Фотофосфорилирование и окислительное фосфорилирование
Темновая фаза фотосинтеза
Цикл Кальвина
Фотодыхание
C4-фотосинтез
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
В природе распространены два пути фотосинтеза растений: C3 и С4. У других организмов есть своя специфика реакций. Все, что объединяет эти разные процессы под термином «фотосинтез», – во всех них в общей сложности происходит преобразование энергии фотонов в химическую связь. Для сравнения: при хемосинтезе происходит преобразование энергии химической связи одних соединений (неорганических) в другие — органические.
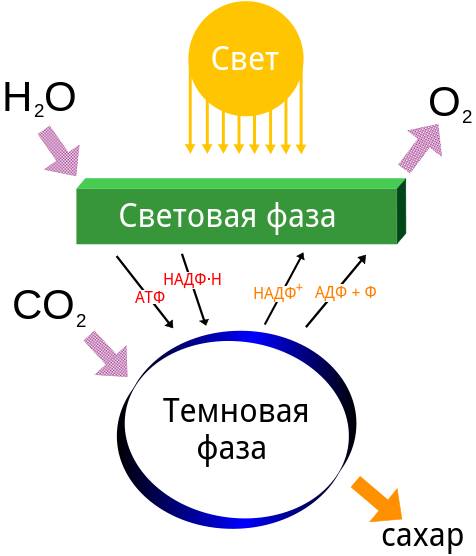
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
6CO2 + 6H2O → C6H12O6 + 6O2
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
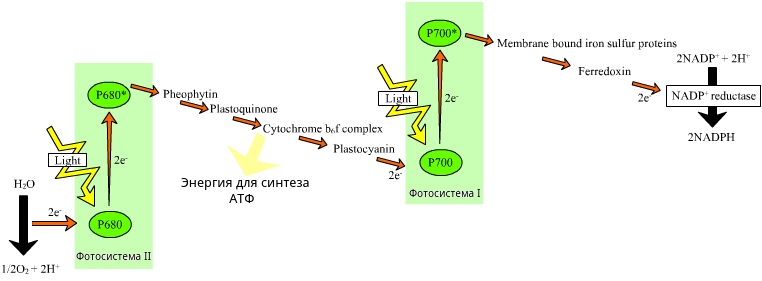
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su
Значение и роль фотосинтеза
Основной источник энергии
Слово «фотосинтез» означает буквально создание или сборку чего-то под действием света. Обычно, говоря о фотосинтезе, имеют в виду процесс, посредством которого растения на солнечном свету синтезируют органические соединения из неорганического сырья. Все формы жизни во Вселенной нуждаются в энергии для роста и поддержания жизни. Водоросли, высшие растения и некоторые типы бактерий улавливают непосредственно энергию солнечного излучения и используют ее для синтеза основных пищевых веществ. Животные не умеют использовать солнечный свет непосредственно в качестве источника энергии, они получают энергию, поедая растения или других животных, питающихся растениями. Итак, в конечном счете источником энергии для всех метаболических процессов на нашей планете, служит Солнце, а процесс фотосинтеза необходим для поддержания всех форм жизни на Земле.
Мы пользуемся ископаемым топливом — углем, природным газом, нефтью и т. д. Все эти виды топлива — не что иное, как продукты разложения наземных и морских растений или животных, и запасенная в них энергия была миллионы лет назад получена из солнечного света. Ветер и дождь тоже обязаны своим возникновением солнечной энергии, а следовательно, энергия ветряных мельниц и гидроэлектростанций в конечном счете также обусловлена солнечным излучением.
Важнейший путь химических реакций при фотосинтезе — это превращение углекислоты и воды в углероды и кислород. Суммарную реакцию можно описать уравнением СО2+Н20? [СН20]+02
Углеводы, образующиеся в этой реакции, содержат больше энергии, чем исходные вещества, т. е. СО2 и Н20. Таким образом, за счет энергии Солнца энергетические вещества (СО2 и Н20) превращаются в богатые энергией продукты — углеводы и кислород. Энергетические уровни различных реакций, описанных суммарным уравнением, можно охарактеризовать величинами окислительно-восстановительных потенциалов, измеряемых в вольтах. Значения потенциалов показывают, сколько энергии запасается или растрачивается в каждой реакции. Итак, фотосинтез можно рассматривать как процесс образования лучистой энергии Солнца в химическую энергию растительных тканей.
Содержание СО2 в атмосфере остается почти полным, несмотря на то, что углекислый газ расходуется в процессе фотосинтеза. Дело в том, что все растения и животные дышат. В процессе дыхания в митохондриях кислород, поглощаемый из атмосферы живыми тканями, используется для окисления углеводов и других компонентов тканей с образованием в конечном счете двуокиси углерода и воды и с сопутствующим выделением энергии. Высвобождающаяся энергия запасается в высокоэнергетические соединения — аденозинтрифосфат (АТФ), который и используется организмом для выполнения всех жизненных функций. Таким образом дыхание приводит к расходованию органических веществ и кислорода и увеличивает содержание СО2 на н планете. На процессы дыхания во всех живых организмах и на сжигание всех видов топлива, содержащих углерод, в совокупности расходуется в масштабах Земли в среднем около 10000 тонн 02 в секунду. При такой скорости.потребления весь кислород в атмосфере должен бы иссякнуть примерно, через 3000 лет. К счастью для нас, расход органических веществ и атомного кислорода уравновешивается созданием углеводов и кислорода в результате фотосинтеза. В идеальных условиях скорость фотосинтеза в зеленых тканях растений примерно в 30 раз превышает скорость дыхания в тех же тканях, таким образом, фотосинтез служит важным фактором, регулирующим содержание 02 на Земле.
