Схематически изобразите клеточный цикл
Клеточные циклы у эукариот качественно не различаются у разных видов и в клетках разных тканей одного вида. Замечены различия, главным образом, в длительности цикла. Среди высших эукариот некоторые клетки делятся через 10 минут, другие через 3 часа, третьи — через 200 часов.
Клеточный цикл у большинства соматических клеток высших эукариот подразделяют на 4 стадии: G1 (gap1, предсинтетический период, или период подготовки к синтезу ДНК), S (synthesis, период синтеза ДНК), G2 (gар 2, постсинтетический период подготовки к клеточному делению) и М (mitosis, собственно процесс клеточного деления). Иногда выделяют G0 — стадию между М и G1 или стадию покоя. В культуре клеток человека весь цикл занимает примерно 24 ч. при этом на стадии G1, S, G2 и М приходится 10, 9, 4 и 1 ч соответственно. Фазы G1, S и G2 вместе составляют интерфазу. Наиболее детальные сведения получены при изучении клеточных циклов дрожжей. Данные генетических и молекулярных исследований показали, что клеточные циклы включают ряд этапов, на которых осуществляется контроль продвижения клетки от одной фазы к другой — точки проверки или чекпойнты (checkpoints). Первая стадия проверки у дрожжей называется START, у млекопитающих — G1-checkpoint. Если клетка не выросла до необходимых размеров и окружающая среда недостаточно хороша; клетка будет оставаться в G1, т. е. не будет сигнала к синтезу ДНК (S-период).
В S-фазе разные участки генома реплицируются в разное время. В культуре клеток человека сначала синтезируется ДНК, которая выявляется в R-бэндах метафазных хромосом, обогащенных генами. В конце S-периода синтезируется ДНК G-бэндов. Полагают, что между этими отрезками S-периода также существует стадия проверки (проверка целостности ДНК, checkpoint DNA intregritу).
Стадия проверки G2 находится на границе G2 и М. Если не завершилась репликация всей ДНК, если клетка не выросла до нормальных размеров и окружающая среда недостаточно хороша, клетка не способна перейти к стадии М.
Третья проверка происходит в течение фазы М: хромосомы должны быть надежно прикреплены к нитям митотического веретена,
чтобы началось разделение хроматид. Схематически клеточный цикл показан на рис….

Рис…. Схема клеточного цикла эукариот.
Нарушения регуляции клеточного цикла и чекпойнт-контроля часто приводят к геномной нестабильности и предрасположенности к раку.
нимание молекулярных механизмов, которые регулируют эти процессы важно как для прикладной медицины, так и для фундаментальных научных исследований. Клеточный цикл проходит одинаково во всех эукариотических клетках, поэтому изучение его у различных организмов создает и наше общее понимание того, как различные события клеточного цикла контролируются и координируются у человека. Успешное деление зависит от того, насколько высоко скоординировано прошло удвоение и сегрегация клеточных компонентов. Координация завершается постановкой осуществления событий клеточного цикла под контроль высоко регулируемых протеинкиназ, известных как циклин-зависимые киназы.
Регуляция клеточного цикла у эукариот.
Различные клеточные процессы, необходимые для успешной репликации и деления клеток, управляются последовательной активаций и дезактивацией семейства циклин-зависимых киназ (Cdks). Активация преимущественно управляется периодической экспрессией циклинов и нуждается в активирующем фосфорилировании киназы. Инактивация контролируется во-первых, ингибирующим фосфорилированием киназ, во-вторых убиквитин-опосредованной деградацией циклинов и в-третьих -взаимодействием всего комплекса с малыми ингибирующими белками.
Основные принципы, положенные в основу регуляции прохождения клетки по циклу таковы:
1.Активация Cdks управляется последовательно экспрессией и объединением с циклинами.
2.Активность каждой пары Cdk-циклин необходима для активации последующей.
3.Разрушение циклинов приводит к однонаправленному клеточному циклу
4.Ингибирование комплексов Cdks-циклины путем фосфорилирования или путем связывания с белками-ингибиторами задерживает активацию Cdks и замедляет движение клетки по циклу в неблагоприятных условиях.
Как это следует из названия, Cdks являются протеин-киназами, которые для своей активации должны связаться с соответствующими циклинами. Процесс синтеза и деградации циклинов высоко координирован, в то время как основной уровень контроля Cdks заключается в периодичности присутствия или отсутствия циклиновой субъединицы. Данные об уровне экспрессии циклинов в течение клеточного цикла приведены на рис. ….
