Фотосинтез происходит в хлоропластах
Фотосинтез происходит в эукариотических клеточных структурах, называемых хлоропластами. Хлоропласт — это тип органеллы растительных клеток, известный как зеленые пластиды. Пластиды помогают хранить и собирать необходимые вещества для производства энергии. Хлоропласт содержит зеленый пигмент, называемый хлорофиллом, который поглощает световую энергию для процесса фотосинтеза. Следовательно, название хлоропласт указывает на то, что эти органеллы представляют собой хлорофиллсодержащие пластиды.
Подобно митохондриям, хлоропласты имеют свою собственную ДНК, ответственны за производство энергии и воспроизводятся независимо от остальной части клетки посредством процесса деления, подобного бактериальному бинарному делению. Они также ответственны за производство аминокислот и липидных компонентов, необходимых для производства хлоропластов. Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласт: структура
Хлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений.
ранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
- Мембрана — содержит внутренние и внешние липидные двухслойные оболочки, которые выступают в качестве защитных покрытий и сохраняют замкнутые структуры хлоропластов. Внутренняя мембрана отделяет строму от межмембранного пространства и регулирует прохождение молекул в/из хлоропласта.
- Межмембранное пространство — пространство между внешней и внутренней мембранами.
- Тилакоидная система — внутренняя система мембран, состоящая из сплющенных мешкообразных мембранных структур, называемых тилакоидами, которые служат местами преобразования энергии света в химическую энергию.
- Тилакоид с просветом (люменом) — отсек в каждом тилакоиде.
- Грана — плотные слоистые стопки тилакоидных мешков (10-20), которые служат местами преобразования энергии света в химическую энергию.
- Строма — плотная жидкость внутри хлоропласта, содержащая внутри оболочки, но вне тилакоидной мембраны. Здесь происходит конверсия углекислого газа в углеводы (сахара).
- Хлорофилл — зеленый фотосинтетический пигмент в хлоропласт-гране, поглощающий световую энергию.
Хлоропласт: фотосинтез
 » data-layzr=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg» alt=»» width=»500″ height=»365″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg 300w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-500×365.jpg 500w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет.jpg 650w» sizes=»(max-width: 500px) 100vw, 500px» />
» data-layzr=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg» alt=»» width=»500″ height=»365″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-300×219.jpg 300w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет-500×365.jpg 500w, https://natworld.info/wp-content/uploads/2017/05/фотосинтез-лист-свет.jpg 650w» sizes=»(max-width: 500px) 100vw, 500px» />
При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Источник: natworld.info
Фотосинтез
Фотосинтез — синтез органических веществ из углекислого газа и воды с обязательным использованием энергии света:
6СО2 + 6Н2О + Qсвета → С6Н12О6 + 6О2.
У высших растений органом фотосинтеза является лист, органоидами фотосинтеза — хлоропласты (строение хлоропластов — лекция №7). В мембраны тилакоидов хлоропластов встроены фотосинтетические пигменты: хлорофиллы и каротиноиды. Существует несколько разных типов хлорофилла (a, b, c, d), главным является хлорофилл a. В молекуле хлорофилла можно выделить порфириновую «головку» с атомом магния в центре и фитольный «хвост». Порфириновая «головка» представляет собой плоскую структуру, является гидрофильной и поэтому лежит на той поверхности мембраны, которая обращена к водной среде стромы. Фитольный «хвост» — гидрофобный и за счет этого удерживает молекулу хлорофилла в мембране.
Хлорофиллы поглощают красный и сине-фиолетовый свет, отражают зеленый и поэтому придают растениям характерную зеленую окраску. Молекулы хлорофилла в мембранах тилакоидов организованы в фотосистемы. У растений и синезеленых водорослей имеются фотосистема-1 и фотосистема-2, у фотосинтезирующих бактерий — фотосистема-1. Только фотосистема-2 может разлагать воду с выделением кислорода и отбирать электроны у водорода воды.
Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы.
Световая фаза
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды:
Н2О + Qсвета → Н+ + ОН—.
Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН:
ОН— → •ОН + е—.
Радикалы •ОН объединяются, образуя воду и свободный кислород:
4НО• → 2Н2О + О2.