История открытия фотосинтеза
В начале XVII в. фламандский врач Ван Гельмонт вырастил в кадке с землей дерево, которое он поливал только дождевой водой. Он заметил, что спустя пять лет, дерево выросло до больших размеров, хотя количество земли в кадке практически не уменьшилось. Ван Гельмонт, естественно, сделал вывод, что материал, из которого образовалось дерево произошел из воды, использованной для полива. В 1777 английский ботаник Стивен Хейлс опубликовал книгу, в которой сообщалось, что в качестве питательного вещества, необходимого для роста, растения используют главным образом воздух. В тот же период знаменитый английский химик Джозеф Пристли (он был одним из первооткрывателей кислорода) провел серию опытов по горению и дыханию и пришел к выводу о том, что зелёные растения способны совершать все те дыхательные процессы, которые были обнаружены в тканях животных. Пристли сжигал свечу в замкнутом объеме воздуха, и обнаруживал, что получавшийся при этом воздух уже не может поддерживать горение. Мышь, помещенная в такой сосуд, умирала. Однако веточка мяты продолжала жить в воздухе неделями. В заключение Пристли обнаружил, что в воздухе, восстановленном веточкой мяты, вновь стала гореть свеча, могла дышать мышь. Теперь мы знаем, что свеча, сгорая, потребляла кислород из замкнутого объема воздуха, но затем воздух снова насыщался кислородом благодаря фотосинтезу, происходившему в оставленной веточке мяты. Спустя несколько лет голландский врач Ингенхауз обнаружил, что растения окисляют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода. Жан Сенебье, занимавший пост министра, подтвердил данные Ингенхауза и продолжил исследование, показав, что в качестве питательного вещества растения используют двуокись углерода, растворенную в воде. В начале XIX века другой швейцарский исследователь де Соседи изучал количественные взаимосвязи между поглощенной растением углекислотой, с одной стороны, и синтезированными органическими веществами и кислородом — с другой. В результате своих опытов он пришел к выводу, что вода также потребляется растением при ассимиляции СО2. В 1817 г. два французских химика, Пельтье и Каванту, выделили из листьев зеленое вещество и назвали его хлорофиллом. Следующей важной вехой в истории изучения фотосинтеза было сделанное в 1845 г. немецким физиком Робертом Майером утверждение о том, что зеленые растения преобразуют энергию, солнечного света в химическую энергию. Представления о фотосинтезе, сложившиеся к середине прошлого века, можно выразить следующим соотношением:
Зеленое растение
СО2+ Н2 О + Свет? О2 + орг. вещества +химическая энергия
Отношение количества С02, поглощенного при фотосинтезе, к количеству выделенного 02, точно измерил французский физиолог растений Бусэнго. В 1864 г. он обнаружил, что фотосинтетическое отношение, т.е. отношение объема выделенного 02 к объему поглощенного С02, почти равно единице. В том же году немецкий ботаник Закс (открывший также у растений дыхание) продемонстрировал образование зерен крахмала при фотосинтезе. Закс помещал зеленые листья на несколько часов в темноту для того, чтобы они израсходовали накопленный в них крахмал. Затем он выносил листья на свет, но при этом освещал лишь половину каждого листа, оставляя другую половину листа в темноте. Спустя некоторое время весь лист целиком обрабатывали парами йода. В результате освещенная часть листа становилась темно-фиолетовой, что свидетельствовало об образовании комплекса крахмала с йодом, тогда как цвет другой половины листа не изменялся. Прямую связь между выделением кислорода и хлоропластами в зеленых листьях, а также соответствие спектра действия фотосинтеза спектру поглощеных хлоропластами установил в 1880 г. Энгельман. Он поместил нитевидную зеленую водоросль имеющую спирально извитые хлоропласты, на предметное стекло, освещая его узким и широким пучком белого света. Вместе с водорослью на предметное стекло наносилась суспензия клеток подвижных бактерий, чувствительных к концентрации кислорода. Предметное стекло помещали в камеру без воздуха и освещали. В этих условиях подвижные бактерии должны были перемещаться в ту часть, где концентрация 02 была выше. После прошествия некоторого времени образец рассматривали под микроскопом и подсчитывали распределение бактериопопуляции. Оказалось, что бактерии концентрировались вокруг зеленых полосок в нитевидной водоросли. В другой серии опытов Энгельман освещал водоросли лучами разного спектрального состава, установив призму между источником света и предметным столиком микроскопа. Наибольшее число бактерий в этом случае скапливалось вокруг тех участков водоросли, которые освещались синим и красным областями спектра. Находящиеся в водорослях хлорофиллы поглощали синий и красный свет. Поскольку к тому времени было уже известно, что для фотосинтеза необходимо поглощение света, Энгельман заключил, что хлорофиллы участвуют в синтезе в качестве пигментов, являющихся активными фоторецепторами. Уровень знаний о фотосинтезе в начале нашего века можно представить следующим образом.
СО2 + Н2О + Свет –О2 + Крахмал + Химическая энергия
Итак, к началу нашего века суммарная реакция фотосинтеза была уже известна. Однако биохимия находилась не на таком высоком уровне, чтобы полно раскрыть механизмы восстановления двуокиси углерода до углеводов. К сожалению, следует признать, что и теперь еще некоторые аспекты фотосинтеза изучены довольно плохо. Издавна делались попытки исследовать влияние интенсивности света, температуры, концентрации углекислоты и т… п. на общий выход фотосинтеза. И хотя в этих работах исследовались растения самых разных видов, большинство измерений было выполнено на одноклеточных зеленых водорослях и на одноклеточной жгутиковой водоросли Эвглена. Одноклеточные организмы удобнее для качественного исследования, поскольку их можно выращивать во всех лабораториях при вполне стандартных условиях. Они могут быть равномерно суспензированы, т. е. взвешены в водных буферных растворах, и нужный объем такой суспензии, или взвеси, можно брать такой дозировки, точно так же, как при работе с обычными растениями. Хлоропласты для опытов лучше всего выделять из листьев высших растений. Чаще всего используют шпинат, потому что его легко выращивать и свежие листья удобны для проведения исследований; иногда используются листья гороха и салата-латука.