Рис….Экспрессия циклинов втечение клеточного цикла
Если покоящиеся клетки стимулировать к вхождению в клеточный цикл, то первым будет экспрессирован циклин D, который может объединяться с Cdk4 и Cdk6. Этот комплекс входит в ядро, где фосфорилирует белок ретинобластомы Rb и два других покет-белка р107 и р130. Фосфорилировани Rb приводит к высвобождению семейства транскрипционных факторов E2F и таким образом способствует экспрессии белков, необходимых для G1 и S-фаз клеточного цикла. Схема работы белка Rb показана на рис…..
Рис…. Схема фосфорилирования белка Rb
Тот же митогенный сигнал, который вызвал экпрессию циклина D, также вызывает и экспрессию второго циклина – Е, и двух ингибиторов Cdks – р21cip1 и р27kip1. Экспрессия белков-ингибиторв в тот момент, когда клетка входит в новый цикл роста и деления кажется контрпродуктивной. Тем не менее, р21 и р27 связываются с коплексом циклин D- Cdk4 не ингибируя его киназную активность и на самом деле оказываются необходимыми для создания этого комплекса и его транспорта в ядро. Напротив, р21 и р27 являются эффективным ингибитором активности комплекса циклин Е- Cdk2. Таким образом, присутствие этих белков в ранней G1-фазе способствует образованию комплексов циклин D- Cdk4 и, в то же время, задерживает активацию циклин Е- Cdk2 комплексов.
Циклин Е- Cdk2 взаимодействует с циклин D- Cdk4/6 в фосфорилировании и инактивации Rb и покет- белков. Двойное фосфорилирование Rb необходимо для полной активации транскрипционной программы S-фазы. Таким образом ограниченное время движение клетки по циклу происходит независимо от внеклеточных сигналов. Внутренние или внешние события могут замедлить или предотвратить продвижение клетки по циклу, но если никакого останавливающего это движение сигнала получено не будет, то инициированный клеточный цикл, включая рост, репликацию, сегрегацию и деление будет продолжаться сам по себе без дальнейших внеклеточных посылов.
Активация циклин Е- Cdk2 ведет к инициации репликации ДНК, причем описано большое число белков, необходимых для осуществления задачи удвоения и сегрегации клеточного содержимого.
клин Е- Cdk2 также фосфорилирует белок-ингибитор р27. Это делает р27 мишенью для распознавания убиквитин-лигазой, а убиквитинированная форма идет путем протеосомной деградации. Таким образом циклин Е- Cdk2 включает диструкцию своего отрицательного регулятора. Циклин Е- Cdk2 также автофосфорилируется по циклину Е, что тоже делает его мишенью для деградации. Это свойство подготавливать к деградации как ингибитор, так и сам циклин делает киназный комплекс одновременно самоактивирующим и самоограничивающим.
Циклин А экспрессируется сразу же после циклина Е на границе G1 и S фаз. Активность обоих комплексов — циклин Е- Cdk2 и циклин А- Cdk2 – для инициации и правильного протекания ДНК-репликации, а также для гарантии того, что репликация проходит в течение одного клеточного цикла только один раз. К тому же циклин А- Cdk2 способствует эффективному протеканию S-фазы, повышая транскрипцию гистоновых и других генов, необходимых для согласованной репликации.
После роста и удвоения клетки оказываются перед необходимостью делиться на две жизнеспособные дочерние клетки. Все завершается митозом, который начинается профазой с конденсацией хромосом и формированием веретена деления. В прометафазе микротрубочки веретена прикрепляются к кинетохорам двух сестринских хроматид, а оболочка ядра растворяется. Если все произошло правильно, то в анафазе веретено растаскивает сестринские хроматиды в разные стороны.