Кислород при этом удаляется во внешнюю среду, а протоны накапливаются внутри тилакоида в «протонном резервуаре». В результате мембрана тилакоида с одной стороны за счет Н+ заряжается положительно, с другой за счет электронов — отрицательно. Когда разность потенциалов между наружной и внутренней сторонами мембраны тилакоида достигает 200 мВ, протоны проталкиваются через каналы АТФ-синтетазы и происходит фосфорилирование АДФ до АТФ; атомарный водород идет на восстановление специфического переносчика НАДФ+ (никотинамидадениндинуклеотидфосфат) до НАДФ·Н2:
2Н+ + 2е— + НАДФ → НАДФ·Н2.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
1 — строма хлоропласта; 2 — тилакоид граны.
Темновая фаза
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является пятиуглеродный сахар рибулозобифосфат (РиБФ); катализирует реакцию фермент рибулозобифосфат-карбоксилаза (РиБФ-карбоксилаза). В результате карбоксилирования рибулозобисфосфата образуется неустойчивое шестиуглеродное соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК). Затем происходит цикл реакций, в которых через ряд промежуточных продуктов фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина»:
6СО2 + 24Н+ + АТФ → С6Н12О6 + 6Н2О.
Кроме глюкозы, в процессе фотосинтеза образуются другие мономеры сложных органических соединений — аминокислоты, глицерин и жирные кислоты, нуклеотиды. В настоящее время различают два типа фотосинтеза: С3— и С4-фотосинтез.
С3-фотосинтез
Это тип фотосинтеза, при котором первым продуктом являются трехуглеродные (С3) соединения. С3-фотосинтез был открыт раньше С4-фотосинтеза (М. Кальвин). Именно С3-фотосинтез описан выше, в рубрике «Темновая фаза». Характерные особенности С3-фотосинтеза: 1) акцептором углекислого газа является РиБФ, 2) реакцию карбоксилирования РиБФ катализирует РиБФ-карбоксилаза, 3) в результате карбоксилирования РиБФ образуется шестиуглеродное соединение, которое распадается на две ФГК. ФГК восстанавливается до триозофосфатов (ТФ). Часть ТФ идет на регенерацию РиБФ, часть превращается в глюкозу.
Фотодыхание
Это светозависимое поглощение кислорода и выделение углекислого газа. Еще в начале прошлого века было установлено, что кислород подавляет фотосинтез. Как оказалось, для РиБФ-карбоксилазы субстратом может быть не только углекислый газ, но и кислород:
О2 + РиБФ → фосфогликолат (2С) + ФГК (3С).
Фермент при этом называется РиБФ-оксигеназой. Кислород является конкурентным ингибитором фиксации углекислого газа. Фосфатная группа отщепляется, и фосфогликолат становится гликолатом, который растение должно утилизировать. Он поступает в пероксисомы, где окисляется до глицина. Глицин поступает в митохондрии, где окисляется до серина, при этом происходит потеря уже фиксированного углерода в виде СО2. В итоге две молекулы гликолата (2С + 2С) превращаются в одну ФГК (3С) и СО2. Фотодыхание приводит к понижению урожайности С3-растений на 30–40% (С3-растения — растения, для которых характерен С3-фотосинтез).
С4-фотосинтез
С4-фотосинтез — фотосинтез, при котором первым продуктом являются четырехуглеродные (С4) соединения. В 1965 году было установлено, что у некоторых растений (сахарный тростник, кукуруза, сорго, просо) первыми продуктами фотосинтеза являются четырехуглеродные кислоты. Такие растения назвали С4-растениями. В 1966 году австралийские ученые Хэтч и Слэк показали, что у С4-растений практически отсутствует фотодыхание и они гораздо эффективнее поглощают углекислый газ. Путь превращений углерода в С4-растениях стали называть путем Хэтча-Слэка.
Для С4-растений характерно особое анатомическое строение листа. Все проводящие пучки окружены двойным слоем клеток: наружный — клетки мезофилла, внутренний — клетки обкладки.
лекислый газ фиксируется в цитоплазме клеток мезофилла, акцептор — фосфоенолпируват (ФЕП, 3С), в результате карбоксилирования ФЕП образуется оксалоацетат (4С). Процесс катализируется ФЕП-карбоксилазой. В отличие от РиБФ-карбоксилазы ФЕП-карбоксилаза обладает большим сродством к СО2 и, самое главное, не взаимодействует с О2. В хлоропластах мезофилла много гран, где активно идут реакции световой фазы. В хлоропластах клеток обкладки идут реакции темновой фазы.
