Гранулярный эндоплазматический ретикулум
Эндоплазматическую сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), удалось обнаружить только с появлением электронного микроскопа. ЭПС есть только в эукариотических клетках и представляет собой сложную систему мембран, образующих уплощенные полости и трубочки. Все вместе это выглядит как сеть. ЭПС относится к одномембранным органоидам клетки.

Мембраны ЭПС отходят от внешней мембраны ядра и по строению сходны с ней.
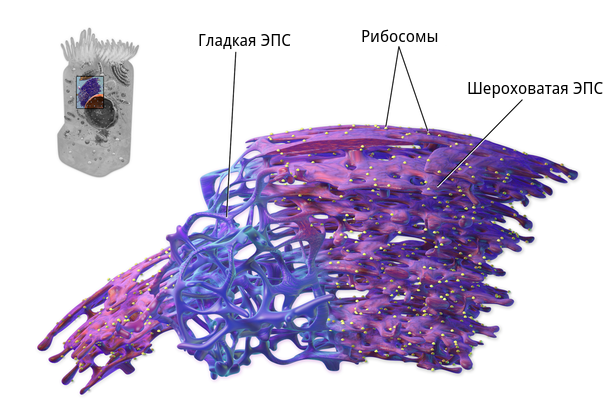
Эндоплазматическая сеть делится на гладкую (агранулярную) и шероховатую (гранулярную). Последняя усеяна прикрепленными к ней рибосомами (из-за этого и возникает «шероховатость»). Основная функция обоих типов связана с синтезом и транспортом веществ. Только шероховатая отвечает за белок, а гладкая — за углеводы и жиры.
По своему строению ЭПС представляет собой множество парных параллельных мембран, пронизывающих почти всю цитоплазму. Пара мембран образует пластинку (полость внутри имеет разную ширину и высоту), однако гладкая эндоплазматическая сеть в большей степени имеет трубчатое строение. Такие уплощенные мембранные мешочки называют цистернами ЭПС.
Рибосомы, расположенные на шероховатой ЭПС, синтезируют белки, которые поступают в каналы ЭПС, созревают (приобретают третичную структуру) там и транспортируются. У таких белков сначала синтезируется сигнальная последовательность (состоящая преимущественно из неполярных аминокислот), конфигурация которой соответствует специфическому рецептору ЭПС. В результате рибосома и эндоплазматическая сеть связываются. При этом рецептор образует канал для перехода синтезируемого белка в цистерны ЭПС.
После того, как белок оказывается в канале эндоплазматического ретикулума сигнальная последовательность от него отделяется. После этого он свертывается в свою третичную структуру. При транспортировке по ЭПС белок приобретает ряд других изменений (фосфорилирование, образование связи с углеводом, т. е. превращение в гликопротеин).
Большинство белков, оказавшихся в шероховатой ЭПС, далее попадают в аппарат (комплекс) Гольджи. Оттуда белки либо секретируются из клетки, либо поступают в другие органоиды (обычно лизосомы), либо откладываются как гранулы запасных веществ.
Следует иметь в виду, что не все белки клетки синтезируются на шероховатой ЭПС. Часть (обычно меньшая) синтезируется свободными рибосомами в гиалоплазме, такие белки используются самой клеткой. У них сигнальная последовательность не синтезируется за ненужностью.
Основной функцией гладкой эндоплазматической сети является синтез липидов (жиров). Например, ЭПС эпителия кишечника синтезирует их из жирных кислот и глицерола, всасывающихся из кишечника. Затем липиды попадают в комплекс Гольджи. Кроме клеток кишечника, гладкая ЭПС хорошо развита в клетках, секретирующих стероидные гормоны (стероиды относятся к липидам). Например, в клетках надпочечников, интерстициальных клетках семенников.
Синтез и транспорт белков, жиров и углеводов не единственные функции ЭПС. В печение эндоплазматический ретикулум участвует в процессах детоксикации. Особая форма гладкой ЭПС — саркоплазматический ретикулум – присутствует в мышечных клетках и обеспечивает сокращение за счет перекачки ионов кальция.
Структура, объем и функциональность эндоплазматической сети клетки не является постоянной на протяжении клеточного цикла, а подвержены тем или иным изменениям.