лед за разделением удвоенного генома в телофазе деконденсируются хроматиды и начинают образовываться новые ядерные оболочки. Наконец, при цитокинезе разделением цитопоазмы завершается образование двух дочерних клеток. Эти драматические морфологические изменения происходят под контролем Cdk1 (Cdc2), ассоциированной с циклинами А и В. Экспрессия циклина В запаздывает по отношению к экспрессии циклина А, возрастая к поздней S-фазе и сохраняя высокий уровень в G2 и М. Так как комплекс циклин В- Cdk1 преимущественно пребывает в неактивной фосфорилированной форме, то постепенное возрастание количества циклина В не сопровождается таким же постепенным возрастанием киназной активности. Дефосфорилирование и активация циклин В- Cdk1 строго коррелирует с морфологическими изменениями, сопровождающими митоз. Ядерные ламины, белки ядрышек, центросомные белки и Eg5 (белок, связанный с движением кинезинов) являются субстратами циклин В- Cdk1 комплекса. Многокопийные органеллы, такие как митохондрии, распределены по цитоплазме равномерно, так, что дочерние клетки наследуют их вероятностно, случайным образом. С другой стороны, органеллы, существующие в единственном числе, такие как аппарат Гольджи, должны быть разделены активно. Cdk1 при ассоциации с другой субъединицей циклина В – В2, локализуясь преимущественно в зоне эндоплазматического ретикулума может играть ведущую роль при диспергировании аппарата Гольджи и гарантировать его разделение при цитокинезе таким образом, чтобы в обеих дочерних клетках оказались компоненты, достаточные для построения заново этого секреторного комплекса. Разные стадии митоза характеризуются очень быстрым и скоординированным исчезновением циклинов А и В. Разрушение митотических циклинов необходимо для того, чтобы гарантировать переход клеток в интерфазу до инициации следующего раунда ДНК-репликации. Это разрушение опосредуется anaphase promoting complex.
Белки-ингибиторы комплексов циклин-зависмых киназ с циклинами.
Важнейшим механизмом негативной регуляции комплексов циклин – Cdk является взаимодествие с малыми белками-ингибиторами. У млекопитающих описано два различных семейства ингибиторов Cdk — CKIs. Семейство Ink4 (р15, р16, р18 и р19), названные так по способности ингибировать Cdk4, связываются с мономерами Cdk4 и Cdk6. Все они представлены на рис….
Рис… Ингибиторы комплексов циклин-CDKs
Ингибиторы связываются с ними в той же области, что и циклин D. Таким образом высокая экспрессия Ink4 блокирует образование активных комплексов циклин D –Cdk4. Так как Ink4 препятствуют образованию комплексов циклин D –Cdk4, то они увеличивают пул р27, способного связываться с комплексами циклин Е – Cdk2 и ингибировать их. Следовательно, ап-регуляция Ink4 напрямую ингибирует активность комплексов циклин D – Cdk4 и опосредованно — активность комплексов циклин Е –Cdk2. Второе семейство CKIs, Cip/Kip семейство (р21cip, p27kip и р57kip) ингибируют комплексы, содержащие Cdk2. Это семейство ингибиторов связывается с формирующимися комплексами, блокируя таким образом доступ к субстрату. Р21, первый из этих ингибиторов, является белком, экспрессия которого активируется р53 в ответ на повреждение ДНК.
Регуляция активности CDKS фосфорилированием.
Ассоциация киназы с циклином приводит к частичной активации киназного комплекса. Полная активация требует фосфорилирования по треонину внутри в районе Т-петли киназы. Киназы, способные к такому активирующему фосфорилированию называются Cdk-activating kinases (Cak). Активность самих Cak постоянна в течение клеточного цикла и, вероятно, небольшие ее модуляции влияют на движение по циклу.
Фосфорилирование Cdks также негативно регулирует их киназную активность. Активность Cdk1-циклин В является наиболее полно охарактеризованным примером ингибирующего фосфорилирования. Впрочем, активность циклин А –Cdk2, циклин Е – Cdk2 и циклин D – Cdk4 комплексов тоже регулируется этим способом. Ингибиторное фосфорилирование осуществляется рядом с N-концом с АТР-связывающим сайтом киназной субъеиницы, у человеческой Cdk1 это треонин-14 и тирозин-15. Фосфорилирование по тирозину-15 в основном осуществляется ядерной киназой Wee1, в то время как Myt1, цитоплазматический белок, связанный с мембранами цитоплазматического ретикулума и Гольджи, ведет фосфорилирование по треонину-14.