Оксалоацетат (4С) превращается в малат, который через плазмодесмы транспортируется в клетки обкладки. Здесь он декарбоксилируется и дегидрируется с образованием пирувата, СО2 и НАДФ·Н2.
Пируват возвращается в клетки мезофилла и регенерирует за счет энергии АТФ в ФЕП. СО2 вновь фиксируется РиБФ-карбоксилазой с образованием ФГК. Регенерация ФЕП требует энергии АТФ, поэтому нужно почти вдвое больше энергии, чем при С3-фотосинтезе.
|
Строение С4-растений: |
|
|
С4-фотосинтез: |
Значение фотосинтеза
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м2 поверхности в час.
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Источник: licey.net
1. Какие виды автотрофного питания вы знаете?
Ответ. Фототрофы — организмы, для которых источником энергии служит солнечный свет. Такой тип питания носит название фотосинтеза. К фотосинтезу способны зелёные растения и многоклеточные водоросли, а также цианобактерии и многие другие группы бактерий благодаря содержащемуся в их клетках пигменту — хлорофиллу. Археи из группы галобактерий способны к бесхлорофилльному фотосинтезу, при котором энергию света улавливает и преобразует белок бактериородопсин.
Хемотрофы — в качестве внешнего источника энергии используют энергию химических связей пищи или восстановленных неорганических соединений — таких, как сероводород, метан, сера, двухвалентное железо и др.
2. Как называются органоиды клетки, в которых происходит фотосинтез?
Ответ. Фотосинтез происходит в хлоропластах (у высших растений), в хромотофорах (у низших растений), в мезосомах у бактерий.
3. Что такое ароморфоз?
Ответ. Ароморфоз – прогрессивное эволюционное изменение строения, приводящее к общему повышению уровня организации организмов. Ароморфоз — это расширение жизненных условий, связанное с усложнением организации и повышением жизнедеятельности
Вопросы после § 24
1. Что представляла собой «великая кислородная революция»?
Ответ. Первичная атмосфера Земли в момент возникновения жизни состояла, по-видимому, из азота, аммиака, метана, водорода и паров воды, но почти не содержала кислорода. Когда в океане появились первые фотосинтезирующие прокариоты, а затем и эукариотические водоросли, атмосфера стала постепенно насыщаться кислородом. Когда содержание кислорода в атмосфере достигло 1 % от нынешнего (так называемая точка Пастера), у организмов, живших в то время, появилась возможность использовать его в процессах окисления органических соединений для получения энергии. Таким образом возникло клеточное дыхание, которое дало живым существам во много раз больше энергии, чем бескислородные процессы. Произошла так называемая «великая кислородная революция». Кислорода стало достаточно для того, чтобы мог возникнуть озоновый слой, защитивший от смертоносного действия ультрафиолета поверхности водоёмов и суши. Организмы, освоив новые, выгодные энергетические процессы, стали заселять поверхностные слои водоёмов, тогда как до этого им приходилось существовать на больших глубинах, чтобы не подвергаться воздействию ультрафиолетовых лучей. Теперь у фототрофов фотосинтез стал проходить более интенсивно, так как чем меньше слой воды, тем лучше он освещается солнцем. Виды живых существ, перешедшие к клеточному дыханию, резко усилили все процессы жизнедеятельности. Это, по всей видимости, способствовало ускорению прогрессивной эволюции. Многократно возросло количество видов, обитающих в воде. Через какое-то время первые живые существа выпели на сушу, надёжно защищённые от ультрафиолета озоновым слоем атмосферы
2. Какое соединение является источником углерода для сахаров, синтезированных в процессе фотосинтеза?
Ответ. Углекислый газ является источником углерода для сахаров, синтезированных в процессе фотосинтеза
3. Какие процессы происходят в световую фазу фотосинтеза? На каких структурах хлоропластов они протекают?
Ответ. Процесс световой фазы фотосинтеза растений включает в себя нециклическое фосфорилирование и фотолиз воды. Реакции происходят на мембранах хлоропластов.
