Женский пол у человека является
Пол возникает сначала как чисто репродуктивное (рекомбинационное) явление. В процессе эволюции он постепенно приобретает также и эволюционные функции. Одновременно и определение пола закономерно переходит от генного (у гермафродитов) к хромосомному (у раздельнополых форм начиная, видимо, с рыб) и геномному (у пчел). Параллельно повышается уровень дифференциации полов и происходит увеличение проявления полового диморфизма: у бесполых форм и у гермафродитов он отсутствует, у раздельнополых моногамов появляется организменный половой диморфизм (вторичных половых признаков), у раздельнополых полигамов — популяционный, включающий половой диморфизм по численности и дисперсии полов.
В ходе онтогенеза определение пола может происходить в момент оплодотворения (хромосомные механизмы), а также контролироваться внутренними (гормоны) и/или внешними факторами. У человека и высших животных большую роль в развитии полового поведения играет также воспитание и обучение.
В организме женщины каждая соматическая клетка содержит 23 пары хромосом, причем хромосомы каждой пары сходны между собой. У мужчин имеется лишь пары, состоящие из сходных хромосом, а 23-я пара образована длинной Х-хромосомой и очень короткой У-хромосомой. Одна из пар хромосом женщины состоит из двух Х-хромоеом. В этих-то различиях и лежит секрет определения пола. Все яйцеклетки содержат Х-хромосому, тогда как среди сперматозоидов половина также содержит Х-хромосому, а другая половина — У-хромосому.
Пол будущего ребенка зависит от того, какой сперматозоид оплодотворил яйцеклетку; если это был сперматозоид с Х-хромосомой, то родится девочка, если же с У-хромосомой — то мальчик. Гормоны, конечно, играют важную роль в определении многих различий между особями разного пола, однако именно хромосомы дают толчок всей цепи событий, которая приведет к подавлению признаков одного и к проявлению признаков другого пола.
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомной теории, пол организма определяется половыми хромосомами (гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
— самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
— самки XX самцы XY (Lygaeus-тип)
— самки XX самцы ХО (Protenor-тип)
— самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
— самки ZW самцы ZZ
— самки Z0 самцы ZZ
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как X и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторые рыбы, растения и др.
Существует два основных типа XY-детерминации пола:
— Как у человека: пол зависит от наличия Y-хромосомы (если она есть, проявляется фенотип самца, если нет — самки). Так, при мутациях, связанных с изменением числа половых хромосом, особи, имеющие в генотипе комбинации XY, XXY, XYY, XXYY и т. п. будут обладать мужским фенотипом различной степени выраженности, а имеющие комбинации XX, X, XXX, ХХХХ и т. п. — женским.
— Как у представителей рода мушек-дрозофил: пол определяется по соотношению числа X-хромосом и числа аутосом, наличие же Y-хромосомы никак не влияет на детерминацию пола, но самцы без неё стерильны, так как в ней находятся гены, ответственные за сперматогенез.
У многих других организмов (птицы, некоторые рептилии и рыбы, бабочки, ручейники, из растений — земляника) наблюдается обратная картина — гомогаметен мужской пол (имеет две гомологичные хромосомы Z), а гетерогаметен женский (имеет одну Z-хромосому и одну состоящую в основном из гетерохроматина и потому генетически инертную W-хромосому).
Вероятно, исходным для бабочек механизмом определения пола был механизм ZO самка/ZZ самец*Ч Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечающий за развитие женского пола ген Fern в W-хромосоме.
При этом механизме определения пола один из полов (гомогаметный) обладает двумя X-хромосомами, в то время, как второй (гетерогаметный) только одной. При этом пол определяется так же, как и у дрозофил: по соотношению числа Х-хромосом и аутосом. ЭЙгот механизм определения пола обнаружен у некоторых насекомых (клопов, бабочек и др.) и круглых червей. У нематоды Caenorhabditis elegans при наборе половых хромосом XX формируется гермафродит, а при наборе ХО — самец.
Источник: students-library.com
26. Генотип как целостная система. Взаимодействие генов, множественное действие генов.
Генотип как целостная система
Свойства генов. На основании знакомства с примерами наследования признаков при моно- и дигибридном скрещивании может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов, каждый из которых определяет развитие только своего признака или свойства. Такое представление о прямой и однозначной связи гена с признаком чаще всего не соответствует действительности. На самом деле существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
-
Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
-
Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
-
Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
-
Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
-
Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) — это множественные гены, или полигены.