зличная локализация этих киназ предполагает механизм, по которому Cdk1 может негативно регулироваться как в ядре, так и в цитоплазме. Ингибирующие фосфорилирование Cdks снимается семейством Cdc25 фосфатаз двойной специфичности. В клетках млекопитающих экспрессируются три формы Cdc25. Cdc25А действует в начале клеточного цикла, способствуя дефосфорилированию и активации циклин Е –Cdk2. Тогда как Cdc25В и Cdc25С обе принимают участие в переходе G2-M. Любопытно, что сами Cdc25 являются субстратами для Cdks: Cdc25А фосфорилируется и активируется циклин Е –Cdk2 , а Cdc25С — циклин В –Cdk1. Это предполагает, что быстрая и полная активация Cdks стимулируется по принципу положительной обратной связи.
Источник: studopedia.ru
Биологическое значение деления клеток. Новые клетки возникают в результате деления уже существующих. Если делится одноклеточный организм, то из него образуются два новых, то есть деление клеток обеспечивает непрерывность жизни.
Многоклеточный организм также начинает свое развитие чаще всего с одной-единственной клетки. Путем многократных делений образуется огромное количество клеток, которые и составляют организм.
Жизненный, или клеточный, цикл — это жизнь клетки от момента ее появления в процессе деления материнской клетки и до ее собственного деления, включая это деление, или гибели. В течение этого цикла клетка растет, видоизменяется таким образом, чтобы успешно выполнять свои функции в организме (этот процесс называется дифференцировкой клетки), затем она выполняет свои функции в течение определенного времени, по истечении которого делится, образуя новые клетки, или отмирает.
У различных видов организмов клеточный цикл занимает разное время: у бактерий — около 20 мин, у инфузории-туфельки — от 10 до 20 ч. Клетки тканей многоклеточных организмов на ранних стадиях развития делятся очень часто, а затем клеточные циклы значительно удлиняются. Например, сразу после рождения животных нервные клетки головного мозга делятся часто: 80 % головного мозга формируется именно тогда. Однако большинство из этих клеток быстро теряет способность к делению, и часть из них доживает не делясь до естественной смерти животного.
Обязательным компонентом[VV88] каждого клеточного цикла является митотический цикл, который включает в себя подготовку клетки к процессу деления и митоз (рис. ).
Интерфаза — это процесс подготовки клетки к делению. Она состоит из трех периодов: пресинтетического, синтетического и постсинтетического.
Пресинтетический период (G) — наиболее продолжительная часть интерфазы. Он может продолжаться у различных видов клеток от 2 — 3 ч до нескольких суток. Этот период следует сразу же за предшествующим делением[VV89] , во время него клетка растет, накапливая энергию и вещества для последующего удвоения ДНК.
Синтетический период (S), который обычно длится 6 — 10 ч, включает в себя удвоение ДНК, синтез белков, необходимых для формирования хромосом, а также увеличение количества РНК. К концу этого периода каждая хромосома уже состоит из двух идентичных хроматид, соединенных друг с другом в области центромеры. В этот же период удваиваются центриоли[VV90] .
Удвоение молекулы ДНК называют также репликацией или редупликацией. Во время репликации часть молекулы «материнской» ДНК раскручивается на две нити с помощью специального фермента (рис. ), причем это достигается разрывом водородных связей между комплементарными азотистыми основаниями. Далее к каждому нуклеотиду разошедшихся нитей ДНК фермент ДНК-полимераза подстраивает комплементарный ему нуклеотид. Таким образом, образуются две двуцепочечные молекулы ДНК, в состав каждой из которых входят одна цепочка «материнской» молекулы и одна синтезированная («дочерняя») цепочка. Такой способ удвоения был назван полуконсервативным. Образовавшиеся две молекулы ДНК абсолютно идентичны.
Раскрутить для репликации сразу всю длинную молекулу ДНК невозможно. Поэтому репликация начинается сразу в нескольких местах молекулы ДНК. При этом синтезируются несколько коротких фрагментов «дочерней» нити, которые при помощи ферментов лигаз сшиваются в единую длинную молекулу.
Удвоение ДНК — сложный процесс, протекающий с участием нескольких ферментов и требующий затрат энергии.
Доказательства участия ферментов в синтезе ДНК были получены американским ученым А. Корнбергом. В 1956 г. ему удалось осуществить синтез ДНК в пробирке, добавляя туда матрицу — ДНК, «строительный материал» — нуклеотидтрифосфаты и экстракт белков из бактерий. Затратив восемь лет труда и переработав сотни килограмм бактерий, Корнберг получил в чистом виде фермент, осуществляющий матричный синтез ДНК — ДНК-полимеразу.
