Вакуоль фото
 Вакуоль — это ёмкость внутри клетки, относящаяся к органоидам и используемая живым организмом для различных нужд. Обычно она имеет вид мешочка. Отделена от клетки единственной мембраной, именуемой тонопластом. Образуются вакуоли из тонопластовых пузырьков. Бывают у растений и животных, водорослей, грибов, бактерий, у вирусов и фагов их нет.
Вакуоль — это ёмкость внутри клетки, относящаяся к органоидам и используемая живым организмом для различных нужд. Обычно она имеет вид мешочка. Отделена от клетки единственной мембраной, именуемой тонопластом. Образуются вакуоли из тонопластовых пузырьков. Бывают у растений и животных, водорослей, грибов, бактерий, у вирусов и фагов их нет.
- Состав вакуоли
- Строение и функции
- Симбиоз двух организмов
Состав вакуоли
Часто основной состав органоида — это раствор необходимых веществ, то есть клеточный сок.
Несмотря на различия животных и растительных организмов, их клеточный сок представлен схожими веществами.
- Вода (например, в клетках кактуса).
- Минеральные соли: хлориды, нитраты, фосфаты (полифосфаты у фотосинтезирующих бактерий), нитраты.
- Углеводы: моносахариды, дисахариды, крахмал (в клетках клубней картофеля), гликоген (у животных).
- Жиры (например, белый жир подкожной жировой клетчатки у человека), поли-β-оксимасляная кислота (у некоторых бактерий).
- Красители: меланин (в коже человека), танин и антоцианы (у растений).
- Заживляющие вещества, заделывающие рану в случае повреждения (например, латекс в клеточной паренхиме коры гевеи).
- Газы, накапливаемые для повышения плавучести и полезного использования. У эвглены зелёной, биология которой двойственна (животное в темноте и растение на свету), накапливается и расходуется переменно углекислый газ или кислород.
Это интересно: энергетический и пластический обмен – процессы в клетке.
Строение и функции
В некоторых органах многоклеточных организмов этот органоид бурно разрастается, вытесняя прочее содержимое клетки на самый её край. Например, в горбе верблюда после прихода в оазис постепенно накапливается смесь воды и жира — вакуоли увеличиваются, горб растёт, набухает, поднимается.
Заметны различия между растительными и животными органоидами. Вакуоль у растений часто единственная в клетке, но крупная и содержащая какие-либо запасы. В животной клетке их много, они мелкие и выполняют в основном выделительные и пищеварительные функции. Рассмотрим основные типы (таблица).
| Тип вакуоли | Строение, расположение | Функции |
| Запасающая | В клетках плодов, семян, корневищ многих растений, и некоторых тканей животных, разрастаясь, занимает почти весь объём | Запас воды, питательных веществ, минералов и витаминов |
| Пищеварительная | Расположена в клетках животных, губок, микроорганизмов. Быстро меняет объём и форму | Обволакивание и переваривание органики с помощью ферментов |
| Сократительная (пульсирующая, выделительная) | В клетках животных и одноклеточных организмов. Отличается формой (у инфузорий — напоминает звёздочку) | Сбор и удаление отходов жизнедеятельности клетки, поддержание в клетке необходимого уровня осмотического давления |
| Аэросома (газовая) | Обычна для клеток растений с плавающими на воде листьями, ряски, плавучих микроводорослей наподобие спирулины, некоторых водных животных | Накачка водородом и другими газами, с целью повышения плавучести (непотопляемости) |
| Токсическая | В клетках многих растений, насекомых, рыб (фугу), ядовитых животных. Содержит алкалоиды, полифенолы и прочее (пример: соланин зелёных картофельных клубней). | Накопление ядов, используемых растениями для защиты от поедания животными и насекомыми, а животными — для «внешнего пищеварения». |
Дополнительные сведения:
- Сократительная (пульсирующая, выделительная) — её биология у одноклеточных сходна с почками и мочевым пузырём у млекопитающих.
- Пищеварительная — этот органоид быстро эволюционирует, меняя размер и содержимое. Сначала он формируется вокруг захваченного пищевого комка, обычно имеющего кислый состав. Под воздействием впрыскиваемых ферментов он увеличивается, показатель кислотности меняется на щелочной. Во время переваривания часть веществ усваивается, всасываясь в клетку, размер уменьшается. Оставшиеся отходы удаляются через сократительную вакуоль или порошицу.
- Выделяют и более узкоспециализированные органоиды, например, лизосомы — характерны для многоклеточных животных, содержат гидролитические ферменты, путём фагоцитоза, пиноцитоза утилизируют чужие бактерии, собственные отмершие органы и ткани.
Симбиоз двух организмов
Симбиоз одного живого существа с другими организмами, находящимися в его пищеварительной вакуоли, рассматривается как один из важных элементов эволюции. Особенность одноклеточных и мелких эукариот: для них обычны специализированные органоиды, по нескольку одновременно, с частой сменой, сочетанием, изменением функций.
Например, многие крупные бактерии, актинии, грибы, морские слизни практикуют пищеварительный захват микроводорослей. При этом переваривание водорослей может притормозиться со вступлением организма в симбиотическую связь с ними.
Устойчивый симбиоз гриба с водорослями внутри его органоидов привёл к появлению лишайников. Эвглена зелёная, как принято считать, имеет в качестве хлоропластов хламидомонад, эволюционировавших внутри её организма. Плавучий папоротник азолла образует заполненные слизью полости, и когда в них попадает сине-зелёная водоросль анабена (Anabaena azollae), полость закрывается, образуя вакуоль для проживания в ней этой водоросли.