Фотосистема I. Молекулы хлорофилла аI поглощают свет с длиной волны 700 нм. Электроны, получившие избыток энергии, участвуют в реакции диссоциации воды (Н2О = Н+ + ОН-). Электроны и ионы водорода реагируют с НАДФ+ (никотинамидадениндинуклеотидфосфата):
НАДФ+ + 2е + 2Н+ = НАДФ • Н + Н+.
Полученное в данной реакции вещество НАДФ • Н играет роль восстановителя в реакциях темновой фазы.
Процесс распада воды до Н+ и ОН-, протекающий при участии электронов, имеющих избыток энергии за счёт фотореакций, получил название фотолиза воды.
Фотосистема II. Молекулы хлорофилла аII поглощают свет с длиной волны 680 нм. Электроны с избыточной энергией по системе цитохромов переносятся на молекулы хлорофилла аI и занимают пустующие орбитали, которые раньше занимали электроны, связавшиеся с ионами водорода в ходе фотолиза воды. (При прохождении электронов по цепочке цитохромов часть их энергии используется для синтеза АТФ.) В результате возникает нехватка электронов в молекулах хлорофилла аII. Эта нехватка восполняется электронами гидроксид-анионов (ОН-), которые образовались в ходе того же фотолиза воды. Отдавая электроны молекулам хлорофилла аII, эти ионы превращаются в гидроксид-радикалы:
ОН- – e = ОН.
Гидроксид-радикал – это чрезвычайно неустойчивое химическое соединение, поэтому, только образовавшись, оно самопроизвольно превращается в воду и свободный кислород, выделяемый растением во внешнюю среду:
4OН = 2Н2O + O2
Таким образом, кислород, которым дышит подавляющее большинство живых организмов на Земле, представляет собой побочный продукт фотосинтеза, образующийся вследствие фотолиза воды
В реакциях световой фазы фотосинтеза накапливается энергия (НАДФ•Н и АТФ), которая тратится в процессах темновой фазы. Синтез АТФ из АДФ за счёт энергии света – очень эффективный процесс: за одно и то же время в хлоропластах образуется в 30 раз больше АТФ, чем в митохондриях
4. Какие процессы происходят в темновую фазу фотосинтеза? Где в хлоропластах они осуществляются?
Ответ.
Если световая фаза может протекать только при освещении растения, то реакции темновой фазы протекают независимо от света. Эти реакции осуществляются в строме хлоропластов, куда из тилакоидов поступают богатые энергией вещества: НАДФ•Н и АТФ. Источник углерода – СО2 – растение получает из воздуха через устьица. В реакциях темновой фазы СО2 восстанавливается до глюкозы, причём этот процесс протекает с затратами энергии, запасённой в молекулах АТФ и НАДФ•Н. Превращение углекислого газа в глюкозу в ходе темновой фазы фотосинтеза получило название цикла Кальвина, по имени его открывателя.
Общее уравнение фотосинтеза может быть представлено в виде:
12 Н2О → 12 [Н2] + 6 О2 (световая реакция)
6 СО2 + 12 [Н2] → С6Н12О6 + 6 Н2О (темновая реакция)
6 СО2 + 12 Н2О → С6Н12О6 + 6 Н2О + 6 О2 (суммарное уравнение фотосинтеза)
Источник: resheba.me
Строение листьев растений
Листья растений по анатомическому строению отличаются большим разнообразием, которое зависит и от вида растения, и от условий их роста. Лист сверху и снизу покрыт эпидермисом — покровной тканью с многочисленными отверстиями, называемыми устьицами. Под верхним эпидермисом расположена палисадная, или столбчатая паренхима, называемая ассимиляционной.
Под ней находится более рыхлая ткань — губчатая паренхима, за которой идет нижний эпидермис. Весь лист пронизан сетью жилок, состоящих из проводящих пучков, по которым проходят вода, минеральные и органические вещества.
В столбчатой и губчатой ткани листа расположены зеленые пластиды — хлоропласты, содержащие пигменты. Наличием хлоропластов и содержащихся в них зеленых пигментов (хлорофиллов) объясняется окраска растений.
Огромная листовая поверхность, достигающая 30 000 — 50 000 кв. м на 1 га у разных растений, хорошо приспособлена для успешного поглощения СО2 из воздуха в процессе фотосинтеза.