Поскольку СО2 хорошо растворяется в воде, а О2 относительно нерастворим в воде, то при фотосинтезе в замкнутой системе давление газа в этой системе может изменяться. Поэтому влияние света на фотосинтетические системы часто исследуют с помощью респиратора Варбурга, позволяющего регистрировать пороговые изменения объема 02 в системе. Впервые респиратор Варбурга был использован применительно к фотосинтезу в 1920г. Для измерения потребления или выделения кислорода в ходе реакции удобнее пользоваться другим прибором — кислородным электродом. В основе этого устройства лежит использование полярографического метода. Кислородный электрод обладает достаточной чувствительностью для того, чтобы обнаружить в таких низких концентрациях как 0,01 ммоль в 1 л. Прибор состоит из катода достаточно тонкой платиновой проволоки, герметично впрессованной в пластину анода, представляющего собой кольцо из серебряной проволоки, погруженной в насыщенный раствор. Электроды отделены от смеси, в которой протекает реакция, мембраной, проницаемой для 02. Реакционная система находится в пластмассовом или стеклянном сосуде и постоянно перемешивается вращающимся стержневым магнитом. Когда к электродам приложено напряжение, платиновый электрод становится отрицательным по отношению к стандартному электроду, кислород в растворе электролитически восстанавливается. При напряжении от 0,5 до 0,8 В величина электрического тока линейно зависит от парциального давления кислорода в растворе. Обычно с кислородным электродом работают при напряжении около 0,6 В. Электрический ток измеряют, присоединив электрод к подходящей регистрирующей системе. Электрод вместе с реакционной смесью орошают потоком воды от термостата. С помощью кислородного электрода измеряют действие света и различных химических веществ на фотосинтез. Преимущество кислородного электрода перед аппаратом Варбурга состоит в том, что кислородный электрод позволяет быстро и непрерывно регистрировать изменения содержания О2 в системе. С другой стороны, в приборе Варбурга можно одновременно исследовать до 20 образцов с различными реакционными смесями, тогда как при работе с кислородным электродом образцы приходится анализировать поочередно.
Примерно до начала 1930-х годов многие исследователи в этой области полагали, что первичная реакция фотосинтеза заключается в расщеплении двуокиси углерода под действием света на углерод и кислород с последующим восстановлением углерода до углеводов с участием воды в ходе нескольких последовательных реакций. Точка зрения изменилась в 1930-х годах в результате двух важных открытий. Во-первых, были описаны разновидности бактерий, способных ассимилировать и синтезировать углеводы, не используя для этого энергию света. Затем, голландский микробиолог Ван Нил сравнил процессы фотосинтеза у бактерий и показал, что некоторые бактерии могут ассимилировать С02 на свету, не выделяя при этом кислорода. Такие бактерии способны к фотосинтезу лишь при наличии подходящего субстрата-донора водорода. Ван Нил предполагал, что фотосинтез зеленых растений и водорослей является частным случаем, когда кислород в фотосинтезе происходит из воды, а не из углекислоты.
Второе важное открытие сделал в 1937 г. Р. Хил в Кембриджском университете. С помощью дифференциального центрифугирования гомогената тканей листа он отделил фотосинтезирующие частицы (хлоропласты) от дыхательных частиц. Полученные Xиллом хлоропласты при освещении сами по себе не выделяли кислорода (возможно, из-за того, что они были повреждены при разделении). Однако они начинали выделять кислород на свету, если в суспензию вносили подходящие акцепторы электрона (окислители), например ферриоксалат калия или феррицианид калия. При выделении одной молекулы 02 фотохимически восстанавливались четыре эквивалента окислителя. Позднее было обнаружено, что многие хиноны и красители восстанавливаются хлоропластами на свету. Однако хлоропласты не могли восстановить СО2, природный акцептор электронов при фотосинтезе. Это явление, известное теперь как реакция Хилла, представляет собой индуцируемый светом перенос электронов от воды к нефизиологическим окислителям (реагентам Хилла) против градиента химического потенциала. Значение реакции Хилла состоит в том, что она продемонстрировала возможность разделения двух процессов — фотохимического выделения кислорода и восстановления углекислоты при фотосинтезе.
Разложение воды, приводящее к выделению свободного кислорода при фотосинтезе, было установлено Рубеном и Каменом, в Калифорнии в 1941 г. Они поместили фотосинтезирующие клетки в воду, обогащенную изотопом кислорода, имеющим массу 18 атомных единиц 180. Изотопный состав кислорода, выделенного клетками, соответствовал составу воды, но не С02. Кроме того, Камен и Рубен открыли радиоактивный изотоп 18О, который впоследствии успешно использовали Бассэт, Бенсон Вин, изучавшие путь превращения углекислоты при фотосинтезе. Кальвин и его сотрудник установили, что восстановление углекислоты до сахаров происходит в результате темновых ферментативных процессов, причем для восстановления одной молекулы углекислоты требуются две молекулы восстановленного АДФ и три молекулы АТФ. К тому времени роль АТФ и пиридиннуклеотидов в дыхании тканей была установлена. Возможность фотосинтетического восстановления АДФ до АТФ выделенными хлорофиллами была доказана в 1951 г. в трех разных лабораториях. В 1954 г. Арнон, Аллен продемонстрировали фотосинтез — они наблюдали ассимиляцию С02 и 02 выделенными хлоропластами шпината. В течение следующего десятилетия из хлоропластов удалось выделить белки, участвующие в переносе электронов в синтезе -ферредоксин, пластоцианин, ферроАТФ-редуктазу, цитохромы и т. д.