Источник: obrazovanie.guru
Строение
Вакуоль растительной клетки имеет одну мембрану – тонопласт. Образование такого элемента происходит при слиянии везикул эндоплазматического ретикулума и клеток Гольджи. В молодых клетках имеется несколько маленьких вакуолей. В процессе старения клетки из мелких вакуолей получается крупная центральная вакуоль. Ее объем может достигать 90% клетки.
Функции
Основная роль – отложение и изоляция питательных веществ. Также эта структура является резервуаром для веществ, которые нужно вывести из организма, либо таковых для поддержания тургора тканей, а также растворов, способствующих росту клеток за счет растяжения. В вакуолях могут содержаться растворы для регуляции водного баланса клеток, с включением красящих элементов, создающих различные оттенки в окраске цветов, плодов, семян растений.
Наличие вакуолей – характерная деталь растений и грибов, однако и в клетках ряда видов простейших, бактерий и животных также содержатся эти органеллы. Интересно, что у ряда живых организмов – пресноводных простейших (амебы, инфузории-туфельки и т.д.) — имеются сократительные вакуоли. В них поступает пресная вода из-за разницы концентрации солей в водоеме и в клетке. По мере растяжения вакуоли в определенный момент происходит выталкивание воды наружу при сокращении ее стенок. Без таких вакуолей организмы не выживут, так как просто лопнут из-за переизбытка жидкости.
Функции вакуолей в клетках растений аналогичны таковым лизосом в клетках животных. Лизосомы представляют собой мешочки, содержащие ферменты, необходимые для переваривания макромолекул. Причем автолиз клеток растений – немаловажная функция вакуолей. Это естественный процесс, когда вредные ли ненужные вещества, а иногда и сама стареющая клетка подвергаются разрушению собственными ферментами.
Источник: beaplanet.ru
Эндоплазматическая сеть
Эндоплазматическая сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), — одномембранный органоид. Представляет собой систему мембран, формирующих «цистерны» и каналы, соединенных друг с другом и ограничивающих единое внутреннее пространство — полости ЭПС. Мембраны с одной стороны связаны с цитоплазматической мембраной, с другой — с наружной ядерной мембраной. Различают два вида ЭПС: 1) шероховатая (гранулярная), содержащая на своей поверхности рибосомы, и 2) гладкая (агранулярная), мембраны которой рибосом не несут.
Функции: 1) транспорт веществ из одной части клетки в другую, 2) разделение цитоплазмы клетки на компартменты ( «отсеки»), 3) синтез углеводов и липидов (гладкая ЭПС), 4) синтез белка (шероховатая ЭПС), 5) место образования аппарата Гольджи.
Аппарат Гольджи
Аппарат Гольджи, или комплекс Гольджи, — одномембранный органоид. Представляет собой стопки уплощенных «цистерн» с расширенными краями. С ними связана система мелких одномембранных пузырьков (пузырьки Гольджи). Каждая стопка обычно состоит из 4-х–6-ти «цистерн», является структурно-функциональной единицей аппарата Гольджи и называется диктиосомой. Число диктиосом в клетке колеблется от одной до нескольких сотен. В растительных клетках диктиосомы обособлены.
Аппарат Гольджи обычно расположен около клеточного ядра (в животных клетках часто вблизи клеточного центра).
Функции аппарата Гольджи: 1) накопление белков, липидов, углеводов, 2) модификация поступивших органических веществ, 3) «упаковка» в мембранные пузырьки белков, липидов, углеводов, 4) секреция белков, липидов, углеводов, 5) синтез углеводов и липидов, 6) место образования лизосом. Секреторная функция является важнейшей, поэтому аппарат Гольджи хорошо развит в секреторных клетках.
Лизосомы
Лизосомы — одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), содержащие набор гидролитических ферментов. Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка в мембранные пузырьки, которые после отделения от аппарата Гольджи становятся собственно лизосомами. Лизосома может содержать от 20 до 60 различных видов гидролитических ферментов. Расщепление веществ с помощью ферментов называют лизисом.
Различают: 1) первичные лизосомы, 2) вторичные лизосомы. Первичными называются лизосомы, отшнуровавшиеся от аппарата Гольджи. Первичные лизосомы являются фактором, обеспечивающим экзоцитоз ферментов из клетки.
Вторичными называются лизосомы, образовавшиеся в результате слияния первичных лизосом с эндоцитозными вакуолями. В этом случае в них происходит переваривание веществ, поступивших в клетку путем фагоцитоза или пиноцитоза, поэтому их можно назвать пищеварительными вакуолями.
Автофагия — процесс уничтожения ненужных клетке структур. Сначала подлежащая уничтожению структура окружается одинарной мембраной, затем образовавшаяся мембранная капсула сливается с первичной лизосомой, в результате также образуется вторичная лизосома (автофагическая вакуоль), в которой эта структура переваривается. Продукты переваривания усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вторичная лизосома, содержащая этот непереваренный материал, называется остаточным тельцем. Путем экзоцитоза непереваренные частицы удаляются из клетки.
Автолиз — саморазрушение клетки, наступающее вследствие высвобождения содержимого лизосом. В норме автолиз имеет место при метаморфозах (исчезновение хвоста у головастика лягушек), инволюции матки после родов, в очагах омертвления тканей.
Функции лизосом: 1) внутриклеточное переваривание органических веществ, 2) уничтожение ненужных клеточных и неклеточных структур, 3) участие в процессах реорганизации клеток.
