водорослей);
— хлорофилл В (желто-зеленый) — 30 % (там же);
— хлорофилл С, D и E встречается реже — у других групп водорослей;
Источник: MirZnanii.com