Стадии фотосинтеза световая и темновая
Темновая фаза фотосинтеза происходит в строме хлоропластов. Её процессы не зависят от света и могут протекать как на свету, так и в темноте, в зависимости от потребностей клетки в глюкозе. Основой темновой фазы являются циклические реакции под названием цикла фиксации углекислого газа, или цикла Кальвина. Этот процесс впервые изучил американский биохимик Мелвин Кальвин (1911 — 1997), лауреат Нобелевской премии по химии (1961). В темновой фазе из углекислого газа, водорода от НАДФ и энергии АТФ синтезируется глюкоза. Реакции фиксации СО2 катализирует рибулозобисфосфаткар-боксилаза (Rubisco) — самый распространенный фермент на Земле.
Итак, темновая фаза — совокупность циклических реакций, которые благодаря химической энергии АТФ обеспечивают образование глюкозы с использованием углекислого газа, являющегося источником углерода, и воды — источника водорода.
В чём заключается планетарная роль фотосинтеза?
Значение фотосинтеза для биосферы трудно переоценить. Именно благодаря этому процессу световая энергия Солнца превращается фото-автотрофами в химическую энергию углеводов, которые в общем дают первичное органическое вещество. С него начинаются цепи питания, по которым энергия передаётся гетеротрофным организмам. Растения служат кормом травоядным животным, которые получают за счёт этого необходимые питательные вещества. Затем травоядные животные становятся пищей для хищников, им также необходима энергия, без которой жизнь невозможна.
Только незначительная часть энергии Солнца улавливается растениями и используется для фотосинтеза. Энергия Солнца в основном идёт на испарение и поддержание температурного режима земной поверхности. Итак, только около 40 — 50% энергии Солнца проникает в биосферу, и только 1 — 2% солнечной энергии превращается в синтезированное органическое вещество.
Зелёные растения и цианобактерии влияют на газовый состав атмосферы. Весь кислород современной атмосферы является продуктом фотосинтеза. Формирование атмосферы полностью изменило состояние земной поверхности, сделало возможным появление аэробного дыхания. В дальнейшем в процессе эволюции, после образования озонового слоя, живые организмы осуществили выход на сушу. Кроме того, фотосинтез препятствует накоплению СО2, защищает планету от перегревания.
Итак, фотосинтез имеет планетарное значение, обеспечивая существование живой природы планеты Земля.
Источник: mozok.click
5. Темновая фаза фотосинтеза
Темновая фаза фотосинтез – это путь восстановления двуокиси углерода до простых сахаров. Этот процесс впервые был изучен американским ученым Кальвином, поэтому в его честь был назван циклом Кальвина.
Цикл Кальвина функционирует у высших растений, водорослей, у большинства фототрофных бактерий.
Первичным акцептором углекислого газа является рибулозо-1,5-бисфосфат. Ключевым ферментом, обеспечивающим фиксацию СО2, является фермент рибулозобисфосфаткарбоксилазаоксигеназа (Рубиско). Его активность строго коррелирует с общей интенсивностью фотосинтеза. Концентрация его в строме очень высока. На долю этого фермента приходится почти 60% растворимого белка стромы.
Цикл Кальвина, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из 4-х этапов:
-
Стадия карбоксилирования: молекулы рибулозо-5-фосфата фосфорилируются с участием АТФ и фермента фосфорибулокиназы, в результате чего образуются молекулы рибулозо-1,5-бисфосфата. К ним, в свою очередь, присоединяется углекислый газ с помощью фермента Рубиско. Полученный продукт расщепляется на 2 триозы – 2 молекулы 3-ФГК.
-
Стадия восстановления ФГК. Образовавшиеся на предыдущем этапе молекулы 3-ФГК восстанавливаются до альдегида 3-ФГА в 2 этапа. Сначала происходит фосфорилирование 3-ФГК при участии АТФ и фосфоглицераткиназы до 1,3-бисфосфоглицериновой кислоты, а затем ее восстановление до фосфоглицеринового альдегида (при участии НАДФ восстановленного).
3) Стадия регенерации первичного акцептора СО2. Происходит с участием фосфатов трех-, четырех-, пяти-, шести- и семиуглеродных сахаров. В результате процессов взаимопревращений фосфорилированных сахаров из 5 молекул трехуглеродных сахаров образуются 3 молекулы пятиуглеродных сахаров, которые затем превращаются в рибулозобисфосфат.