Для репликации нужно сначала расплести двойную спираль ДНК. Это тоже делают специальные ферменты — геликазы, разрывающие водородные связи между основаниями. Но расплетенные участки более чувствительны к повреждающим факторам — и к облучению, и к многим химическим веществам, вызывающим мутации. Чтобы они оставались в незащищенном состоянии как можно меньше, синтез на обеих цепочках идет одновременно.
Новые нуклеотиды присоединяются только к гидроксилу третьего атома рибозы[VV91] . Цепи и ДНК, и РНК могут удлиняться лишь таким способом, т. е. только в одном направлении.
Построение новой полинуклеотидной цепочки происходит в соответствии с принципом комплементарности: напротив аденина будет встраиваться тимин, напротив гуанина — цитозин и т. п. Тут, правда, возникает проблема, поскольку цепь может наращиваться только с одного конца. Когда цепь ДНК расплетается, на одной из цепочек оказывается свободным 3′-конец, а на другой — 5′-конец. Молекула ДНК-полимеразы на первой цепочке идет сразу вслед за расплетающими ферментами и синтезирует свою цепочку без всяких проблем. На этой цепочке новые нуклеотиды просто постоянно добавляются, и синтез продвигается вперед. Но вторая цепь направлена в другую сторону! Как же быть?
Оказывается, на ней синтез ДНК идет прерывисто (рис. ). Когда молекула ДНК достаточно расплетется, молекула ДНК-полимеразы садится на некоторую точку расплетенного участка второй цепочки и синтезирует комплементарную ей цепь ДНК в направлениипротивоположном лидирующей цепи. Эта молекула ДНК-полимеразы двигается не к месту расплетания, как на первой цепочке, а напротив, к свободному концу. После того, как молекула ДНК расплетется еще сильнее, новая молекула ДНК-полимеразы садится на новую точку расплетенного участка и двигается к синтезированному ранее участку. Нуклеотиды, как и положено, присоединяются в нужном направлении, образуя новые коротенькие цепочки [VV92] (их называют «фрагменты Оказаки»). Когда ДНК-полимераза достроит один фрагмент и «упрется» в синтезированный ранее другой, она отделится от ДНК. Затем она (или другая молекула ДНК-полимеразы) сможет опять сесть на вновь расплетенный участок и построить еще один фрагмент Оказаки. Отдельные фрагменты Оказаки соединяются друг с другом особым ферментом — ДНК-лигазой.Структура, которая образуется при репликации ДНК, называется репликативной вилкой(см. рис. ).
На рис. показан участок расплетенной ДНК. По верхней цепочке ДНК-полимераза двигается вправо к месту расплетения, непрерывно достраивая вторую (комплементарную) цепочку ДНК; направление синтеза показано стрелкой. По нижней цепочке молекулы ДНК-полимеразы двигаются налево, достраивая отдельные фрагменты комплементарной цепочки; здесь представлены два таких фрагмента (фрагменты Оказаки). Таким образом, синтез на обеих цепочках направлен от 3′-конца матричной цепочки к ее 5′-концу. Справа изображен нерасплетенный участок ДНК. На рис. показано, что исходная цепь ДНК расплелась дальше. По верхней цепочке ДНК-полимераза продолжает двигаться дальше (белая стрелка); на нижней цепочке возникает новый фрагмент Оказаки (белая стрелка). Растущий второй участок синтеза на нижней цепочке (его рост показан тоже белой стрелкой) достиг конца первого готового участка, и оба синтезированных фрагмента сшиваются ферментом лигазой.[VV93]
Постсинтетический период (G2) наступает после удвоения хромосом. Он длится 2—5 ч; за это время накапливается энергия для предстоящего митоза и синтезируются белки микротрубочек, которые впоследствии образуют веретено деления. Теперь клетка может приступать к митозу.
Клеточный цикл представляет собой однонаправленный процесс, где клетка последовательно проходит разные его периоды, без их пропуска или возврата к предыдущим стадиям.