-
Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
-
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование —тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы (рис. 3.5). Как видно из схемы, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).

Рис. 3.5. Наследование окраски цветков при неполном доминировании у ночной красавицы.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса (см. § 3.7).
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Эритроциты людей этой группы имеют два типа антигенов: антиген А (детерминируемый геном / имеющимся в одной из хромосом) и антиген В (детерминируемый геном /а, локализованным в другой гомологичной хромосоме). Только в этом случае проявляют свое действие оба аллеля — 1А (в гомозиготном состоянии контролирует II группу крови, группу А) и IB (в гомозиготном состоянии контролирует III группу крови, группу В). Аллели 1А и IB работают в гетерозиготе как бы независимо друг от друга.
Пример наследования групп крови иллюстрирует и прояв-ление множественного аллелизма: ген/может быть представлен тремя разными аллелями, а есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно-жественных аллелей, из которых каждый диплоидный организм может иметь два любых аллеля (и только). Между этими аллелями возможны все перечисленные варианты аллельных взаимодействий.
Явление множественного аллелизма распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при оплодотворении у грибов, опылении у семенных растений, детерминирующих окраску шерсти животных и т. д.
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
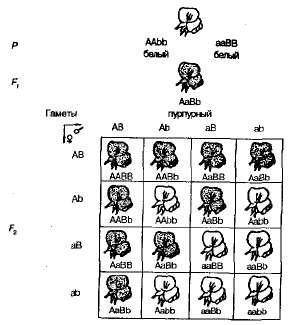
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ, то
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые (знак «—» в формуле генотипа обозначает, что это место может занять как доминантный, так и рецессивный аллель).
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16. Решетка Пеннета наглядно показывает причину этого явления (рис. 3.6).
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический.
По изменению числа и соотношения фенотип и чес ких классов при дигибридном расщеплении в F2 рассматривают несколько типов эпистатических взаимодействий: доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипичес-ким выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2 .
Рис. 3.6. Наследование окраски цветков у душистого горошка
Тогда все генотипы (А1 ,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растения а1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
По типу кумулятивной полимерии наследуются многие признаки: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Таким образом, многочисленные наблюдения свидетельствуют о том, что проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
Взаимодействие генов
Отношение между генами и признаками достаточно сложное. В организме не всегда один ген определяет только один признак и, наоборот, один признак определяется только одним геном. Чаще один ген может способствовать проявлению сразу нескольких признаков, и наоборот. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Множественное действие генов (плейотропия) — процессы влияния одного гена на формирование нескольких признаков.
Например, у человека ген, определяющий рыжую окраску волос, обусловливает более светлую кожу и появление веснушек.
Иногда гены, определяющие морфологические признаки, влияют на физиологические функции, снижая жизнестойкость и плодовитость, или оказываются летальными. Так, ген, вызывающий голубую окраску у норки, снижает ее плодовитость. Доминантный ген серой окраски у каракулевых овец в гомозиготном состоянии детален, поскольку у таких ягнят недоразвит желудок и они погибают при переходе на питание травой.
Комплементарное взаимодействие генов. На развитие одного признака могут влиять несколько генов. Взаимодействие нескольких неаллельных генов, приводящее к развитию одного признака, называется комплементарным. Например, у кур имеются четыре формы гребня, проявление какой-либо из них связано со взаимодействием двух пар неаллельных генов. Розовидный гребень обусловлен действием доминантного гена одной аллели, гороховидный — доминантного гена другой аллели. У гибридов при наличии двух доминантных неаллельных генов образуется ореховидный гребень, а при отсутствии всех доминантных генов, т.е. у рецессивной гомозиготы по двум неаллельным генам, образуется простой гребень.
Результатом взаимодействия генов является окраска шерсти у собак, мышей, лошадей, форма тыквы, окраска цветков душистого горошка.
Полимерия — такое взаимодействие неаллельных генов, когда степень развития признака зависит от общего количества доминантных генов. По этому принципу наследуется окраска зерен овса, пшеницы, цвет кожи у человека. Например, у негров в двух парах неаллельных генов 4 доминантных, а у людей с белой кожей — ни одного, все гены рецессивные. Сочетания разного количества доминантных и рецессивных генов приводят к образованию мулатов с разной интенсивностью окраски кожи: от темной до светлой.