Источник: biology.su
История открытия
Впервые эндоплазматический ретикулум был обнаружен американским учёным К. Портером в 1945 году посредством электронной микроскопии.
Строение
Эндоплазматический ретикулум состоит из разветвлённой сети трубочек и карманов, окружённых мембраной. Площадь мембран эндоплазматического ретикулума составляет более половины общей площади всех мембран клетки.
Мембрана ЭПР морфологически идентична оболочке клеточного ядра и составляет с ней одно целое. Таким образом, полости эндоплазматического ретикулума открываются в межмембранную полость ядерной оболочки. Мембраны ЭПС обеспечивают активный транспорт ряда элементов против градиента концентрации. Нити, образующие эндоплазматический ретикулум имеют в поперечнике 0,05-0,1 мкм (иногда до 0,3 мкм), толщина двухслойных мембран, образующих стенку канальцев составляет около 50 ангстрем (5 нм, 0.005 мкм). Эти структуры содержат ненасыщенные фосфолипиды, а также некоторое количество холестерина и сфинголипидов. В их состав также входят белки.
Трубочки, диаметр которых колеблется в пределах 0.1-0.3 мкм, заполнены гомогенным содержимым. Их функция — осуществление коммуникации между содержимым пузырьков ЭПС, внешней средой и ядром клетки.
Эндоплазматический ретикулум не является стабильной структурой и подвержен частым изменениям.
Выделяют два вида ЭПР:
- гранулярный эндоплазматический ретикулум
- агранулярный (гладкий) эндоплазматический ретикулум
На поверхности гранулярного эндоплазматического ретикулума находится большое количество рибосом, которые отсутствуют на поверхности агранулярного ЭПР.
Гранулярный и агранулярный эндоплазматический ретикулум выполняют различные функции в клетке.
Функции эндоплазматического ретикулума
При участии эндоплазматического ретикулума происходит трансляция и транспорт белков, синтез и транспорт липидов и стероидов. Для ЭПС характерно также накопление продуктов синтеза. Эндоплазматический ретикулум принимает участие в том числе и в создании новой ядерной оболочки (например после митоза). Эндоплазматический ретикулум содержит внутриклеточный запас кальция, который является, в частности, медиатором сокращения мышечной клетки. В клетках мышечных волокон расположена особая форма эндоплазматического ретикулума — саркоплазматическая сеть.
Функции агранулярного эндоплазматического ретикулума
Агранулярный эндоплазматический ретикулум участвует во многих процессах метаболизма. Ферменты агранулярного эндоплазматического ретикулума участвуют в синтезе различных липидов и фосфолипидов, жирных кислот и стероидов. Также агранулярный эндоплазматический ретикулум играет важную роль в углеводном обмене, обеззараживании клетки и запасании кальция. В частности, в связи с этим в клетках надпочечников и печени преобладает агранулярный эндоплазматический ретикулум.
Синтез гормонов
К гормонам, которые образуются в агранулярном ЭПС, принадлежат, например, половые гормоны позвоночных животных и стероидные гормоны надпочечников. Клетки яичек и яичников, ответственные за синтез гормонов, содержат большое количество агранулярного эндоплазматического ретикулума.
Накопление и преобразование углеводов
Углеводы в организме накапливаются в печени в виде гликогена. Посредством гликолиза гликоген в печени трансформируется в глюкозу, что является важнейшим процессом в поддержании уровня глюкозы в крови. Один из ферментов агранулярного ЭПС отщепляет от первого продукта гликолиза, глюкоза-6-фосфата, фосфогруппу, позволяя таким образом глюкозе покинуть клетку и повысить уровень сахаров в крови.
Нейтрализация ядов
Гладкий эндоплазматический ретикулум клеток печени принимает активное участие в нейтрализации всевозможных ядов. Ферменты гладкого ЭПР присоединяют встретившиеся молекулы активных веществ, которые таким образом могут быть растворены быстрее. В случае непрерывного поступления ядов, медикаментов или алкоголя, образуется большее количество агранулярного ЭПР, что повышает дозу действующего вещества, необходимую для достижения прежнего эффекта.