Однако клетки могут выходить из цикла, переходить в стадию покоя, или в G0-период. В многоклеточных организмах многие клетки теряют способность к размножению, теряют способность переходить из периода покоя в новый период деления, т.е. в G-период, который начинает путь клетки к ее делению. К таким клеткам относятся нейроны, кардиомиоциты, клетки хрусталика и многие другие. Существуют также органы с редко делящимися клетками; так, например, клетки печени могут возвращаться в митотический цикл через несколько месяцев покоя. Быстро размножающиеся клетки взрослых организмов, такие как кроветворные, или базальные клетки эпидермиса и тонкой кишки, могут входить в митотический цикл каждые 12—36 ч. Самые короткие митотические циклы, около 30 мин, наблюдаются при быстром дроблении яиц низших организмов (иглокожие, земноводные).
Нарушения последовательности нуклеотидов в ДНК — генные мутации. Подробнее о мутациях мы будем говорить позже. Здесь же рассмотрим только один из их типов — генные мутации.[VV94] Изменения порядка расположения нуклеотидов в гене (а значит, изменение наследственной информации) называются генными мутациями. Такое изменение порядка может возникать в результате выпадения пары оснований, вставки новой пары или в результате замены одной пары оснований на другую.
В некоторых участках ДНК порядок расположения нуклеотидов удивительно постоянен. Это приводит к сохранению целого ряда признаков (например, похожих белков у разных организмов) в течение многих миллионов лет. Большинство нарушений в структуре ДНК исправляются сложной системой ферментов репарации. Репарация (от лат. репарацио — восстановление) — восстановление структуры ДНК, нарушенной во время репликации либо под действием физических или химических факторов (например, ультрафиолета или радиоактивных излучений). Репарация осуществляется системой ферментов, которые узнают место повреждения, вырезают поврежденные участки ДНК и др. Иногда эта система не срабатывает, и тогда порядок нуклеотидов меняется. Особенно часто новый, измененный порядок нуклеотидов возникает в ходе удвоения ДНК. Мутации возникают постоянно и в любых условиях среды у всех организмов. Мутации не всегда приводят к изменению структуры и функции образуемых белков; поэтому часто мутации не влияют на признаки и жизнеспособность организма; такие мутации называют нейтральными мутациями. Однако многие мутации, приводящие к замене всего одной или нескольких аминокислот в белке, нарушают его функционирование и оказываются вредными. Такие мутации могут быть причиной наследственных болезней. Могут возникать и мутации, которые полезны для организма. Мутации всегда случайны (не направлены на приспособление к тем условиям, в которых они возникают).
s1. Что такое клеточный цикл? 2. Что называется интерфазой? 3. Какие основные события происходят в G, S и G-периодах? 4. Каков набор хромосом клеток, находящихся в Gи G-периодах? 5.Что такое мутации и в каких случаях они возникают?6.Всегда ли мутации приводят к изменению структуры белка, почему?
Источник: studopedia.su
Введение
Для того чтобы клетка смогла полноценно разделиться, она должна увеличиться в размерах и создать достаточное количество органоидов. А для того чтобы не растерять наследственную информацию при делении пополам, она должна изготовить копии своих хромосом. И, наконец, для того чтобы распределить наследственную информацию строго поровну между двумя дочерними клеткам, она должна в правильном порядке расположить хромосомы перед их распределением по дочерним клеткам. Все эти важные задачи решаются в процессе клеточного цикла.
Клеточный цикл имеет важное значение, т.к. он демонстрирует важнейшие свойства клетки: способность к размножению, росту и дифференцировке. Обмен тоже идёт, но его не рассматривают при изучении клеточного цикла.
Определение понятия
Клеточный цикл — это период жизни клетки от рождения до образования дочерних клеток.
У животных клеток клеточный цикл, как промежуток времени между двумя делениями (митозами), длится в среднем от 10 до 24 часов.
Клеточный цикл состоит из нескольких периодов (синоним: фазы), которые закономерно сменяют друг друга. В совокупности первые фазы клеточного цикла (G1, G0, S и G2) носят название интерфазы, а последняя фаза называется митозом.
Рис. 1. Клеточный цикл.
Периоды (фазы) клеточного цикла
1. Период первого роста G1 (от английского Growth — рост), составляет 30-40% цикла, и период покоя G0
Синонимы: постмитотический (наступает после митоза) период, пресинтетический (проходит перед синтезом ДНК) период.
Клеточный цикл начинается от рождения клетки в результате митоза. После деления дочерние клетки уменьшены в размерах и органоидов в них меньше, чем в норме. Поэтому «новорожденная» маленькая клетка в первом периоде (фазе) клеточного цкла (G1) растёт и увеличивается в размерах, а также формирует недостающие органоиды. Идёт активный синтез белков, необходимых для ввсего этого. В результате клетка становится полноценной, можно сказать, «взрослой».