Различают две основных группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами. Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме происходит взаимодействие между белками — ферментами, синтез которых опрелятся генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1) для образования определенного признака необходимо взаимодействие двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2) фермент, что был синтезирован с участием одного гена, полностью подавляет или инактивирует действие фермента, что был образован другим неаллельным геном;
3) два ферменты, образование которых контролируется двумя неаллельми генами, влияющими на один признак или на один процесс так, что их совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов
Гены, которые занимают идентичные (гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия — полное доминирование, которое впервые описано Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление одной из аллелей доминирует над проявлением другой. При полном доминировании расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу — 3:1. В медицинской практике с двух тысяч моногенных наследственных болезней почти в половины имеет место доминированое проявления патологических генов над нормальными. В гетерозигот патологический аллель проявляется в большинстве случаев признаками заболевания (доминантный фенотип).
Неполное доминирование — форма взаимодействия, при которой у гетерозиготного организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а), вследствие чего проявляется промежуточный между родительскими признак. Здесь расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов вызывает формирование зависимого от него продукта, то есть оказываются продукты обеих аллелей. Классическим примером такого проявления является система групп крови, в частности система АBО, когда эритроциты человека несут на поверхности антигены, контролируемые обеими аллелями. Такая форма проявления носит название кодоминированием.
Сверхдоминирование — когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе АА-нормальная продолжительность жизни; Аа — удлиненная триватисть жизни; аа — летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в природе количество аллелей может быть более двух, если какой то локус может находится в разных состояниях. В таких случаях говорят о множественные аллели или множественный аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами, например: А, А1, А3 … Аллельные гена локализуются в одинаковых участках гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с различием гомологичних хромосом) попадает только по одному из них. Для множественных аллелей характерное влияние всех аллелей на один и тот же признак. Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивный относительно первого, однако доминирует над следующими и т.д. Одним из примеров проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение, поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделивских закономерностей расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность, эпистаз, полимерию и модифицирующее действие (плейотропия).
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) — альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка — интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
Источник: StudFiles.net
26. Генотип как целостная система. Взаимодействие генов, множественное действие генов.
Генотип как целостная система
Свойства генов. На основании знакомства с примерами наследования признаков при моно- и дигибридном скрещивании может сложиться впечатление, что генотип организма слагается из суммы отдельных, независимо действующих генов, каждый из которых определяет развитие только своего признака или свойства. Такое представление о прямой и однозначной связи гена с признаком чаще всего не соответствует действительности. На самом деле существует огромное количество признаков и свойств живых организмов, которые определяются двумя и более парами генов, и наоборот, один ген часто контролирует многие признаки. Кроме того, действие гена может быть изменено соседством других генов и условиями внешней среды. Таким образом, в онтогенезе действуют не отдельные гены, а весь генотип как целостная система со сложными связями и взаимодействиями между ее компонентами. Эта система динамична: появление в результате мутаций новых аллелей или генов, формирование новых хромосом и даже новых геномов приводит к заметному изменению генотипа во времени.
Характер проявления действия гена в составе генотипа как системы может изменяться в различных ситуациях и под влиянием различных факторов. В этом можно легко убедится, если рассмотреть свойства генов и особенности их проявления в признаках:
-
Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов.
-
Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма.
-
Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена).
-
Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
-
Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков) — это множественные гены, или полигены.
-
Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков. Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
-
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды.
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов —неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование.
Доминирование —тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы (рис. 3.5). Как видно из схемы, гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. При скрещивании растения с красными цветками и растения с белыми цветками в F1, у всех растений цветки розовые, т. е. наблюдается промежуточный характер наследования. При скрещивании гибридов с розовой окраской цветков в F2 имеет место совпадение расщепления по фенотипу и генотипу, так как доминантная гомозигота (АА) отличается от гетерозиготы (Аа). Так, в рассматриваемом примере с растениями ночной красавицы расщепление в F2 по окраске цветков обычно следующее — 1 красная (АА): 2 розовые (Аа): 1 белая (аа).
Рис. 3.5. Наследование окраски цветков при неполном доминировании у ночной красавицы.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса (см. § 3.7).