Вакуоли
Вакуоли — одномембранные органоиды, представляют собой «емкости», заполненные водными растворами органических и неорганических веществ. В образовании вакуолей принимают участие ЭПС и аппарат Гольджи. Молодые растительные клетки содержат много мелких вакуолей, которые затем по мере роста и дифференцировки клетки сливаются друг с другом и образуют одну большую центральную вакуоль. Центральная вакуоль может занимать до 95% объема зрелой клетки, ядро и органоиды оттесняются при этом к клеточной оболочке. Мембрана, ограничивающая растительную вакуоль, называется тонопластом. Жидкость, заполняющая растительную вакуоль, называется клеточным соком. В состав клеточного сока входят водорастворимые органические и неорганические соли, моносахариды, дисахариды, аминокислоты, конечные или токсические продукты обмена веществ (гликозиды, алкалоиды), некоторые пигменты (антоцианы).
В животных клетках имеются мелкие пищеварительные и автофагические вакуоли, относящиеся к группе вторичных лизосом и содержащие гидролитические ферменты. У одноклеточных животных есть еще сократительные вакуоли, выполняющие функцию осморегуляции и выделения.
Функции вакуоли: 1) накопление и хранение воды, 2) регуляция водно-солевого обмена, 3) поддержание тургорного давления, 4) накопление водорастворимых метаболитов, запасных питательных веществ, 5) окрашивание цветов и плодов и привлечение тем самым опылителей и распространителей семян, 6) см. функции лизосом.
Эндоплазматическая сеть, аппарат Гольджи, лизосомы и вакуоли образуют единую вакуолярную сеть клетки, отдельные элементы которой могут переходить друг в друга.
Митохондрии
Форма, размеры и количество митохондрий чрезвычайно варьируют. По форме митохондрии могут быть палочковидными, округлыми, спиральными, чашевидными, разветвленными. Длина митохондрий колеблется в пределах от 1,5 до 10 мкм, диаметр — от 0,25 до 1,00 мкм. Количество митохондрий в клетке может достигать нескольких тысяч и зависит от метаболической активности клетки.
Митохондрия ограничена двумя мембранами. Наружная мембрана митохондрий (1) гладкая, внутренняя (2) образует многочисленные складки — кристы (4). Кристы увеличивают площадь поверхности внутренней мембраны, на которой размещаются мультиферментные системы (5), участвующие в процессах синтеза молекул АТФ. Внутреннее пространство митохондрий заполнено матриксом (3). В матриксе содержатся кольцевая ДНК (6), специфические иРНК, рибосомы прокариотического типа (70S-типа), ферменты цикла Кребса.
Митохондриальная ДНК не связана с белками («голая»), прикреплена к внутренней мембране митохондрии и несет информацию о строении примерно 30 белков. Для построения митохондрии требуется гораздо больше белков, поэтому информация о большинстве митохондриальных белков содержится в ядерной ДНК, и эти белки синтезируются в цитоплазме клетки. Митохондрии способны автономно размножаться путем деления надвое. Между наружной и внутренней мембранами находится протонный резервуар, где происходит накопление Н+.
Функции митохондрий: 1) синтез АТФ, 2) кислородное расщепление органических веществ.
Согласно одной из гипотез (теория симбиогенеза) митохондрии произошли от древних свободноживущих аэробных прокариотических организмов, которые, случайно проникнув в клетку-хозяина, затем образовали с ней взаимовыгодный симбиотический комплекс. В пользу этой гипотезы свидетельствуют следующие данные. Во-первых, митохондриальная ДНК имеет такие же особенности строения как и ДНК современных бактерий (замкнута в кольцо, не связана с белками). Во-вторых, митохондриальные рибосомы и рибосомы бактерий относятся к одному типу — 70S-типу. В-третьих, механизм деления митохондрий сходен с таковым бактерий. В-четвертых, синтез митохондриальных и бактериальных белков подавляется одинаковыми антибиотиками.
Пластиды
Пластиды характерны только для растительных клеток. Различают три основных типа пластид: лейкопласты — бесцветные пластиды в клетках неокрашенных частей растений, хромопласты — окрашенные пластиды обычно желтого, красного и оранжевого цветов, хлоропласты — зеленые пластиды.
Хлоропласты. В клетках высших растений хлоропласты имеют форму двояковыпуклой линзы. Длина хлоропластов колеблется в пределах от 5 до 10 мкм, диаметр — от 2 до 4 мкм. Хлоропласты ограничены двумя мембранами. Наружная мембрана (1) гладкая, внутренняя (2) имеет сложную складчатую структуру. Наименьшая складка называется тилакоидом (4). Группа тилакоидов, уложенных наподобие стопки монет, называется граной (5). В хлоропласте содержится в среднем 40–60 гран, расположенных в шахматном порядке. Граны связываются друг с другом уплощенными каналами — ламеллами (6). В мембраны тилакоидов встроены фотосинтетические пигменты и ферменты, обеспечивающие синтез АТФ. Главным фотосинтетическим пигментом является хлорофилл, который и обусловливает зеленый цвет хлоропластов.
Внутреннее пространство хлоропластов заполнено стромой (3). В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты цикла Кальвина, зерна крахмала (7). Внутри каждого тилакоида находится протонный резервуар, происходит накопление Н+. Хлоропласты, также как митохондрии, способны к автономному размножению путем деления надвое. Они содержатся в клетках зеленых частей высших растений, особенно много хлоропластов в листьях и зеленых плодах. Хлоропласты низших растений называют хроматофорами.
Функция хлоропластов: фотосинтез. Полагают, что хлоропласты произошли от древних эндосимбиотических цианобактерий (теория симбиогенеза). Основанием для такого предположения является сходство хлоропластов и современных бактерий по ряду признаков (кольцевая, «голая» ДНК, рибосомы 70S-типа, способ размножения).