4) Стадия синтеза углеводных продуктов фотосинтеза. На этом этапе из фосфоглицеринового альдегида и фосфодиоксиацетона под действием фермента альдолазы синтезируется фруктозо-1,6-бисфосфат, который затем может превращаться или в крахмал, или сахарозу.
Для синтеза 1 молекулы глюкозы в цикле Кальвина необходимо 12 молекул НАДФ восстановленного и 18 молекул АТФ.
Процесс фотосинтеза осуществляется только на свету. Однако у растений часто возникает потребность в глюкозе, когда фотосинтез невозможен (в темное время суток, при прорастании семян). В этих случаях потребности в глюкозе удовлетворяются либо за счет распада сложных углеводов, либо путем новообразования глюкозы, которое называется глюконеогенезом.
Глюконеогенез – процесс образования глюкозы из неуглеводных предшественников: пирувата, аминокислот, глицерина, жирных кислот и др.). По своему химизму этот процесс напоминает обращенный гликолиз.
Глюконеогенез происходит и в организмах человека и животных. Однако ферменты, которые катализируют реакции гликолиза, имеются только в клетках печени и надпочечников. Поэтому глюконеогенез может осуществляться только в этих органах, он идет в тех случаях, когда возникает потребность в глюкозе (например, у клеток головного мозга), а запасов гликогена недостаточно и другие возможности обеспечить эти потребности отсутствуют.
Источник: StudFiles.net
Фотосинтез
Фотосинтез – это совокупность процессов синтеза органических соединений из неорганических благодаря преобразованию световой энергии в энергию химических связей. К фототрофным организмам принадлежат зеленые растения, некоторые прокариоты – цианобактерии, пурпурные и зеленые серобактерии, растительные жгутиковые.
Исследования процесса фотосинтеза начались во второй половине XVIII века. Важное открытие сделал выдающийся русский ученый К. А. Тимирязев, который обосновал учение о космической роли зеленых растений. Растения поглощают солнечные лучи и превращают световую энергию в энергию химических связей синтезированных ими органических соединений. Тем самым они обеспечивают сохранение и развитие жизни на Земле. Ученый также теоретически обосновал и экспериментально доказал роль хлорофилла в поглощении света в процессе фотосинтеза.
Хлорофиллы являются основными из фотосинтезирующих пигментов. По структуре они похожи на гем гемоглобина, но вместо железа содержат магний. Содержание железа необходимо для обеспечения синтеза молекул хлорофилла. Существует несколько хлорофиллов, которые отличаются своим химическим строением. Обязательным для всех фототрофов является хлорофилл а. Хлорофилл b встречается у зеленых растений, хлорофилл с – у диатомовых и бурых водорослей. Хлорофилл d характерен для красных водорослей.
Зеленые и пурпурные фотосинтезирующие бактерии имеют особые бактериохлорофиллы. Фотосинтез бактерий имеет много общего с фотосинтезом растений. Отличается он тем, что у бактерий донором водорода является сероводород, а у растений – вода. У зеленых и пурпурных бактерий нет фотосистемы II. Бактериальный фотосинтез не сопровождается выделением кислорода. Суммарное уравнение бактериального фотосинтеза:
6С02 + 12H2S → C6H12O6+ 12S + 6Н20.
В основе фотосинтеза лежит окислительно-восстановительный процесс. Он связан с перенесением электронов от соединений-поставщиков электронов-доноров к соединениям, которые их воспринимают – акцепторам. Световая энергия превращается в энергию синтезированных органических соединений (углеводов).
На мембранах хлоропластов есть особые структуры – реакционные центры, которые содержат хлорофилла. У зеленых растений и цианобактерий различают две фотосистемы – первую (I) и вторую (II), которые имеют разные реакционные центры и связаны между собой через систему перенесения электронов.
Две фазы фотосинтеза
Состоит процесс фотосинтеза из двух фаз: световой и темновой.
Световая фаза фотосинтеза
Происходит лишь при наличии света на внутренних мембранах митохондрий в мембранах особых структур – тилакоидов. Фотосинтезирующие пигменты улавливают кванты света (фотоны). Это приводит к «возбуждению» одного из электронов молекулы хлорофилла. С помощью молекул-переносчиков электрон перемещается на внешнюю поверхность мембраны тилакоидов, приобретая определенную потенциальную энергию.