Углекислый газ проникает в лист растения через устьица, расположенные в эпидермисе, поступает в межклетники и, проникая через оболочку клеток, попадает в цитоплазму, а затем в хлоропласты, где и осуществляется процесс ассимиляции.
Образующийся в этом процессе кислород диффундирует с поверхности хлоропластов в свободном состоянии.
Таким образом, через устьица осуществляется газообмен листьев с внешней средой — поступление углекислого газа и выделение кислорода в процессе фотосинтеза, выделение углекислого газа и поглощение кислорода в процессе дыхания. Кроме того, устьица служат для выделения паров воды.
Несмотря на то, что общая площадь устьичных отверстий составляет лишь 1—2% всей листовой поверхности, тем не менее при открытых устьицах углекислый газ проникает в листья со скоростью, превышающей в 50 раз поглощение его щелочью. Количество устьиц очень велико — от нескольких десятков до 1500 на 1 кв. мм.
Хлоропласты
Хлоропласты — зеленые пластиды, в которых происходит процесс фотосинтеза. Они расположены в цитоплазме. У высших растений хлоропласты имеют дискообразную или линзовидную форму, у низших они более разнообразны.
Размер хлоропластов у высших растений довольно постоянен, составляя в среднем 1 —10 мк. Обычно в клетке содержится большое количество хлоропластов, в среднем 20—50, а иногда и больше. Расположены они главным образом в листьях, много их в незрелых плодах. В растении общее количество хлоропластов огромно; во взрослом дереве дуба, например, площадь их равняется 2 га.
Хлоропласт имеет мембранную структуру. От цитоплазмы он отделен двухмембранной оболочкой. В хлоропласте находятся ламеллы, белково-липоидные пластинки, собранные в пучки и называемые гранами.
Хлорофилл расположен в ламеллах в виде мономолекулярного слоя. Между ламеллами находится водянистая белковая жидкость — строма; в ней встречаются крахмальные зерна и капли масла.
Строение хлоропласта хорошо приспособлено к фотосинтезу, так как разделение хлорофиллоносного аппарата на мелкие пластинки значительно увеличивает активную поверхность хлоропласта, что облегчает доступ энергии и перенос ее к химическим системам, участвующим в фотосинтезе.
Данные А. А. Табенцкого показывают, что хлоропласты все время изменяются в онтогенезе растения. В молодых листьях наблюдается мелкогранулярная структура хлоропластов, в листьях, закончивших рост,— крупногранулярная.
В старых листьях уже наблюдается распад хлоропластов. В сухом веществе хлоропластов содержится 20—45% белков, 20—40% липоидов, 10—12% углеводов и других запасных веществ, 10% минеральных элементов, 5—10% зеленых пигментов (хлорофилл а и хлорофилл б), 1—2% каротиноидов, а также небольшое количество РНК и ДНК. Содержание воды достигает 75%.
В хлоропластах имеется большой набор гидролитических и окислительно-восстановительных ферментов. Исследованиями Н. М. Сисакяна показано, что в хлоропластах происходит и синтез многих ферментов. Благодаря этому они принимают участие во всем сложном комплексе процессов жизнедеятельности растения.
Пигменты, их свойства и условия образования
Пигменты можно извлечь из листьев растений спиртом или ацетоном. В вытяжке находятся следующие пигменты: зеленые — хлорофилл а и хлорофилл б; желтые — каротин и ксантофилл (каротиноиды).
Хлорофилл
Хлорофилл представляет собой
одно из интереснейших веществ на земной поверхности
(Ч. Дарвин),
так как благодаря ему возможен синтез органических веществ из неорганических СО2 и Н2О.
Хлорофилл не растворяется в воде, под влиянием солей, кислот и щелочей легко изменяется, поэтому было очень трудно установить его химический состав. Для извлечения хлорофилла обычно применяют этиловый спирт или ацетон.
Хлорофилл имеет следующие суммарные формулы: хлорофилл а — С55Н72О5N4Mg, хлорофилл б — С55Н70О6N4Mg.
У хлорофилла а больше на 2 атома водорода и меньше на 1 атом кислорода, чем у хлорофилла б. Формулы хлорофилла можно представить и так:
Центральное место в молекуле хлорофилла занимает Мg; его можно вытеснить, подействовав на спиртовую вытяжку хлорофилла соляной кислотой. Зеленый пигмент превращается в бурый, называемый феофитином, в котором Мg замещается двумя атомами Н из соляной кислоты.