Саркоплазматический ретикулум
Особую форму агранулярного эндоплазматического ретикулума, саркоплазматический ретикулум, образует ЭПС в мышечных клетках, в которых ионы кальция активно закачиваются из цитоплазмы в полости ЭПР против градиента концентрации в невозбуждённом состоянии клетки и освобождаются в цитоплазму для инициации сокращения. Концентрация ионов кальция в ЭПС может достигать 10−3 моль, в то время как в цитозоле порядка 10−7 моль (в состоянии покоя). Таким образом, мембрана саркоплазматического ретикулума обеспечивает активный перенос против градиентов концентрации больших порядков. И приём и освобождение ионов кальция в ЭПС находится в тонкой взаимосвязи от физиологических условий.
Концентрация ионов кальция в цитозоле влияет на множество внутриклеточных и межклеточных процессов, таких как: активация или торможение ферментов, экспрессия генов, синаптическая пластичность нейронов, сокращения мышечных клеток, освобождение антител из клеток имунной системы.
Функции гранулярного эндоплазматического ретикулума
Гранулярный эндоплазматический ретикулум имеет две функции: синтез белков и производство мембран.
Синтез белков
Белки, производимые клеткой, синтезируются на поверхности рибосом, которые могут быть присоединены к поверхности ЭПС. Полученные полипептидные цепочки помещаются в полости гранулярного эндоплазматического ретикулума (куда попадают и полипептидные цепочки, синтезированные в цитозоле), где впоследствии правильным образом обрезаются и сворачиваются. Таким образом, линейные последовательности аминокислот получают после транслокации в эндоплазматический ретикулум необходимую трёхмерную структуру, после чего повторно перемещаются в цитозоль.
Синтез мембран
Рибосомы, прикреплённые на поверхности гранулярного ЭПР, производят белки, что, наряду с производством фосфолипидов, среди прочего расширяет собственную поверхность мембраны ЭПР, которая посредством транспортных везикул посылает фрагменты мембраны в другие части мембранной системы.
Источник: dic.academic.ru
Аппарат Гольджи (комплекс Гольджи, пластинчатый комплекс).
Комплекс Гольджи представляет собой скопление мембранных структур в виде стопки (диктиосома). Между стопками располагаются тонкие прослойки гиалоплазмы. В секретирующих клетках аппарат Гольджи обычно поляризован: с одной стороны мембранные мешочки непрерывно образуются (цис-участок), а с другой – происходит отделение вакуолей (транс-участок). Цистерны аппарата Гольджи связаны с канальцами ЭПР. В некоторых клетках аппарат Гольджи имеет вид сложных сетей.
Функции аппарата Гольджи:
-
Синтез полисахаридов, их взаимосвязь с белками, приводящая к образованию гликопротеинов (например, гликокаликса);
-
Модификация белков в аппарате Гольджи. Белки по мере движения по цистернам аппарата Гольджи «созревают», т.е. подвергаются модификациям: некоторые их аминокислоты фосфорилируются, ацетилируются и др. Подвергаются модификации и олигосахаридные цепи белков. При этом возникает специальный комплекс олигосахаридов;
-
Выведение готовых секретов за пределы клетки. Синтезированный на рибосомах и модифицированный в АГ экспортируемый белок упаковывается в вакуоли на транс-участке диктиосомы. Такие вакуоли движутся к поверхности клетки, соприкасаются с плазматический мембраной и сливаются с ней (экзоцитоз). Таким образом, содержимое вакуолей оказывается за пределами клетки;
-
Сортировка белков в аппарате Гольджи.
Лизосомы.
Представляют собой пузырьки, ограниченные одиночной мембраной с разнородным содержимым внутри. Все лизосомы содержат ферменты гидролазы (известно примерно 40 видов), что позволяет им участвовать в процессах внутриклеточного переваривания. По морфологии можно выделить первичные, вторичные лизосомы, аутофагосомы и телолизосомы (остаточные тельца).
Первичные лизосомы – мелкие мембранные пузырьки 50-100 нм, содержащие набор гидролаз. Это неактивные структуры, еще не вступившие в процессы расщепления субстратов.
Вторичные лизосомы – продукт слияния первичных лизосом с фагоцитарными, или пиноцитозными вакуолями. При этом гидролазы первичной вакуоли получают доступ к субстратам, которые они начинают расщеплять.
Аутофагосомы (аутолизосомы) – вторичные лизосомы, выполняющие функцию уничтожения измененных, либо отслуживших свой срок клеточных компонентов.