Таким образом, в здоровых зеленых листьях, под действием света образуются АДФ и АТФ и энергия гидросвязей используются для восстановления С02 до углеводов в присутствии ферментов, причем активность ферментов регулируется светом.
Лимитирующие факторы
Интенсивность, или скорость процесса фотосинтеза в растении зависит от ряда внутренних и внешних факторов. Из внутренних факторов наиболее важное значение имеют структура листа и содержание в нем хлорофилла, скорость накопления продуктов фотосинтеза в хлоропластах, влияние ферментов, а также наличие малых концентраций необходимых неорганических веществ. Внешние параметры — это количество и качество света, попадающего на листья, температура окружающей среды, концентрация углекислоты и кислорода в атмосфере вблизи растения.
Скорость фотосинтеза возрастает линейно, или прямо пропорционально увеличению интенсивности света. По мере дальнейшего увеличения интенсивности света нарастание фотосинтеза становится все менее и менее выраженным, и, наконец, прекращается, когда освещенность достигает определенного уровня 10000 люкс. Дальнейшее увеличение интенсивности света уже не влияет на скорость фотосинтеза. Область стабильной скорости фотосинтеза называется областью светонасыщения. Если нужно увеличить скорость фотосинтеза в этой области, следует изменять не интенсивность света, а какие-либо другие факторы. Интенсивность солнечного света, попадающего в ясный летний день на поверхность земли, во многих местах нашей планеты составляет примерно 100000 люкс. Следовательно, растениям, за исключением тех, которые растут в густых лесах и в тени, падающего солнечного света бывает достаточно для насыщения их фотосинтетической активности (энергия квантов, соответствующих крайним участкам видимого диапазона – фиолетового и красного, различается всего лишь в два раза, и все фотоны этого диапазона в принципе способны осуществить запуск фотосинтеза).
В случае низких интенсивностей света скорость фотосинтеза при 15 и 25°С одинакова. Реакции, протекающие при таких интенсивностях света, которые соответствуют области лимитирования света, подобно истинным фотохимическим реакциям, не чувствительны к температурам. Однако при более высоких интенсивностях скорость фотосинтеза при 25°С гораздо выше, чем при 15°С. Следовательно, в области светового насыщения уровень фотосинтеза зависит не только от поглощения фотонов, но и от других факторов. Большинство растений в умеренном климате хорошо функционируют в интервале температур от 10 до 35°С, наиболее благоприятные условия — это температура около 25°С.
В области лимитирования светом скорость фотосинтеза не изменяется при уменьшении концентрации СО2. Отсюда можно сделать вывод, что С02 участвует непосредственно в фотохимической реакции. В то же время при более высоких интенсивностях освещения, лежащих за пределами области лимитирования, фотосинтез существенно возрастает при увеличении концентрации СО2. У некоторых зерновых культур фотосинтез линейно возрастал при увеличении концентрации СО2 до 0,5%. (эти измерения проводили в кратковременных опытах, поскольку длительное воздействие высоких концентраций СО2 повреждает листы). Высоких значений скорость фотосинтеза достигает при содержании С02 около 0,1%. Средняя концентрация углекислоты в атмосфере составляет от 0,03%. Поэтому в обычных условиях растениям не хватает СО2 для того, чтобы с максимальной эффективностью использовать попадающий на них солнечный свет. Если помещенное в замкнутый объем растение освещать светом насыщающей интенсивности, то концентрация СО2 в объеме воздуха будет постепенно уменьшаться и достигнет постоянного уровня, известного под названием «С02 компенсационного пункта». В этой точке появление СО2 при фотосинтезе уравновешивается выделением О2 в результате дыхания (темнового и светового). У растений разных видов положения компенсационных пунктов различны.
—PAGE_BREAK—Световые и темновые реакции.
Еще в 1905 г. английский физиолог растений Ф. Ф. Блекмэн, интерпретируя форму кривой светового насыщения фотосинтеза, высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. светочувствительную реакцию и нефотохимическую, т. е. темновую, реакцию. Темновая реакция, будучи ферментативной, протекает медленнее, чем световая реакция, и поэтому при высоких интенсивностях света скорость фотосинтеза полностью определяется скоростью темновой реакции. Световая реакция либо вообще не зависит от температуры, либо зависимость эта выражена очень слабо, тогда темновая реакция, как и все ферментативные процессы, зависит от температуры в довольно значительно и степени. Следует ясно представлять себе, что реакция, называемая темновой, может протекать как в темноте, так и на свету. Световую и темновую реакции можно разделить, используя вспышки света, длящиеся краткие доли секунды. Вспышки света длительностью меньше одной миллисекунды (10-3 с) можно получить либо с помощью механического приспособления, поставив на пути пучка постоянного света вращающийся диск со щелью, либо электрически, заряжая конденсатор и разряжая его через вакуумную или газоразрядную лампу. В качестве источников света пользуются также рубиновыми лазерами с длиной волны излучения 694 нм. В 1932 г. Эмерсон и Арнольд освещали суспензию клеток вспышками света от газоразрядной лампы с длительностью около 10-3с. Они измеряли скорость выделения кислорода в зависимости от энергии вспышек, длительности темнового промежутка между вспышками и температуры суспензии клеток. При увеличении интенсивности вспышек насыщение фотосинтеза в нормальных клетках наступало, когда выделялась одна молекула 02 на 2500 молекул хлорофилла. Эмерсон и Арнольд сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию. Они также обнаружили, что при увеличении темновых интервалов между последовательными вспышками за пределы 0,06 с выход кислорода в расчете на одну вспышку уже не зависел от длительности темнового интервала, тогда как при более коротких промежутках он возрастал с увеличением длительности темнового интервала (от 0 до 0,06 с). Таким образом, темновая реакция, которая определяет уровень насыщения фотосинтеза, завершается примерно за 0,06 с. На основе этих данных было рассчитано, что среднее время, характеризующее скорость реакции, составило около 0,02 с при 25°С.