Восстановить зеленый цвет вытяжки очень легко внесением в молекулу феофитина магния или другого металла. Следовательно, зеленый цвет хлорофилла связан с наличием в его составе металла.
При воздействии на спиртовую вытяжку хлорофилла щелочью происходит отщепление спиртовых групп (фитола и метилового спирта); в этом случае зеленая окраска хлорофилла сохраняется, что указывает на сохранение ядра молекулы хлорофилла при этой реакции.
Химический состав хлорофилла у всех растений одинаков. Содержание хлорофилла а всегда больше (примерно в 3 раза), чем хлорофилла б. Общее количество хлорофилла невелико и составляет около 1 % от сухого вещества листа.
По своей химической природе хлорофилл близок к красящему веществу крови — гемоглобину, центральное место в молекуле которого занимает не магний, а железо. В соответствии с этим различаются и их физиологические функции: хлорофилл принимает участие в важнейшем восстановительном процессе в растении — фотосинтезе, а гемоглобин — в процессе дыхания животных организмов, перенося кислород.
Оптические свойства пигментов
Хлорофилл поглощает солнечную энергию и направляет ее на химические реакции, которые не могут протекать без энергии, получаемой извне. Раствор хлорофилла в проходящем свете имеет зеленый цвет, но при увеличении толщины слоя или концентрации хлорофилла он приобретает красный цвет.
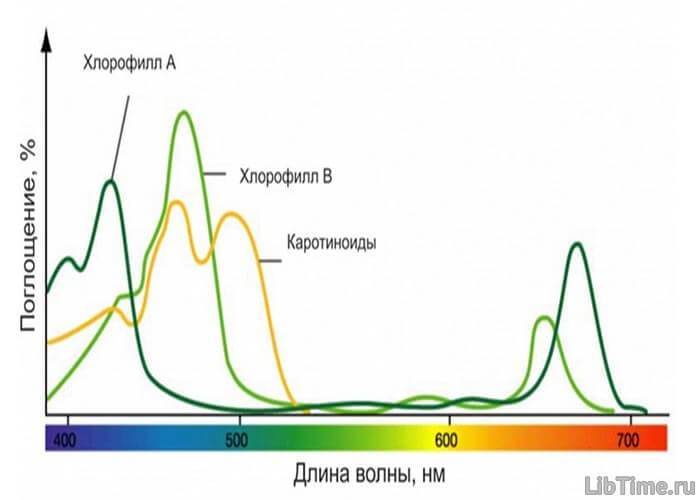
Хлорофилл поглощает свет не сплошь, а избирательно. При пропускании белого света через призму получается спектр, состоящий из семи видимых цветов, которые постепенно переходят друг в друга.
При пропускании белого света через призму и раствор хлорофилла на полученном спектре наиболее интенсивное поглощение будет в красных и сине-фиолетовых лучах. Зеленые лучи поглощаются мало, поэтому в тонком слое хлорофилл имеет в проходящем свете зеленый цвет.
Однако с увеличением концентрации хлорофилла полосы поглощения расширяются (значительная часть зеленых лучей также поглощается) и без поглощения проходит только часть крайних красных. Спектры поглощения хлорофилла а и б очень близки.
В отраженном свете хлорофилл кажется вишнево-красным, так как он излучает поглощенный свет с изменением длины его волны. Это свойство хлорофилла называется флюоресценцией.
Каротин и ксантофилл
Каротин и ксантофилл имеют полосы поглощения только в синих и фиолетовых лучах. Их спектры близки друг другу.
Поглощенная этими пигментами энергия передается хлорофиллу а, который является непосредственным участником фотосинтеза. Каротин считают провитамином А, так как при его расщеплении образуются 2 молекулы витамина А. Формула каротина — С40Н56, ксантофилла — С40Н54(ОН)2.
Условия образования хлорофилла
Образование хлорофилла осуществляется в 2 фазы: первая фаза — темновая, во время которой образуется предшественник хлорофилла — протохлорофилл, а вторая — световая, при которой из протохлорофилла на свету образуется хлорофилл.
Образование хлорофилла зависит как от вида растения, так и от ряда внешних условий. Некоторые растения, например проростки хвойных, могут позеленеть и без участия света, в темноте, но у большинства растений хлорофилл образуется из протохлорофилла только на свету.