Лейкопласты. Форма варьирует (шаровидные, округлые, чашевидные и др.). Лейкопласты ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя образует малочисленные тилакоиды. В строме имеются кольцевая «голая» ДНК, рибосомы 70S-типа, ферменты синтеза и гидролиза запасных питательных веществ. Пигменты отсутствуют. Особенно много лейкопластов имеют клетки подземных органов растения (корни, клубни, корневища и др.). Функция лейкопластов: синтез, накопление и хранение запасных питательных веществ. Амилопласты — лейкопласты, которые синтезируют и накапливают крахмал, элайопласты — масла, протеинопласты — белки. В одном и том же лейкопласте могут накапливаться разные вещества.
Хромопласты. Ограничены двумя мембранами. Наружная мембрана гладкая, внутренняя или также гладкая, или образует единичные тилакоиды. В строме имеются кольцевая ДНК и пигменты — каротиноиды, придающие хромопластам желтую, красную или оранжевую окраску. Форма накопления пигментов различная: в виде кристаллов, растворены в липидных каплях (8) и др. Содержатся в клетках зрелых плодов, лепестков, осенних листьев, редко — корнеплодов. Хромопласты считаются конечной стадией развития пластид.
Функция хромопластов: окрашивание цветов и плодов и тем самым привлечение опылителей и распространителей семян.
Все виды пластид могут образовываться из пропластид. Пропластиды — мелкие органоиды, содержащиеся в меристематических тканях. Поскольку пластиды имеют общее происхождение, между ними возможны взаимопревращения. Лейкопласты могут превращаться в хлоропласты (позеленение клубней картофеля на свету), хлоропласты — в хромопласты (пожелтение листьев и покраснение плодов). Превращение хромопластов в лейкопласты или хлоропласты считается невозможным.
Рибосомы
Рибосомы — немембранные органоиды, диаметр примерно 20 нм. Рибосомы состоят из двух субъединиц — большой и малой, на которые могут диссоциировать. Химический состав рибосом — белки и рРНК. Молекулы рРНК составляют 50–63% массы рибосомы и образуют ее структурный каркас. Различают два типа рибосом: 1) эукариотические (с константами седиментации целой рибосомы — 80S, малой субъединицы — 40S, большой — 60S) и 2) прокариотические (соответственно 70S, 30S, 50S).
В составе рибосом эукариотического типа 4 молекулы рРНК и около 100 молекул белка, прокариотического типа — 3 молекулы рРНК и около 55 молекул белка. Во время биосинтеза белка рибосомы могут «работать» поодиночке или объединяться в комплексы — полирибосомы (полисомы). В таких комплексах они связаны друг с другом одной молекулой иРНК. Прокариотические клетки имеют рибосомы только 70S-типа. Эукариотические клетки имеют рибосомы как 80S-типа (шероховатые мембраны ЭПС, цитоплазма), так и 70S-типа (митохондрии, хлоропласты).
Субъединицы рибосомы эукариот образуются в ядрышке. Объединение субъединиц в целую рибосому происходит в цитоплазме, как правило, во время биосинтеза белка.
Функция рибосом: сборка полипептидной цепочки (синтез белка).
Цитоскелет
Цитоскелет образован микротрубочками и микрофиламентами. Микротрубочки — цилиндрические неразветвленные структуры. Длина микротрубочек колеблется от 100 мкм до 1 мм, диаметр составляет примерно 24 нм, толщина стенки — 5 нм. Основной химический компонент — белок тубулин. Микротрубочки разрушаются под воздействием колхицина. Микрофиламенты — нити диаметром 5–7 нм, состоят из белка актина. Микротрубочки и микрофиламенты образуют в цитоплазме сложные переплетения. Функции цитоскелета: 1) определение формы клетки, 2) опора для органоидов, 3) образование веретена деления, 4) участие в движениях клетки, 5) организация тока цитоплазмы.
Клеточный центр
Клеточный центр включает в себя две центриоли и центросферу. Центриоль представляет собой цилиндр, стенка которого образована девятью группами из трех слившихся микротрубочек (9 триплетов), соединенных между собой через определенные интервалы поперечными сшивками. Центриоли объединены в пары, где они расположены под прямым углом друг к другу. Перед делением клетки центриоли расходятся к противоположным полюсам, и возле каждой из них возникает дочерняя центриоль. Они формируют веретено деления, способствующее равномерному распределению генетического материала между дочерними клетками. В клетках высших растений (голосеменные, покрытосеменные) клеточный центр центриолей не имеет. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы, они возникают в результате дупликации уже имеющихся центриолей. Функции: 1) обеспечение расхождения хромосом к полюсам клетки во время митоза или мейоза, 2) центр организации цитоскелета.
Органоиды движения
Присутствуют не во всех клетках. К органоидам движения относятся реснички (инфузории, эпителий дыхательных путей), жгутики (жгутиконосцы, сперматозоиды), ложноножки (корненожки, лейкоциты), миофибриллы (мышечные клетки) и др.
Жгутики и реснички — органоиды нитевидной формы, представляют собой аксонему, ограниченную мембраной. Аксонема — цилиндрическая структура; стенка цилиндра образована девятью парами микротрубочек, в его центре находятся две одиночные микротрубочки. В основании аксонемы находятся базальные тельца, представленные двумя взаимно перпендикулярными центриолями (каждое базальное тельце состоит из девяти триплетов микротрубочек, в его центре микротрубочек нет). Длина жгутика достигает 150 мкм, реснички в несколько раз короче.
Миофибриллы состоят из актиновых и миозиновых миофиламентов, обеспечивающих сокращение мышечных клеток.