Структурная и биохимическая организация аппарата фотосинтеза
Современные представления о структурной и функциональной организации фотосинтетического аппарата включают широкий круг вопросов, связанных с характеристикой химического состава пластид, спецификой их структурной организации, физиолого-генетическими закономерностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами клетки. У наземных растений специальным органом фотосинтетической деятельности служит лист, где локализованы специализированные структуры клетки — хлоропласты, содержащие пигменты и другие компоненты, необходимые для процессов поглощения и преобразования энергии света в химический потенциал. Кроме листа функционально активные хлоропласты присутствуют в стеблях растений, черешках, остях и чешуях колоса и даже в освещаемых корнях ряда растений. Однако именно лист был сформирован в ходе длительной эволюции как специальный орган для выполнения основной функции зеленого растения – фотосинтеза, поэтому анатомия листа, расположение хлорофиллсодержащих клеток и тканей, их соотношение с другими элементами морфемной структуры листа подчинены наиболее эффективному течению процесса фотосинтеза, и они в наибольшей степени подвергаются интенсивным изменениям в условиях экологического стресса.
В связи с этим проблему структурно — функциональной организации фотосинтетического аппарата целесообразно рассмотреть в двух основных уровнях — на уровне листа как органа фотосинтеза и хлоропластов, где целиком сосредоточен весь механизм фотосинтеза.
Организация фотосинтетического аппарата на уровне листа может быть рассмотрена на основе анализа его мезострктуры. Понятие «мезоструктура» было предложено в 1975 году. По представлениям о структурной и функциональной особенностях фотоситнетического аппарата с характеристикой химического состава, структурной организации, физиолого-генетическими особенностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами специальным органом фотосинтетического процесса является лист, где локализованы специализированные образования — хлоропласты, содержащие пигменты, необходимые для процессов поглощения и преобразования света в химический потенциал. Кроме того, активные хлоропласты присутствуют в стеблях, остях и чешуях колоса и даже в освещенных частях корней некоторых растений. Однако именно лист был сформирован всем ходом эволюции как специальный орган для выполнения основной функции зеленого растения — фотосинтеза.
Мезоструктура включает систему морфофизиологических характеристик фотосинтетического аппарата листа, хлоренхимы и клезофилла. Основные показатели мезоструктуры фотосинте-
тического аппарата (по А. Т. Мокроносову) включают: площадь, число клеток, хлорофилл, белок, объем клетки, количество хлоропластов в клетке, объем хлоропласта, площадь сечения хлоропласта и его поверхность. Анализ мезоструктуры и функциональной активности фотосинтетического аппарата у многих видов растений помогают определить наиболее часто встречающиеся значения исследуемых показателей и пределы варьирования отдельных характеристик. Согласно этим данным, основные показатели мезоструктуры фотосинтетического аппарата (Мокроносов, 19В1):
I — площадь листа;
II — число клеток на 1 см2,
III — хлорофилл на 1 дм2, ключевые ферменты на 1 дм2, объем клетки, тыс. мкм2, число хлоропластов в клетке;
IV — объем хлоропластов, площадь проекции хлоропласта, мкм2, поверхность хлоропласта, мкм2.
Среднее число хлоропластов у закончившего рост листа обычно достигает 10-30, у некоторых видов оно превышает 400. Это соответствует млн хлоропластов в расчете на 1 см2площади листа. Хлоропласты сосредоточены в клетках различных тканей в количестве 15 – 80 штук на клетку. Средний объем хлоропласта — один мкм2. У большинства растений суммарный объем всех хлоропластов составляет 10-20%, у древесных растений — до 35% объема клетки. Отношение общей поверхности хлоропластов к площади листа находится в пределах 3-8. В одном хлоропласте содержится разное количество молекул хлорофилла, у тенелюбивых видов их число возрастает. Приведенные выше показатели могут значительно варьировать в зависимости от физиологического состояния и экологических условий роста растений. По данным А. Т. Мокроносова, в молодом листе активизация фотосинтеза при удалении 50-80% листа обеспечивается увеличением числа хлоропластов в клетке без изменения их индивидуальной активности, в то время как в листе, окончившем рост, усиление фотосинтеза после дефолиации происходит за счет повышения активности каждого хлоропласта без изменения их числа. Анализ мезоструктуры показал, что адаптация к условиям освещения вызывает перестройку, которая оптимизирует светопоглошающие свойства листа.
Хлоропласты имеют наиболее высокую степень организации внутренних мембранных структур по сравнению с другими органоидами клетки. По степени упорядоченности структур хлоропласты можно сравнить только с рецепторными клетками сетчатки глаза, также выполняющими функцию преобразования световой энергии. Высокая степень организации внутренней структуры хлоропласта определяется рядом моментов:
1) необходимостью пространственного разделения восстановленных и окисленных фотопродуктов, возникающих в результате первичных актов разделения заряда в реакционном центре;
2) необходимостью строгой упорядоченности компонентов реакционного центра, где сопряжёны быстропротекающие фотофизиологические и более медленные энзиматические реакции: преобразование энергии фотовозбужденной молекулы пигмента требует ее определенной ориентации по отношению к химическому акцептору энергии, что предполагает наличие определенных структур, где пигмент и акцептор жестко ориентированы друг относительно друга;
3) пространственная организация электронно-транспортной цепи требует последовательной и строго ориентированной организации переносчиков в мембране, обеспечивающей возможность быстрого и регулируемого транспорта электронов и протонов;
4) для сопряжения транспорта электронов и синтеза АТФ требуется определенным образом организованная система хлоропластов.