В отсутствие света получаются этиолированные растения, имеющие тонкий, слабый, сильно вытянутый стебель и очень мелкие бледно-желтые листья. Если выставить этиолированные растения на свет, то листья быстро позеленеют. Это объясняется тем, что в листьях уже имеется протохлорофилл, который под воздействием света легко превращается в хлорофилл.
Большое влияние на образование хлорофилла оказывает температура; при холодной весне у некоторых кустарников листья не зеленеют до установления теплой погоды: при понижении температуры подавляется образование протохлорофилла.
Минимальной температурой, при которой начинается образование хлорофилла, является 2°, максимальной, при которой образование хлорофилла не происходит, 40°. Кроме определенной температуры, для образования хлорофилла необходимы элементы минерального питания, особенно железо.
При его отсутствии у растений наблюдается заболевание, называемое хлорозом. По-видимому, железо является катализатором при синтезе протохлорофилла, так как в состав молекулы хлорофилла оно не входит. Для образования хлорофилла также необходимы азот и магний, входящие в состав его молекулы. Важным условием является и наличие в клетках листа пластид, способных к позеленению.
При их отсутствии листья растений остаются белыми, растение не способно к фотосинтезу и может жить только до тех пор, пока не израсходует запасы семени. Это явление называется альбинизмом. Оно связано с изменением наследственной природы данного растения.
Количественные отношения между хлорофиллом и усваиваемой углекислотой
При большем содержании хлорофилла в растении процесс фотосинтеза начинается при меньшей интенсивности света и даже при более низкой температуре. С увеличением содержания хлорофилла в листьях фотосинтез возрастает, но до известного предела. Следовательно, нет прямой зависимости между содержанием хлорофилла и интенсивностью поглощения СО2.
Количество ассимилированного листом СО2 в час в пересчете на единицу содержащегося в листе хлорофилла тем выше, чем меньше хлорофилла. Р. Вильштеттером и А. Штолем была предложена единица, характеризующая соотношение между количеством хлорофилла и поглощенным углекислым газом.
Количество разложенной в единицу времени углекислоты, приходящееся на единицу веса хлорофилла, они назвали ассимиляционным числом.
Ассимиляционное число непостоянно: оно больше при малом содержании хлорофилла и меньше при высоком содержании его в листьях. Следовательно, молекула хлорофилла используется более продуктивно при низком его содержании в листе и продуктивность хлорофилла уменьшается с увеличением его количества. Данные введены в таблицу.
Таблица
Ассимиляционное число в зависимости от содержания хлорофилла
(по Р. Вильштеттеру и А. Штолю)
| Растения |
Содержание хлорофилла в 10г. листьев (мг) |
Ассимиляционное число |
|
Вяз: зеленая раса желтая раса |
16,2 1,2
|
6,9 82,0 |
| Сирень | 16,2 | 5,8 |
| Этиолированные проростки фасоли после освещения в течение:
6 часов 4 дней |
0,3 7,8 |
133,0 13,3 |
Изданных таблицы видно, что нет прямой зависимости между содержанием хлорофилла и количеством поглощенной СО2. Хлорофилл в растениях всегда содержится в избытке и, очевидно, не весь участвует в фотосинтезе. Это объясняется тем, что при фотосинтезе наряду с процессами фотохимическими, которые осуществляются при участии хлорофилла, есть процессы чисто химические, которым свет не нужен.
Темновые реакции в растениях протекают значительно медленнее, чем световые. Скорость световой реакции равна 0,00001 секунды, темновой — 0,04 секунды. Впервые темновые реакции в процессе фотосинтеза обнаружены Ф. Блэкманом.
Он установил, что темновая реакция зависит от температуры, и с повышением ее скорость темновых процессов увеличивается. Длительность световых реакций ничтожна, поэтому скорость процесса фотосинтеза определяется главным образом продолжительностью темновых процессов.
Иногда при благоприятных для фотосинтеза условиях (достаточное количество хлорофилла и света) он протекает медленно. Это объясняется тем, что продукты, образующиеся при фотохимических реакциях, не успевают перерабатываться при темновых. Малое количество хлорофилла позволяет всем образующимся продуктам в фотохимической реакции быстро и полностью перерабатываться при темновой реакции.
Источник: LibTime.ru