Источник: licey.net
Общая характеристика
Тип инфузорий объединяет большое количество видов (свыше 6000) наиболее высокоорганизованных простейших.
Для них характерно присутствие ресничек, имеющихся обычно в большом числе. Реснички служат органеллами движения, они могут слипаться вместе, образуя более сложно устроенные органеллы. У некоторых сосущие инфузории реснички имеются только на ранних стадиях жизненного цикла. Для всех инфузорий характерен ядерный дуализм, т. е. двойственность. Это означает, что они имеют не менее двух ядер, различающихся как по размеру, так и по функции. Одно из ядер, значительно более крупное, называется макронуклеусом, а второе, маленькое — микронуклеусом. Некоторые виды инфузорий имеют по нескольку микро- и макронуклеусов. Микронуклеус служит половым, или генеративным, ядром, играющим основную роль в половом процессе. Макронуклеус — соматическое, или вегетативное, ядро, регулирующее все жизненные процессы, кроме полового процесса.
Бесполое размножение инфузорий происходит путем поперечного де-ления. Половой процесс у инфузорий протекает своеобразно, в виде конъюгации, которая не наблюдается у простейших других классов. Конъюгация заключается во временном сближении двух особей и взаимном обмене частями их микронуклеусов.
Инфузории — обитатели главным образом пресных водоемов, но встречаются также в солоноватой воде и в морях, некоторые виды приспособи-лись к существованию во влажной почве. Среди инфузорий много парази-тов (около 1000 видов) беспозвоночных и позвоночных животных.
Класс иразделяется на два класса:
- Ресничные инфузории (Ciliata);
- Сосущие инфузории (Suctoria).
Класс инфузории ресничные
Латинское название Ciliatas
Ресничные инфузории составляют подавляющее большинство класса.. По образу жизни и способу питания они довольно разнообразны. Одни из них ведут плавающий образ жизни, другие обитают на дне или держатся на водных растениях. Большинство ресничных инфузорий питается бак-териями и мелкими водорослями, однако есть и инфузории-хищники, поедающие других простейших, в том числе и других инфузорий. Среди ресничных инфузорий немало паразитов. Таковы инфузории, паразитирующие на коже рыб или в кишечнике различных животных и человека.
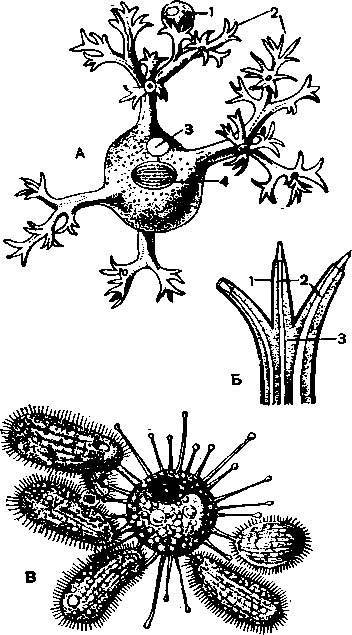
А — обыкновенная туфелька (Paramecium caudatum); 1— реснички; 2 — макронуклеу; 3— микронуклеус; 4— перистом; 5 — рот; 6 — глотка; 7 — образование пищеварительной в,акуолн; 8 — пищеварительные вакуоли; 9 — дефекация; 10 — резервуар сократительной ваку !>ли; 11, 12 — приводящие каналы сократительных вакуолей; 13 — трихоцисты; Б — брюхоре сннчная Stylonichia mytilus; 1 — адоральные мембранеллы; 2, 3, 4 и 5—группы лобных, брюшных, анальных и хвостовых цирр; 6 — ряд маргинальных цирр; 7 — спинные ще тпнкн; 8 — край перистома; 9 — иреоральные реснички; 10 — волнообразная перепонка; 11 — перистом; 12 — приводящий канал сократительной вакуоли; 13 — резервуар сократительной вакуоли; 14 — микронуклеус; 15 — макронуклеус; 16 — пищеварительная вакуоля; В — ползающая сти- лоннхия; 1 — адоральные мембранеллы; 2, 3, 4 и 5 — лобные, брюшные, анальные и XBOCI овые цпрры; 6 — маргинальные цирры; 7 — спинные щетинки; 8 — приводящие каналы; 9 — С( *кра- / тительная вакуоля.
Инфузории имеют довольно разнообразную форму тела. Однако у многих видов в связи с приспособлением их к плавающему образу жизни форма тела удлиненная, обтекаемая. Примером может служить обыкновенная туфелька (Paramecium caudatum) (рис. 2, А). Размеры также различны, некоторые виды достигают довольно крупных размеров, до 2 мм длины (Spirostomum).
Тело ипокрыто тонкой, но прочной оболочкой — пелликулой, имеющей довольно сложную структуру. Пелликула гибка и эла-стична, поэтому она не служит препятствием для некоторого изменения ЛЬормы тела. Многие инфузории могут изгибать его, протискивать между различными предметами. У крупной инфузории «трубача» (Stentor) (рп 43, А) тело вытянуто в виде граммофонной трубы, но оно может сильшЬ сжиматься и принимать шарообразную форму.
(/Р*снички — органеллы движения инфузорий. Они представляют собой очень тонкие и короткие многочисленные плазматические волоски. Ультг4атонкое строение ресничек и жгутиков, изученное с помощью* электронного микроскопа, показало их поразительное сходство.