Липопротеидные мембраны как структурная основа энергетических процессов возникают на самых ранних этапах эволюции, предполагают, что основные липидные компоненты мембран — фосфолипиды — образовались в определенных биологических условиях. Формирование липидных комплексов обусловило возможность включения в них различных соединений, что по-видимому, явилось основой первичных каталитических функций данных структур.
Проведенные в последние годы электронномикроскопические исследования обнаружили организованные мембранные структуры у организмов, стоящих на самой низкой ступени эволюции. У некоторых бактерий мембранные фотесинтезирующие структуры клеток тесно упакованных органелл расположены по периферии клетки и связаны с цитоплазматическими мембранами; кроме того, в клетках зеленых водорослей процесс фотосинтеза связан с системой двойных замкнутых мембран — тилакоидов, локализованных в периферической части клетки. У данной группы фотосинтетических организмов впервые появляется хлорофилл, а образование специализированных органелл – хлоропластов встречается у криптофитовых водорослей. В них находятся по два хлоропласта, содержащих от одного до нескольких тилакоидов. Сходное строение фотосинтетического аппарата имеет место и у других групп водорослей: красных, бурых, и др. В процессе эволюции мембранная структура фотосинтетического процесса усложняется.
Микроскопические исследования хлоропласта, техника криоскопии позволили сформулировать пространственную модель объемной организация хлоропластов. Наиболее известна гранулярно-решетчатая модель Дж. Хеслоп-Харрисона (1964).
Таким образом, фотосинтез – это сложный процесс преобразования световой энергии в энергию химических связей органических веществ, необходимых для жизнедеятельности как самих фотосинтезирующих организмов, так и других организмов, не способных к самостоятельному синтезу органических веществ.
Изучение проблем фотосинтеза, помимо общебиологических, имеет и прикладное значение. В частности, проблемы питания, создания систем жизнеобеспечения при космических исследованиях, использования фотосинтезирующих организмов для создания различных биотехнических устройств непосредственно связаны с фотосинтезом.
Список литературы
1. Д.Халл, К.Рао «Фотосинтез». М.,1983
2. Мокроносов А.Г. «Фотосинтетическая реакция и целостность растительного организма». М.,1983
9. Красновский А.А. «Преобразование энергии света при фотосинтезе»
Для подготовки данной работы были использованы материалы с сайта www.ronl.ru/
Источник: www.ronl.ru
Этапы фотосинтеза
На самом деле, фотосинтез — очень сложный процесс. На первом его этапе идёт поглощение солнечной энергии и её передача другим молекулам, причастным к процессу. На втором этапе — разделение квантов света на заряды, в результате чего становится возможной передача электронов по фотосинтетической цепи. Благодаря этому происходит создание АТФ и НАДФН. Оба этапа имеют общее название — светозависимая стадия фотосинтеза.
Энергия, что накапливается в результате поглощения квантов света, используется в дальнейшем для образования кислорода. Но наличие самого света для этого уже не требуется. На третьем этапе происходят различные биохимические реакции, в результате которых из углекислого газа могут вырабатываться глюкоза, сахар, крахмал и т.д.
Значение фотосинтеза
Именно благодаря данному процессу Солнце является главным источником энергии на нашей планете. Многие организмы и вовсе живут лишь за счёт солнечной энергии. И они же, буквально, выдыхают её в окружающее пространство. Это позволяет другим живым организмам пользоваться ей. К примеру, всем нам известно, что мощнейшими источниками энергии для человечества являются нефть, природный газ, торф и уголь. Но мало кто знает, что вся энергия, что выделяется при сжигании этих полезных ископаемых, была запасена в результате фотосинтеза.
Но важнейшим свойством фотосинтеза, разумеется, является поглощение углекислого газа и выработка кислорода. Ведь именно благодаря этому и существует всё живое на нашей планете. Так что недооценивать важность этого процесса никак нельзя.
Источник: naturae.ru
Определение фотосинтеза
Фотосинтез — это химический процесс, посредством которого растения, некоторые бактерии и водоросли производят глюкозу и кислород из углекислого газа и воды, используя только свет в качестве источника энергии.
Этот процесс чрезвычайно важен для жизни на Земле, поскольку благодаря ему выделяется кислород, от которого зависит вся жизнь.
Зачем растениям нужна глюкоза (пища)?
Подобно людям и другим живым существам, растения также нуждаются в питании для поддержания жизнедеятельности. Значение глюкозы для растений заключается в следующем:
Глюкоза, полученная в результате фотосинтеза, используется во время дыхания для высвобождения энергии, необходимой растению для других жизненно важных процессов.
Растительные клетки также превращают часть глюкозы в крахмал, который используют по мере необходимости. По этой причине мертвые растения используются в качестве биомассы, ведь в них хранится химическая энергия.
Глюкоза также необходима, чтобы производить другие химические вещества, такие как белки, жиры и растительные сахара, необходимые для обеспечения роста и других важных процессов.
Фазы фотосинтеза
Процесс фотосинтеза разделен на две фазы: световую и темновую.
Как следует из названия, световые фазы нуждаются в солнечном свете. В светозависимых реакциях энергия солнечного света поглощается хлорофиллом и преобразуется в запасенную химическую энергию в виде молекулы электронного носителя НАДФН (никотинамидадениндинуклеотидфосфат) и молекулы энергии АТФ (аденозинтрифосфат). Световые фазы протекают в тилакоидных мембранах в пределах хлоропласта.
Темновая фаза фотосинтеза или цикл Кальвина
В темновой фазе или цикле Кальвина возбужденные электроны из световой фазы обеспечивают энергию для образования углеводов из молекул углекислого газа. Не зависящие от света фазы иногда называют циклом Кальвина из-за цикличности процесса.