Чем обычно заканчивается для клетки период роста G1?
- Вступллением клетки в процесс дифференцировки. За счёт дифференцировки клетка приобретает специальные особенности для выполнения функций, необходимых всему органу и организму. Запускается дифференцировка управляющими веществами (гормонами), воздействующими на соответствующие молекулярные рецепторы клетки. Клетка, завершившая свою дифференцировку, выпадает из круговорота делений и находится в периоде покоя G0. Требуется воздействие активирующих веществ (митогенов) для того, чтобы она претерпела дедифференцировку и вновь вернулась в клеточный цикл.
- Гибелью (смертью) клетки.
- Вступлением в следующий период клеточного цикла -синтетический.
2. Синтетический период S (от английского Synthesis — синтез), составляет 30-50% цикла
Понятие синтеза в названии этого периода относится к синтезу (репликации) ДНК, а не к каким-либо другим процессам синтеза. Достигнув определенного размера в результате прохождения периода первого роста, клетка вступает в синтетический период, или фазу, S, в котором происходит синтез ДНК. За счёт репликации ДНК клетка удваивает свой генетический материал (хромосомы), т.к. в ядре образуется точная копия каждой хромосомы. Каждая хроммосома становится двойной и весь хромосомный набор становится двойным, или диплоидным. В результате клетка теперь готова поделить наследственный материал поровну между двумя дочерними клетками, не потеряв при этом ни одного гена.
3. Период второго роста G2 (от английского Growth — рост), составляет 10-20% цикла
Синонимы: премитотический (проходит перед митозом) период, постсинтетический (наступает после синтетического) период.
Период G2 является подготовительным к очередному делению клетки. Во время второго периода роста G2 клетка производит белки, требующиеся для митоза, в частности, тубулин для веретена деления; создаёт запас энергии в виде АТФ; проверяет, закончена ли репликация ДНК, и готовится к делению.
4. Период митотического деления M (от английского Mitosis — митоз), составляет 5-10% цикла
Митоз M (синоним: митотический цикл), заключается в том, что клетка правильно делится на две дочерние клетки. Благодаря механизмам комплементарного синтеза при репликации ДНК в синтетическом периоде и механизму распределения хроматид в митозе каждая дочерняя клетка получает идентичный набор хромосом, являющийся точной копией хромосомного набора материнской клетки. Короче говоря, за счёт танцевв хромосом, они расределяются пополам и поровну между двумя дочерними клетками, образовавшимися в результате деления. Подробнее о митозе…
После деления клетка оказывается в новой фазе G1, и клеточный цикл завершается.
Рис. 2. Клеточный цикл. Источник изображения: http://pisum.bionet.nsc.ru/kosterin/lectures/lecture9/lecture9.htm
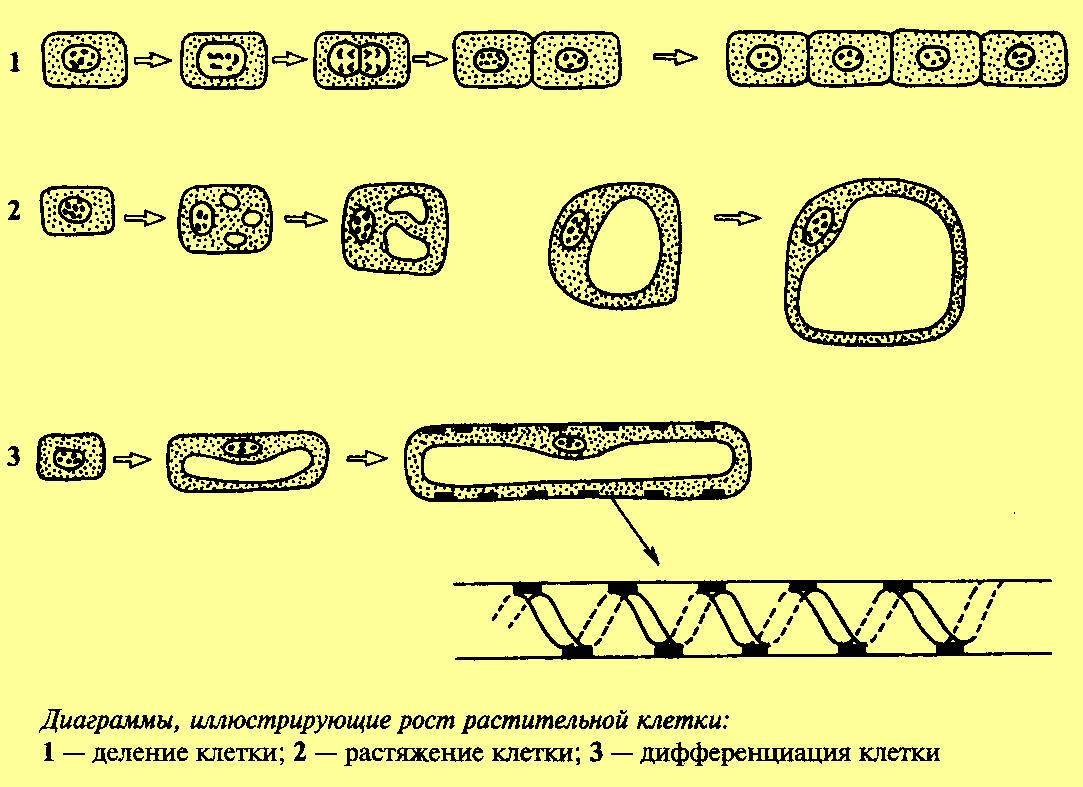