У одних инфузорий реснички равномерно покрывают все тело. На-пример, у туфельки около 10 000—15 000 ресничек, расположенных правильными рядами. У других реснички сосредоточены в опреде-ленных местах тела.’Колебания ресничек представляют, по существу, гребные движения, состоящие из удара назад, при котором ресничка быстро движется в одной ш^кости, и возвращения в исходное положение, когда ресничка медленно двйк&тся вперед, плавно описывая полукруг. При комнатной температуре решшчки совершают около 30 взмахов в секунду. Движения ресничек происходят согласованно, в результате чего получаются правильные вол-нообразные колебания всех рядов ресничек. Туфелька движется со ско-рость^ до 2,5 мм/сек, т. е. за секунду проходит расстояние, в 10—15 раз превышающее длину ее тела.
А — строение поверхности тела Paramecium nephridiatum; 1 — попарно сидящие реснички; 2 — невроплазматическая сеть; 3 — ребрышки пелликулы; 4 — трихоцисты; 5 — отверстие трихоцисты- Б ресничный аппарат перистома стилонихии (Stylonichia mytilus) с брюшной стороны; Вто же в поперечном разрезе; 1 — предротовые реснички; 2 — ротовые реснички; 3 — предротовая волнообразная перепонка; 4 — внутренняя волнообразная перепонка; 5 — ротовая волнообразная перепонка; 6 — мембранеллы; 7 — спинные щетинки.
Кроме простых ресничек, у них имеются более крупные об-разования, обычно окружающие ротовое углубление или расположенные на других частях тела. Это так называемые мембранеллы (рис. 2, Б). Каждая мембранелла представляет собой ряд ресничек, слипшихся вм* сте в одну пластинку, часто имеющую треугольную форму (рис. 3, Б), ИЙсли слипается более длинный ряд ресничек, образуется волнообразная /пере-понка, или мембрана. Такие перепонки имеются у многих в ротовом углублении или в глотке. Строение ресничного аппарата и рас-положение различных ресничных образований служат важными система-тическими признаками.
Цитоплазма ресничных отчетливо разделяется на наружный, более светлый и плотный слой — эктоплазму и более жидкий и зернистый внутренний слой — эндоплазму (рис. 2).
Эктоплазма имеет сложное строение, образуя большое количество органелл. Она выделяет на своей поверхности уже упоминавшуюся ранее эластичную пелликулу. У туфельки пелликула имеет сложную скульптуру: она образована правильными шестигранниками, в центре которых по-мещаются реснички. По-видимому, подобное строение увеличивает прочность наружной оболочки. Эктоплазме также принадлежат реснички и мембранеллы вместе с базальными тельцами. В эктоплазме многих ин-фузорий расположены в большом количестве так называемые трихоцисты (рис. 4). Это удлиненные палочковидные тельца, сильно прелом-ляющие свет. При раздражении трихоцисты выбрасываются через особые канальцы наружу в виде тончайшей струйки жидкости, застывающей в воде тонкой упругой нитью. Трихоцисты — органеллы нападения и защиты. Хищные при помощи трихоцист парализуют добычу; «мирные» — защищаются от нападения хищников. По происхождению трихоцисты представляют собой видоизменение двигательных органелл и образуются из базальных телец.
В эктоплазме при соответствующей обработке можно обнаружить сеть тончайших волоконец, лежащих вблизи базальных телец и трихоцист (рис. 3, А). Полагают, что эти волоконца — нейрофаны — проводят раздражения и обусловливают согласованную работу ресничного аппарата. Однако во многих случаях подобные волоконца имеют опорное значение. Выше было указано., что многие из них могут изменять форму тела. Это обусловлено тем, что в эктоплазме расположены особые со-кратительные нити, или мионемы. Так, у трубача (Stentor) и некоторых других система сократительных мионем состоит из множества продольно расположенных волоконец, идущих вдоль тела и вы-стилающих околоротовое углубление (рис. 5 А). Наибольшей степени сложности достигает система мионем у Caloscolex из желудка жвачных, описанная проф. В. А. Догелем (рис. 5, Б). У сидячих инфузории сувоек имеется довольно сложна устроенный стебелек, внутри которого также проходят мионемы. При раздражении сувоек их стебелек свертывается в спираль (рис. 45).
Определенная форма тела, иногда довольно причудливая, обусловлена присутствием в эктоплазме плотных скелетных образований. Чаще всего это целая система опорных волоконец (рис. 5, В).
Таким образом, эктоплазма достигает высокой степени дифференцировки и содержит большое количество органелл: ресничный аппарат, трихоцисты, мионемы, нейрофаны и опорные скелетные волокна. В эндоплазме также расположены целые системы органелл: пищеварительных и выделительных (частично помещающихся, однако, и в эктоплазме).
А — мионемы трубача (Stentor); Б — сложная система мионем у Caloscolex; В — система опорных волоконец у Trichodina pediculus, паразитирующей на гидрах.
Пищеварительные органеллы инфузорий начинаются ртом, или цитостомо м, представляющим собой отверстие в пелликуле. У многих рот помещается на дне особого углубления — околоротовой впадины, или перистома (рис. 26, А). У многих , питающихся мелкими организмами (бактериями), перис-том окружен спирально расположенным венчиком мембранелл (разноресничные и кругоресничные). В перистоме может быть расположена волнообразная перепонка (рис. 26 и 3, Б).
Мерцательные движения ресничек и мембранелл вызывают токи воды, которыми пищевые частицы (бактерии и пр.) подносятся ко рту. У многих хищных перистома нет, и пищу они заглатывают сильно рас-тягивающимся ртом (рис. 40, В).
Рот ведет в «глотку», или цитофаринкс, представляющую собой короткий канал, иногда также выстланный ресничками. У внутреннего края глотки образуется пузырек, состоящий из капельки жидкости, выделяемой эндоплазмой, в которую попадают накапливающиеся на дне глотки пищевые частицы. Так образуется пищеварительная вакуоля (рис. 2, А).