Хотя темновые фазы не используют свет в качестве реагента (и, как результат, могут происходить днем или ночью), им необходимо, чтобы продукты светозависимых реакций функционировали. Независимые от света молекулы зависят от молекул энергоносителей — АТФ и НАДФН — для создания новых молекул углеводов. После передачи энергии молекулы энергоносители возвращаются к световым фазам для получения более энергичных электронов. Кроме того, несколько ферментов темновой фазы активируются с помощью света.
Схема фаз фотосинтеза
Заметка: Это означает, что темновые фазы не будут продолжаться, если растения будут лишены света слишком долго, так как они используют продукты световых фаз.
Строение листьев растений
Мы не можем полностью изучить фотосинтез, не зная больше о строении листа. Лист адаптирован для того, чтобы играть жизненно важную роль в процессе фотосинтеза.
Внешнее строение листьев
Площадь
Одной из самых главных особенностей растений является большая площадь поверхности листьев. Большинство зеленых растений имеют широкие, плоские и открытые листья, которые способны захватывать столько солнечной энергии (солнечного света), сколько необходимо для фотосинтеза.
Центральная жилка и черешок
Центральная жилка и черешок соединяются вместе и являются основанием листа. Черешок располагает лист таким образом, чтобы он получал как можно больше света.
Листовая пластинка
Простые листья имеют одну листовую пластину, а сложные — несколько. Листовая пластинка — одна из самых главных составляющих листа, которая непосредственно участвует в процессе фотосинтеза.
Жилы
Сеть жилок в листьях переносит воду от стеблей к листьям. Выделяемая глюкоза также направляется в другие части растения из листьев через жилки. Кроме того, эти части листа поддерживают и удерживают листовую пластину плоской для большего захвата солнечного света. Расположение жилок (жилкование) зависит от вида растения.
Основание листа
Основанием листа выступает самая нижняя его часть, которая сочленена со стеблем. Зачастую, у основания листа располагается парное количество прилистников.
Край листа
В зависимости от вида растения, край листа может иметь различную форму, включая: цельнокрайнюю, зубчатую, пильчатую, выемчатую, городчатую и т.п.
Верхушка листа
Как и край листа, верхушка бывает различной формы, включая: острую, округлую, туповатую, вытянутую, оттянутою и т.д.
Внутреннее строение листьев
Ниже представлена близкая схема внутреннего строения тканей листьев:
Кутикула
Кутикула выступает главным, защитным слоем на поверхности растения. Как правило, она толще на верхней части листа. Кутикула покрыта веществом, похожим на воск, благодаря которому защищает растение от воды.
Эпидермис
Эпидермис — слой клеток, который является покровной тканью листа. Его главная функция — защита внутренних тканей листа от обезвоживания, механических повреждений и инфекций. Он также регулирует процесс газообмена и транспирации.
Мезофилл
Мезофилл — это основная ткань растения. Здесь происходит процесс фотосинтеза. У большинства растений мезофилл разделен на два слоя: верхний — палисадный и нижний — губчатый.
Защитные клетки
Защитные клетки — специализированные клетки в эпидермисе листьев, которые используются для контроля газообмена. Они выполняют защитную функцию для устьица. Устьичные поры становятся большими, когда вода есть в свободном доступе, в противном случае, защитные клетки становятся вялыми.
Устьице
Фотосинтез зависит от проникновения углекислого газа (CO2) из воздуха через устьица в ткани мезофилла. Кислород (O2), полученный как побочный продукт фотосинтеза, выходит из растения через устьица. Когда устьица открытые, вода теряется в результате испарения и должна быть восполнена через поток транспирации, водой, поглощенной корнями. Растения вынуждены уравновешивать количество поглощенного СО2 из воздуха и потерю воды через устьичные поры.
Условия, необходимые для фотосинтеза
Ниже приведены условия, которые необходимы растениям для осуществления процесса фотосинтеза:
Углекислый газ. Бесцветный природный газ без запаха, обнаруженный в воздухе и имеет научное обозначение CO2. Он образуется при горении углерода и органических соединений, а также возникает в процессе дыхания.
Вода. Прозрачное жидкое химическое вещество без запаха и вкуса (в нормальных условиях).
Свет. Хотя искусственный свет также подходит для растений, естественный солнечный свет, как правило, создает лучшие условия для фотосинтеза, потому что в нем присутствует природное ультрафиолетовое излучение, которое оказывает положительное влияние на растения.
Хлорофилл. Это зеленый пигмент, найденный в листьях растений.
Питательные вещества и минералы. Химические вещества и органические соединения, которые корни растений поглощают из почвы.
Что образуется в результате фотосинтеза?
Глюкоза;
Кислород.
Заметка: Растения получают CO2 из воздуха через их листья, и воду из почвы через корни. Световая энергия исходит от Солнца. Полученный кислород выделяется в воздух из листьев. Получаемую глюкозу можно превратить в другие вещества, такие как крахмал, который используется как запас энергии.
Если факторы, способствующие фотосинтезу, отсутствуют или присутствуют в недостаточном количестве, это может негативно повлиять на растение. Например, меньшее количество света создает благоприятные условия для насекомых, которые едят листья растения, а недостаток воды замедляет.
Где происходит фотосинтез?
Фотосинтез происходит внутри растительных клеток, в мелких пластидах, называемых хлоропластами. Хлоропласты (в основном встречающиеся в слое мезофилла) содержат зеленое вещество, называемое хлорофиллом. Ниже приведены другие части клетки, которые работают с хлоропластом, чтобы осуществить фотосинтез.
Строение растительной клетки
Функции частей растительной клетки
Клеточная стенка: обеспечивает структурную и механическую поддержку, защищает клетки от патогенов, фиксирует и определяет форму клетки, контролирует скорость и направление роста, а также придает форму растениям.
Цитоплазма: обеспечивает платформу для большинства химических процессов, контролируемых ферментами.
Мембрана: действует как барьер, контролируя движение веществ в клетку и из нее.