Рис. 3. Клеточный цикл растительных клеток. Источник изображения: http://fizrast.ru/razvitie/rost/osobennosti.html
Регуляция клеточного цикла
На молекулярном уровне переход от одной фазы цикла к другой регулируют два белка — циклин и циклинзависимая киназа (CDK).
Для регуляции клеточного цикла используется процесс обратимого фосфорилирования/дефосфорилирования регуляторных белков, т.е. присоединение к ним фосфатов с последующим отщеплением. Ключевым веществом, регулирующим вступление клетки в митоз (т.е. её переход от фазы G2 к фазе M), является специфическая серин/треонин-протеинкиназа, которая носит название фактор созревания — ФС, или MPF, от английского maturation promoting factor. В активной форме этот белковый фермент катализирует фосфорилирование многих белков, принимающих участие в митозе. Это, например, входящий в состав хроматина гистон H1, ламин (компонент цитоскелета, находящийся в ядерной мембране), факторы транскрипции, белки митотического веретена, а также ряд ферментов. Фосфорилирование этих белков фактором созревания MPF активирует их и запускает процесс митоза. После завершения митоза регуляторная субъединица ФС, циклин, маркируется убиквитином и подвергается распаду (протеолизу). Теперь наступает очередь протеинфосфатаз, которые дефосфорилируют белки, принимавшие участие в митозе, чем переводят их в неактивное состояние. В итоге клетка возвращается в состояние интерфазы.
ФС (MPF) — это гетеродимерный фермент, включающий в себя регуляторную субъединицу, а именно циклин, и каталитическую субъединицу, а именно циклинзависимую киназу ЦЗК (CDK от англ. cyclin dependent kinase), она же p34cdc2; 34 кДа. Активной формой этого фермента является лишь димер ЦЗК+циклин. Кроме того, активность ЦЗК регулируется путем обратимого фосфорилирования самого фермента. Циклины получили такое название потому, что их концентрация циклически изменяется в соответствии с периодами клеточного цикла, в частности, она снижается перед началом деления клетки.
В клетках позвоночных присутствует ряд различных циклинов и циклинзависимых киназ. Разнообразные сочетания двух субъединиц фермента регулируют запуск митоза, начало процесса транскрипции в G1-фазе, переход критической точки после завершения транскрипции, начало процесса репликации ДНК в S-периоде интерфазы (стартовый переход) и другие ключевые переходы клеточного цикла (на схеме не приведены).
В ооцитах лягушки вступление в митоз (G2/M-переход) регулируется путем изменения концентрации циклина. Циклин непрерывно синтезируется в интерфазе до достижения максимальной концентрации в фазе М, когда запускается весь каскад фосфорилирования белков, катализируемый ФС. К окончанию митоза циклин быстро разрушается протеиназами, также активируемыми ФС. В других клеточных системах активность ФС регулируется за счет различной степени фосфорилирования самого фермента.
Источник: kineziolog.su