У туфельки при изобилии пищи примерно каждую минуту образуется новая пищеварительная вакуоля. Содержащие пищу вакуоли отрываются от глотки и перемещаются в эндоплазме инфузории, совершая определенный путь. Так, у туфельки каждая пищеварительная вакуоля сначала описывает малый круг в задней половине тела, а затем большой круг, доходя до переднего конца тела . Во время передвижения в вакуоле происходит переваривание пищи и всасывание переваренной пищи в эндоплазму. В пищеварительные вакуоли эндоплазма выделяет ферменты.
А — Bursaria truncatella; Б — Dileptus unser; В — Spathidium spatula; Г — Didinium, пожирающие туфельку.
Установлено, что на разных этапах пищеварения кислотность содержимого вакуоли различна. Вначале содержимое вакуоли имеет кислую реакцию, затем щелочную.
Вакуоли, содержащие непереваренные остатки пищи, подходят к поверхности эктоплазмы. У многих инфузорий на определенном месте тела, ближе к заднему концу, в пелликуле имеется особое отверстие — цитопрокт, через которое и совершается дефекация (рис. 2, А). Процесс дефекации происходит значительно реже процесса образования пищеварительных вакуолей (через 7—10 мин), так как перед дефекацией несколько вакуолей с непереваренными остатками пищи сливаются в одну. Весь процесс пищеварения у туфельки, от образования вакуолей до дефекации, длится в зависимости от температуры от 1 до 3 ч.
Как было сказано выше, среди инфузорий немало хищников, питающихся другими (рис. 6). Например, крупная хищная Bursaria заглатывает туфелек и других, загоняя их в глотку движением мембранелл. У других хищников заглатывание происходит иначе. Рот их сильно растяжим, и они заглатывают и втягивают довольно крупных инфузорий. Некоторые хищные могут поедать инфузорий, значительно превышающих их собственные размеры. Так, сравнительно небольшие Didinium (рис. 40, Г) нападают на туфелек, убивают их особым хоботком, затем постепенно втягивают и переваривают.
Выделительные органеллы представлены у них одной, двумя или несколькими сократительными вакуолями, расположенными в определенных частях тела (рис. 2). Сократительные вакуоли часто имеют довольно сложное строение (рис. 7). Помимо самой вакуоли, периодически сжимающейся (состояние систолы) и расширяющейся (диастолы), к ней ведут расположенные в эндоплазме приводящие каналы. Благодаря этому выделяющиеся вещества поступают в сократительную вакуолю из различных частей тела инфузории. От вакуоли к пелликуле ведет выводной проток, открывающийся особым отверстием наружу (рис. 7).
П
А — сократительные вакуоли и приводящие каналы Paramecium caudatum; Б — сократи¬тельные вакуоли Campanella umbel- laria в состоянии диастолы (слева) и систолы (справа); В — схема строения сократительной вакуоли Cycloposthium; вакуоля открывается наружу постоянным каналом, окруженным осо¬быми мионемами-замыкателями (2); 2 — пелликула; Г — сократительная вакуоля Parame¬cium trichium с извитым выводным каналом (2).
ри наличии двух вакуолей (например, у туфельки) они сокращаются поочередно. При 16°С каждая вакуоля сокращается через 20—25 сек (у туфельки).
Инфузории, подобно другим простейшим, способны реагировать на разнообразные внешние раздражения. В отличие от многих жгутиковых инфузории не имеют светочувствительных органелл. Роль чувствительных органелл играют главным образом реснички и мембранел- лы. У одних реснички сохраняют при этом двигательную функцию; у других же, например у стилонихии, спинные реснички служат только осязательными органеллами.
Реакция на раздражение выражается в замедлении или ускорении, а также в изменении направления движения (туфельки), в свертывании перистома и сжатии тела (стенторы, сувойки), в сокращении стебелька
(сувойки) и т. п. Инфузории весьма чувствительны к малейшему прикосновению посторонних пре-метов. Они очень чувствительны и к изменению химического состава среды, причем различные вещества действуют на них по-разному, вызывая либо положительную, либо отрицательную реакцию. Способность различно реагировать на разные химические вещества имеет большое значение в жизни инфузорий при нахождении необходимой им пищи и наиболее благоприятных условий существования. Для дыхания оний необходимо достаточное количество растворенного в воде кислорода. Они,
как и другие простейшие, дышат всей поверхностью тела. Поэтому инфузории положительно реаги-руют на попадание в капельку воды пузырька воздуха, собираясь возле него. Инфузории реагируют положительно или отрицательно на изменение температуры среды, причем каждый вид характеризуется приспособленностью к опре-
стоит из одного или нескольких макронуклеусов, имеющих различную форму (рис. 2 и 43), и одного или нескольких микронуклеусов. В деталях строение ядерного аппарата сильно варьирует. Так, обыкновенная туфелька (Paramecium caudatum) имеет один крупный макронуклеус и один микронуклеус, помещающийся в углублении макронуклеуса. У другого вида этого же рода — Р. aurelia — имеются два микронуклеуса. У сувоек макронуклеус имеет подковообразную форму, а у трубача, кроме очень удлиненного четковидного макронуклеуса, есть несколько микронуклеусов (рис. 43). Дифференцировка ядерного аппарата на вегетативное ядро — макронуклеус и на половое, или генеративное, ядро — микронуклеус характерна для всех ресничных инфузорий.