Телолизосомы (остаточные тельца) – вторичные лизосомы, содержащие не до конца переваренные продукты обмена, либо пигментные вещества.
Источник: StudFiles.net
История открытия
Впервые эндоплазматический ретикулум был обнаружен канадскими учеными Китом Портером, Альбертом Клодом и Эрнстом Фулламом в 1945 году с помощью электронной микроскопии.
Строение
Эндоплазматический ретикулум состоит из разветвленной сети трубочек и карманов, окруженных мембраной. Площадь мембран эндоплазматического ретикулума составляет более половины общей площади всех мембран клетки.
Мембрана ЭПР морфологически идентична оболочке клеточного ядра и составляет с ней одно целое. Таким образом, полости эндоплазматического ретикулума открываются в межмембранного полость ядерной оболочки. Мембраны ЭПС обеспечивают активный транспорт ряда элементов против градиента концентрации. Пузырьки и канальцы, создают эндоплазматический ретикулум, имеют в поперечнике 0,05-0,1 микрона (иногда до 0,3 микрона), толщина двухслойных мембран, стенку канальцев, составляет около 50 ангстрем. Эти структуры содержат ненасыщенные фосфолипиды, а также некоторое количество холестерина и сфинголипидов. В их состав также входят белки. Тончайшие трубочки, диаметр которых колеблется в пределах 1000-3000 ангстрем, заполненные гомогенным содержимым и соединяют большие по размеру части эндоплазматического ретикулума. Подобно компонентов цитоскелета мембраны полярные: с одного конца они наращиваются, а с другой распадаются на отдельные фрагменты. Эндоплазматический ретикулум не является стабильной структурой и подвержен частым изменениям. Выделяют два типа ЭПР:
- Шершавый (гранулярный) эндоплазматический ретикулум,
- Гладкий (агранулярный) эндоплазматический ретикулум.
На поверхности шероховатого эндоплазматического ретикулума находится большое количество рибосом, которые отсутствуют на поверхности гладкого ЭПР.
Шершавый и гладкий эндоплазматический ретикулум выполняют некоторые различные функции в клетке.
Функции эндоплазматического ретикулума
При участии эндоплазматического ретикулума происходит трансляция и транспорт мембранных белков, сектеруються, синтез и транспорт липидов и стероидов. Для ЭПС характерно также накопление продуктов синтеза. Эндоплазматический ретикулум принимает участие в том числе и в создании новой ядерной оболочки (например после митоза). Эндоплазматический ретикулум содержит внутриклеточный запас кальция, который является медиатором многих реакций ответы клетки, в частности сокращение мышечных клеток. В клетках мышечных волокон расположена особая форма эндоплазматического ретикулума — саркоплазматическая ретикулум.
Функции гладкого эндоплазматического ретикулума
Гладкий эндоплазматический ретикулум участвует во многих процессах метаболизма. Ферменты гладкого эндоплазматического ретикулума участвуют в синтезе липидов и фосфолипидов, жирных кислот и стероидов. Также агранулярный эндоплазматический ретикулум играет важную роль в углеводном обмене, обеззараживании клетки и запасании кальция. В частности, в связи с этим в клетках надпочечников и печени преобладает гладкой эндоплазматический ретикулум.
Синтез гормонов
К гормонам, которые образуются в гладком ЭПС, относятся, например, половые гормоны позвоночных животных и стероидные гормоны (синтезируемых в надпочечниках). Клетки яичек и яичников, отвечающих за синтез гормонов, содержат большое количество гладкого ЭПР.
Накопление и преобразование углеводов
Углеводы в организме накапливаются в печени в виде гликогена. С помощью гликолиза гликоген в печени трансформируется в глюкозу, которая является важнейшим процессом в поддержании уровня глюкозы в крови. Один из ферментов гладкого ЭПС отщепляет от первого продукта гликолиза, глюкоза-6-фосфата, фосфогрупу, позволяя, таким образом, глюкозе оставить клетку и повысить уровень сахара в крови.
Нейтрализация ядов
Гладкий эндоплазматический ретикулум клеток печени принимает активное участие в нейтрализации токсинов. Ферменты гладкого ЭПР присоединяют к нерастворимых радикалов гидрофильные последовательности, в результате чего токсин становится лучше растворимым и может выводиться из клетки, а затем и из организма через мочу. В случае непрерывного поступления ядов, медикаментов или алкоголя, площадь гладкого ЭПР увеличивается, что позволяет нейтрализовать большее количество токсина за счет увеличения рабочей поверхности ЭПР.