Этот электрон в фотосистеме I может возвратиться на свой энергетический уровень и восстанавливать ее. Может также передаваться НАДФ (никотинамидадениндинуклеотидфосфат). Взаимодействуя с ионами водорода, электроны восстанавливают это соединение. Восстановленный НАДФ (НАДФ • Н) поставляет водород для восстановления атмосферного С02 до глюкозы.
Подобные процессы происходят в фотосистеме II. Возбужденные электроны могут передаваться фотосистеме I и восстанавливать ее. Восстановление фотосистемы II происходит за счет электронов, которые поставляют молекулы воды. Молекулы воды расщепляются (фотолиз воды) на протоны водорода и молекулярный кислород, который выделяется в атмосферу. Электроны используются для восстановления фотосистемы II. Уравнение фотолиза воды:
2Н20 → 4Н+ + 02 + 2е.
При возвращении электронов из внешней поверхности мембраны тилакоидов на предыдущий энергетический уровень выделяется энергия. Она запасается в виде химических связей молекул АТФ, которые синтезируются во время реакций в обеих фотосистемах. Процесс синтеза АТФ с АДФ и фосфорной кислотой называется фотофосфорилированием. Некоторая часть энергии используется для испарения воды.
Во время световой фазы фотосинтеза образуются богатые энергией соединения: АТФ и НАДФ • Н. При распаде (фотолизе) молекулы воды в атмосферу выделяется молекулярный кислород.
Темновая фаза фотосинтеза
Реакции протекают во внутренней среде хлоропластов. Могут происходить как при наличии света, так и без него. Синтезируются органические вещества (С02 восстанавливается до глюкозы) с использованием энергии, которая образовалась в световой фазе.
Процесс восстановления углекислого газа является циклическим и называется циклом Кальвина. Назван в честь американского исследователя М. Кальвина, который открыл этот циклический процесс.
Начинается цикл с реакции атмосферного углекислого газа с рибулезобифосфатом. Катализирует процесс фермент карбоксилаза. Рибулезобифосфат – это пятиуглеродный сахар, соединенный с двумя остатками фосфорной кислоты. Происходит целый ряд химических преобразований, каждое из которых катализирует свой специфический фермент. Как конечный продукт фотосинтеза образуется глюкоза, а также восстанавливается рибулезобифосфат.
Суммарное уравнение процесса фотосинтеза:
6С02 + 6Н20 → С6Н12О6 + 602
Значение фотосинтеза
Благодаря процессу фотосинтеза поглощается световая энергия Солнца и происходит преобразование ее в энергию химических связей синтезированных углеводов. По цепям питания энергия передается гетеротрофным организмам. В процессе фотосинтеза поглощается углекислый газ и выделяется кислород. Весь атмосферный кислород имеет фотосинтетическое происхождение. Ежегодно выделяется свыше 200 млрд. тонн свободного кислорода. Кислород защищает жизнь на Земле от ультрафиолетового излучения, создавая озоновый экран атмосферы.
Процесс фотосинтеза малоэффективен, так как в синтезированное органическое вещество переводится лишь 1-2 % солнечной энергии. Связано это с тем, что растения недостаточно поглощают свет, часть его поглощается атмосферой и т. п. Большая часть солнечного света отражается от поверхности Земли назад в космос.
Источник: xn—-9sbecybtxb6o.xn--p1ai
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ

Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
РиБФ + CO2 + H2O → 2ФГК
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
6CO2 + 6H2O → 2ТФ
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фосфогликолат — это фосфогликолевая кислота. От нее сразу отщепляется фосфатная группа, и она превращается в гликолевую кислоту (гликолат). Для его «утилизации» снова нужен кислород. Поэтому чем больше в атмосфере кислорода, тем больше он будет стимулировать фотодыхание и тем больше растению будет требоваться кислорода, чтобы избавиться от продуктов реакции.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) — CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
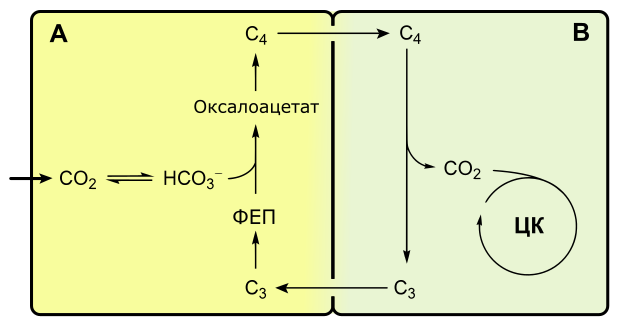
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Источник: biology.su