Микронуклеус отличается от макронуклеуса не только размером, но и числом хромосом. В то время как микронуклеус обладает диплоидным набором хромосом, макронуклеус полиплоиден, т. е. набор хромосом повторен у него много раз. Так, у туфельки Paramecium caudatum макронуклеус является 80-плоидным (по другим данным, 160-плоидным), а у близкого вида Р. aurelia — 1000-плоидным. У некоторых степень плоидности может доходить до 10—15 тыс.
Таким образом, ресничные инфузории по сравнению с другими простейшими имеют очень сложное строение. Оно усложняется в двух направлениях. Мы видели, что у инфузорий имеется большое число различных органелл, часто образующих целые системы, например систему пище-варительных, выделительных органелл и т. п. С другой стороны, для инфузорий характерно умножение, или полимеризация, многих органелл. Несомненно, реснички с базальными тельцами по происхождению соответствуют жгутиковому аппарату жгутиковых. Но по сравнению с полимеризацией локомоторных органелл у многожгутиковых, у инфузорий полимеризация идет значительно дальше. Развивается сложная система органелл, состоящая из огромного количества ресничек, частью превращающихся в мембранеллы, цирры и т. п. При этом сложность организации выражается в согласованном функционировании всего двигательного аппарата. Для инфузорий характерно также умножение числа ядер. Они имеют не менее двух ядер. Однако, в отличие от многожгутиковых, этот процесс усложняется еще дифференцировкой ядер.
Размножение
Сосущие инфузории Suctoria
А — сосущая Dendrocometes para¬doxus; 1 — пойманная добыча; 2 — разветв¬ленные щупальца; 3 — сократительная вакуоля; 4— макронуклеус; Б — сосательное щупальце Dendrocometes; 1— пеллику¬ла; 2— канальцы; 3— цитоплазма; В— Sphaerophrya, сосущая нескольких ресничных.
Сосущие инфузории — небольшая специализированная группа хищников и паразитов, включающая несколько десятков видов. Все сук- тории — сидячие животные, в связи с чем во взрослом состоянии полностью лишены ресничек. Они прикрепляются различным образом к суб-страту, часто к покровам животных, например ракообразных, при помощи особой ножки, или подошвы.
У сосущих инфузорий отсутствуют обычные для ресничных органеллы: перистом, рот, глотка, пищеварительные вакуоли и т. п. Питание происходит при помощи особых сосательных щупалец, представляющих собой плазматические трубочки с каналом, ведущим внутрь эндоплазмы (рис. 46). Стоит какой-либо ресничному виду или жгутиконосцу случайно прикоснуться к сосательному щупальцу, как это простейшее прилипает к нему и оказывается в плену у сосущей инфузории. Последняя подтягивает к жертве другие щупальца. Оболочка простейшего растворяется в местах прикрепления щупалец, и цитоплазма жертвы переливается по трубочкам внутрь, в эндоплазму сосущей инфузории.
Сосательные щупальца могут быть расположены различно: пли радиально (Sphaerophrya, рис. В), или пучками на свободном конце сидящей инфузории, или на особых выростах — «руках» (Dendrocometes paradoxus; рис. А, Б). Сосущие живут и в пресных водах и в морях. Одни из них — хищники, поедающие других инфузорий; другие паразитируют внутри них. На-пример, Sphaerophrya sol паразитирует внутри туфельки.
Бесполое размножение сосущих инфузорий происходит путем наружного или внутреннего почкования. От материнской особи отделяются почки, снабженные пояском ресничек. Их называют бродяжками. Некоторое время они плавают, затем прикрепляются к субстрату, теряют реснички и превращаются в типичных сосущих инфузорий.
У сосущих имеется макронуклеус и микронуклеус. Половой процесс, так же как у ресничных, происходит путем конъюгации.
Наличие у бродяжек ресничек и двух ядер, а у взрослых сосущих полового процесса путем конъюгации сближает их с ресничными инфузориями и позволяет считать, что сосущие произошли от ресничных.
Источник: natural-museum.ru
Центральная вакуоль
Центральная вакуоль занимает более половины объема зрелых клеток, особенно в паренхиме и колленхиме. Основные функции – запас воды, накопление ионов, поддержание тургора.
Мембрана вакуоли называется тонопластом, а внутреннее содержимое — клеточным соком. Он представляет собой концентрированный раствор. Состав клеточного сока: вода, минеральные соли, сахара, танины, органические кислоты, кислород, диоксид углерода, пигменты антоцианы, продукты клеточного метаболизма и др.
Тонопласт избирательно проницаем. Через него в вакуоль поступает вода. Возникает тургорное давление, и цитоплазма прижимается к клеточной стенке. За счет такого осмотического поглощения воды клетка растягивается во время роста.
Ряд содержащихся в клеточном соке антоцианов придают разноцветную окраску цветкам и плодам. Так вакуоль выполняет функцию привлечения насекомых-опылителей и животных-распространителей плодов и семян. Совместно с каратиноидами антоцианы придают окраску осенней листве, когда разрушается хлорофилл.
Центральная вакуоль может содержать гидролитические ферменты, что позволяет ей выполнять функцию лизосом. После гибели клетки ферменты попадают в цитоплазму, и происходит автолиз.
В вакуолях накапливаются такие отходы жизнедеятельности как кристаллы оксалата кальция. Среди вторичных продуктов метаболизма — алкалоиды, которые предположительно выполняют защитную функцию наряду с танинами, препятствуя поеданию животными.
У некоторых растений в клеточном соке накапливается млечный сок, представляющий собой беловатую эмульсию. У ряда растений есть клетки, которые его экскретируют.
В центральных вакуолях также запасаются питательные вещества (сахароза, инулин), которые используются при необходимости, как и содержащиеся здесь минеральные соли.
Источник: biology.su