Саркоплазматическому ретикулум
Особая форма гладкого эндоплазматического ретикулума, саркоплазматическая ретикулум, образуется в мышечных клетках, где ионы кальция активно закачиваются из цитоплазмы в полости ЭПР против градиента концентрации в невозбужденном состоянии клетки и освобождаются в цитоплазму для инициации сокращения. Концентрация ионов кальция в ЭПС может достигать 10 -3 моль / л, тогда как в цитоплазме — порядка 10 -7 моль / л (в состоянии покоя). Таким образом, мембрана саркоплазматического ретикулума обеспечивает активный перенос против градиентов концентрации больших порядков. И прием и увольнение ионов кальция в ЭПС находится в жесткой зависимости от физиологических условий. 2
Концентрация ионов кальция в цитозоле влияет на множество внутриклеточных и межклеточных процессов, таких как включение и ингибирование ферментов, экспрессия генов, синаптическая пластичность нейронов, сокращения мышечных клеток, освобождение антител из клеток имунной системы.
Функции гранулярного эндоплазматического ретикулума
Гранулярий эндоплазматический ретикулум имеет две функции: синтез белков и производство мембран.
Синтез белков
Белки, вырабатываемые клеткой, синтезируются в рибосомах, которые могут быть присоединены к поверхности ЭПС, именно это и придает поверхности «шершавый», гранулярный вид. На поверхности ЭПР синтезируется много белков, предназначенных для сортировки и транслокации в определенных отделов клетки, то есть интегральные мембранные белки, белки органелл и белки, предназначенные для секреции. Однако, рибосомы на поверхности ЭПР на всегда находятся там, а присоединяются к нему после начала синтеза белка, предназначенного для сортировки в ЭПР. Свободная рибосома синтезирует белок, пока цитозольные частицы распознавания сигнала на узнают сигнал с 5-15 гидрофобных аминокислот (N-сигнальная последовательность) после положительно зярядженои аминокислоты в начале цепочки. Этот сигнальный фрагмент остается в мембране до конца синтеза, после чего он отрезается от готового белка.
Полученные полипептидной цепи помещаются в полости гранулярного эндоплазматического ретикулума, где впоследствии подвергаются первым шагам посттрансляционной модификации и сворачиваются. Таким образом, линейные цепи аминокислот получают после транслокации в эндоплазматический ретикулум необходимую трехмерную структуру.
Синтез мембран
С помощью производства фосфолипидов, ЭПР расширяет поверхность своих собственных мембран, с помощью транспортных везикул транспортируются в другие части мембранной системы.
Источник: info-farm.ru
Гранулярная эндоплазматическая сеть
Гранулярный (шерховатый) эндоплазматический ретикулум производит мембраны и секреторные белки. Рибосомы, прикрепленные к гранулярной ЭПР, синтезируют белки в процессе трансляции. В некоторых лейкоцитах (белых кровяных клетках) шероховатый ЭПР продуцирует антитела. В клетках поджелудочной железы он продуцирует инсулин.
Гранулярный и агранулярный ЭПР, как правило, взаимосвязаны, а белки и мембраны, продуцируемые шероховатым ЭПР, перемещаются в гладкий ЭПР. Некоторые белки отправляются на аппарат Гольджи специальными транспортными везикулами. После того, как белки были модифицированы в Гольджи, они транспортируются в надлежащие пункты назначения внутри клетки или экспортируются из клетки путем экзоцитоза.
Агранулярная эндоплазматическая сеть
Агранулярный (гладкий) эндоплазматический ретикулум обладает широким спектром функций, включая синтез углеводов и липидов. Липиды, такие как фосфолипиды и холестерин, необходимы для создания клеточных мембран. Гладкий ЭПР также служит переходной областью для везикул, которые транспортируют продукты эндоплазматической сети в различные пункты назначения.
В клетках печени агранулярный ЭПР продуцирует ферменты, помогающие детоксифицировать определенные соединения. В мышцах он помогает в сокращении мышечных клеток, а в клетках мозга синтезирует мужские и женские гормоны.
Источник: natworld.info