Кодаминирвание— участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Эритроциты людей этой группы имеют два типа антигенов: антиген А (детерминируемый геном / имеющимся в одной из хромосом) и антиген В (детерминируемый геном /а, локализованным в другой гомологичной хромосоме). Только в этом случае проявляют свое действие оба аллеля — 1А (в гомозиготном состоянии контролирует II группу крови, группу А) и IB (в гомозиготном состоянии контролирует III группу крови, группу В). Аллели 1А и IB работают в гетерозиготе как бы независимо друг от друга.
Пример наследования групп крови иллюстрирует и прояв-ление множественного аллелизма: ген/может быть представлен тремя разными аллелями, а есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно-жественных аллелей, из которых каждый диплоидный организм может иметь два любых аллеля (и только). Между этими аллелями возможны все перечисленные варианты аллельных взаимодействий.
Явление множественного аллелизма распространено в природе. Известны обширные серии множественных аллелей, определяющих тип совместимости при оплодотворении у грибов, опылении у семенных растений, детерминирующих окраску шерсти животных и т. д.
Взаимодействия неаллельных генов.Неаллельные взаимодействия генов описаны у многих растений и животных. Они приводят к появлению в потомстве дигетерозиготы необычного расщепления по фенотипу: 9:3:4; 9:6:1; 13:3; 12:3:1; 15:1, т.е. модификации общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия двух, трех и большего числа неаллельных генов. Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарным, или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате которого появляется признак, отсутствующий у обоих родителей. Например, при скрещивании двух сортов душистого горошка с белыми цветками появляется потомство с пурпурными цветками. Если обозначить генотип одного сорта ААbb, а другого — ааВВ, то
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, вто время как поодиночке ни ген А, ни ген B не обеспечивали синтез этого пигмента. Синтез антоциана представляет собой сложную цепь последовательных биохимических реакций, контролируемых несколькими неаллельными генами, и только при наличии как минимум двух доминантных генов (А-В-) развивается пурпурная окраска. В остальных случаях {ааВ- и A-bb) цветки у растения белые (знак «—» в формуле генотипа обозначает, что это место может занять как доминантный, так и рецессивный аллель).
При самоопылении растений душистого горошка из F1 в F2 наблюдалось расщепление на пурпурно- и белоцветковые формы в соотношении, близком к 9:7. Пурпурные цветки были обнаружены у 9/16 растений, белые — у 7/16. Решетка Пеннета наглядно показывает причину этого явления (рис. 3.6).
Эпистаз — это такой тип взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена. Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами. Подавляемый ген носит название гипостатический.
По изменению числа и соотношения фенотип и чес ких классов при дигибридном расщеплении в F2 рассматривают несколько типов эпистатических взаимодействий: доминантный эпистаз (А>В или В>А) с расщеплением 12:3:1; рецессивный эпистаз (а>В или b>А), который выражается в расщеплении 9:3:4, и т. д.
Полимерия проявляется в том, что один признак формируется под влиянием нескольких генов с одинаковым фенотипичес-ким выражением. Такие гены называются полимерными. В этом случае принят принцип однозначного действия генов на развитие признака. Например, при скрещивании растений пастушьей сумки с треугольными и овальными плодами (стручочками) в F1 образуются растения с плодами треугольной формы. При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными стручочками в соотношении 15:1. Это объясняется тем, что существуют два гена, действующих однозначно. В этих случаях их обозначают одинаково— А1и A2 .
Рис. 3.6. Наследование окраски цветков у душистого горошка
Тогда все генотипы (А1 ,-А2,-, А1-а2а2, a1a1A2-) будут иметь одинаковый фенотип — треугольные стручочки, и только растения а1а1а2a2 будут отличаться —- образовывать овальные стручочки. Это случай некумулятивной полимерии.
Полимерные гены могут действовать и по типу кумулятивной полимерии. Чем больше подобных генов в генотипе организма, тем сильнее проявление данного признака, т. е. с увеличением дозы гена (А1 А2 А3 и т. д.) его действие суммируется, или кумулируется. Например, интенсивность окраски эндосперма зерен пшеницы пропорциональна числу доминантных аллелей разных генов в тригибридном скрещивании. Наиболее окрашенными были зерна А1А1А2А2А3,А 3 а зерна а1а1а2a2а3а 3 не имели пигмента.
По типу кумулятивной полимерии наследуются многие признаки: молочность, яйценоскость, масса и другие признаки сельскохозяйственных животных; многие важные параметры физической силы, здоровья и умственных способностей человека; длина колоса у злаков; содержание сахара в корнеплодах сахарной свеклы или липидов в семенах подсолнечника и т. д.