Хлоропласты: как было описано выше, они содержат хлорофилл, зеленое вещество, которое поглощает световую энергию в процессе фотосинтеза.
Вакуоль: полость внутри клеточной цитоплазмы, которая накапливает воду.
Клеточное ядро: содержит генетическую марку (ДНК), которая контролирует деятельность клетки.
Хлорофилл поглощает световую энергию, необходимую для фотосинтеза. Важно отметить, что поглощаются не все цветовые длины волны света. Растения в основном поглощают красную и синюю волны — они не поглощают свет в зеленом диапазоне.
Углекислый газ в процессе фотосинтеза
Растения получают углекислый газ из воздуха через их листья. Углекислый газ просачивается через маленькое отверстие в нижней части листа — устьицу.
Нижняя часть листа имеет свободно расположенные клетки, чтобы углекислый газ достиг других клеток в листьях. Это также позволяет кислороду, образующемуся при фотосинтезе, легко покидать лист.
Углекислый газ присутствует в воздухе, которым мы дышим, в очень низких концентрациях и служит необходимым фактором темновой фазы фотосинтеза.
Свет в процессе фотосинтеза
Лист обычно имеет большую площадь поверхности, поэтому он может поглощать много света. Его верхняя поверхность защищена от потери воды, болезней и воздействия погоды восковым слоем (кутикулой). Верх листа находится там, где падает свет. Этот слой мезофилла называется палисадным. Он приспособлен для поглощения большого количества света, ведь в нем находится много хлоропластов.
В световых фазах, процесс фотосинтеза увеличивается с большим количеством света. Больше молекул хлорофилла ионизируется, и больше генерируется АТФ и НАДФН, если световые фотоны сосредоточены на зеленом листе. Хотя свет чрезвычайно важен в световых фазах, необходимо отметить, что чрезмерное его количество может повредить хлорофилл, и уменьшить процесс фотосинтеза.
Световые фазы не слишком сильно зависят от температуры, воды или углекислого газа, хотя все они нужны для завершения процесса фотосинтеза.
Вода в процессе фотосинтеза
Растения получают воду, необходимую для фотосинтеза через свои корни. Они имеют корневые волоски, которые разрастаются в почве. Корни характеризуются большой площадью поверхности и тонкими стенками, что позволяет воде легко проходить сквозь них.
На изображении представлены растения и их клетки с достаточным количеством воды (слева) и ее нехваткой (справа).
Заметка: Корневые клетки не содержат хлоропластов, поскольку они, как правило, находятся в темноте и не могут фотосинтезировать.
Если растение не впитывает достаточное количество воды, оно увядает. Без воды, растение будет не способно фотосинтезировать достаточно быстро, и может даже погибнуть.
Какое значение имеет вода для растений?
Обеспечивает растворенными минералами, которые поддерживают здоровье растений;
Является средой для транспортировки минеральных ресурсов;
Поддерживает устойчивость и прямостояние;
Охлаждает и насыщает влагой;
Дает возможность проводить различные химические реакции в растительных клетках.
Значение фотосинтеза в природе
Биохимический процесс фотосинтеза использует энергию солнечного света для преобразования воды и углекислого газа в кислород и глюкозу. Глюкоза используется в качестве строительных блоков в растениях для роста тканей. Таким образом, фотосинтез — это способ, благодаря которому формируются корни, стебли, листья, цветы и плоды. Без процесса фотосинтеза растения не смогут расти или размножаться.
Продуценты
Из-за фотосинтетической способности, растения известны как продуценты и служат основой почти каждой пищевой цепи на Земле. (Водоросли являются эквивалентом растений в водных экосистемах). Вся пища, которую мы едим, происходит от организмов, являющихся фотосинтетиками. Мы питаемся этими растениями напрямую или едим животных, таких как коровы или свиньи, которые потребляют растительную пищу.
Основа пищевой цепи
Внутри водных систем, растения и водоросли также составляют основу пищевой цепи. Водоросли служат пищей для беспозвоночных, которые, в свою очередь, выступают источником питания для более крупных организмов. Без фотосинтеза в водной среде жизнь была бы невозможна.
Удаление углекислого газа
Фотосинтез превращает углекислый газ в кислород. Во время фотосинтеза углекислый газ из атмосферы поступает в растение, а затем выделяется в виде кислорода. В сегодняшнем мире, где уровни двуокиси углерода растут ужасающими темпами, любой процесс, который устраняет углекислый газ из атмосферы, является экологически важным.
Круговорот питательных веществ
Растения и другие фотосинтезирующие организмы играют жизненно важную роль в круговороте питательных веществ. Азот в воздухе фиксируется в растительных тканях и становится доступным для создания белков. Микроэлементы, находящиеся в почве, также могут быть включены в растительную ткань и стать доступными для травоядных животных, дальше по пищевой цепи.
Фотосинтетическая зависимость
Фотосинтез зависит от интенсивности и качества света. На экваторе, где солнечный свет обилен весь год и вода не является ограничивающим фактором, растения имеют высокие темпы роста, и могут стать довольно большими. И наоборот, фотосинтез в более глубоких частях океана встречается реже, поскольку свет не проникает в эти слои, и в результате эта экосистема оказывается более бесплодной.

 » data-layzr=»https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза.jpg» alt=»» width=»700″ height=»350″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза.jpg 700w, https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза-300×150.jpg 300w, https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза-500×250.jpg 500w» sizes=»(max-width: 700px) 100vw, 700px» />
» data-layzr=»https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза.jpg» alt=»» width=»700″ height=»350″ data-layzr-srcset=»https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза.jpg 700w, https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза-300×150.jpg 300w, https://natworld.info/wp-content/uploads/2017/05/фазы-фотосинтеза-500×250.jpg 500w» sizes=»(max-width: 700px) 100vw, 700px» />