Таким образом, многочисленные наблюдения свидетельствуют о том, что проявление большей части признаков представляет собой результат влияния комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
Взаимодействие генов
Отношение между генами и признаками достаточно сложное. В организме не всегда один ген определяет только один признак и, наоборот, один признак определяется только одним геном. Чаще один ген может способствовать проявлению сразу нескольких признаков, и наоборот. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Множественное действие генов (плейотропия) — процессы влияния одного гена на формирование нескольких признаков.
Например, у человека ген, определяющий рыжую окраску волос, обусловливает более светлую кожу и появление веснушек.
Иногда гены, определяющие морфологические признаки, влияют на физиологические функции, снижая жизнестойкость и плодовитость, или оказываются летальными. Так, ген, вызывающий голубую окраску у норки, снижает ее плодовитость. Доминантный ген серой окраски у каракулевых овец в гомозиготном состоянии детален, поскольку у таких ягнят недоразвит желудок и они погибают при переходе на питание травой.
Комплементарное взаимодействие генов. На развитие одного признака могут влиять несколько генов. Взаимодействие нескольких неаллельных генов, приводящее к развитию одного признака, называется комплементарным. Например, у кур имеются четыре формы гребня, проявление какой-либо из них связано со взаимодействием двух пар неаллельных генов. Розовидный гребень обусловлен действием доминантного гена одной аллели, гороховидный — доминантного гена другой аллели. У гибридов при наличии двух доминантных неаллельных генов образуется ореховидный гребень, а при отсутствии всех доминантных генов, т.е. у рецессивной гомозиготы по двум неаллельным генам, образуется простой гребень.
Результатом взаимодействия генов является окраска шерсти у собак, мышей, лошадей, форма тыквы, окраска цветков душистого горошка.
Полимерия — такое взаимодействие неаллельных генов, когда степень развития признака зависит от общего количества доминантных генов. По этому принципу наследуется окраска зерен овса, пшеницы, цвет кожи у человека. Например, у негров в двух парах неаллельных генов 4 доминантных, а у людей с белой кожей — ни одного, все гены рецессивные. Сочетания разного количества доминантных и рецессивных генов приводят к образованию мулатов с разной интенсивностью окраски кожи: от темной до светлой.
Различают две основных группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами. Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме происходит взаимодействие между белками — ферментами, синтез которых опрелятся генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1) для образования определенного признака необходимо взаимодействие двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2) фермент, что был синтезирован с участием одного гена, полностью подавляет или инактивирует действие фермента, что был образован другим неаллельным геном;
3) два ферменты, образование которых контролируется двумя неаллельми генами, влияющими на один признак или на один процесс так, что их совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов
Гены, которые занимают идентичные (гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия — полное доминирование, которое впервые описано Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление одной из аллелей доминирует над проявлением другой. При полном доминировании расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу — 3:1. В медицинской практике с двух тысяч моногенных наследственных болезней почти в половины имеет место доминированое проявления патологических генов над нормальными. В гетерозигот патологический аллель проявляется в большинстве случаев признаками заболевания (доминантный фенотип).
Неполное доминирование — форма взаимодействия, при которой у гетерозиготного организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а), вследствие чего проявляется промежуточный между родительскими признак. Здесь расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов вызывает формирование зависимого от него продукта, то есть оказываются продукты обеих аллелей. Классическим примером такого проявления является система групп крови, в частности система АBО, когда эритроциты человека несут на поверхности антигены, контролируемые обеими аллелями. Такая форма проявления носит название кодоминированием.
Сверхдоминирование — когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе АА-нормальная продолжительность жизни; Аа — удлиненная триватисть жизни; аа — летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в природе количество аллелей может быть более двух, если какой то локус может находится в разных состояниях. В таких случаях говорят о множественные аллели или множественный аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами, например: А, А1, А3 … Аллельные гена локализуются в одинаковых участках гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с различием гомологичних хромосом) попадает только по одному из них. Для множественных аллелей характерное влияние всех аллелей на один и тот же признак. Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивный относительно первого, однако доминирует над следующими и т.д. Одним из примеров проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение, поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделивских закономерностей расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность, эпистаз, полимерию и модифицирующее действие (плейотропия).
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) — альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка — интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
Источник: StudFiles.net