В каких клетках имеются хлоропласты
(греч. «хлорос» — зеленый) — двухмембранные органеллы достаточно сложного строения, содержащие хлорофилл и осуществляющие фотосинтез. Характерны только для растительных клеток ( рис. 1). У водорослей носителями хлорофилла являются хроматофоры — предшественники пластид, они же встречаются у животного — эвглены зеленой (форма разнообразная). Хлоропласты высших растений имеют форму двояковыпуклой линзы, наиболее рационально улавливающей свет. В клетке находится в среднем 10-30 (до 1000) хлоропластов. Длина пластиды составляет 5-10 мкм, толщина — 1-3, ширина — 2-4 мкм. Хлоропласты покрыты наружной гладкой мембраной, в то время как внутренняя мембрана образует в полости пластиды структуры, называемые тилакоидами (мешками). Дисковидные тилакоиды формируют граны, а трубковидные тилакоиды — тилакоиды стромы, соединяющие все граны в единую систему. В одной гране содержится от нескольких до 50 тилакоидов, а число гран в хлоропласте достигает 40-60. Пространство между тилакоидами стромы и гранами заполнено «основным веществом» — стромой.
стоящей из белков, липидов, углеводов, ферментов, АТФ. Кроме того, в строме находятся пластидная ДНК. РНК, рибосомы. Мембраны тилакоидов имеют типичное строение, но в отличие от других органелл они содержат красящие вещества — пигменты хлорофилл (зеленый) и каротиноиды (красно-оранжево-желтые). Хлорофилл — основной пигмент, связан с глобулярными белками в белково-пигментные комплексы, расположенные по наружной стороне мембраны тилакоидов гран. Каротиноиды — дополнительные пигменты, находятся в липидном слое мембраны, где они не видны, так как растворены в жирах. Но место их расположения точно соответствует белково-пигментному комплексу, поэтому пигменты в мембранах не образуют непрерывного слоя, а распределены мозаично. Строение хлоропластов тесно связано с их функцией. В них происходит фотосинтез; на мембранах тилакоидов гран проходят световые реакции, в строме — фиксация углерода (темновые реакции). Хлоропласты — полуавтономные органеллы, в которых синтезируются собственные белки, однако долго жить вне клетки они не могут, так как находятся под общим контролем ядра клетки. Размножаются они путем деления пополам или могут образовываться из пропластид или из лейкопластов. Пропластиды передаются через зиготу в виде очень маленьких телец, их диаметр составляет 0,4-1,0 мкм, они бесцветны и покрыты двойной мембраной. Пропластиды находятся в клетках конуса нарастания стебля и корня, зачатках листьев. В зеленых органах листьях, стеблях — они превращаются в хлоропласты. По окончании жизненного цикла хлорофилл разрушается (обычное изменением длины светового дня и понижением температуры), часть хлоропластов превращается в хромопласты — зеленые листья и плоды краснеют или желтеют, после чего опадают.

Рис. 1. Строение: а — хлоропласта, б — лейкопласта, в — хромопласта; 1 — внешняя мембрана, 2 — внутренняя мембрана, 3 — метрике (строма), 4 — тилакоиды стромы (ламеллы), 5 — грана, в — тилакоид граны, 7 —крахмальное зерно, 8 — каротино-иды в каплях липидов, 9 — ДНК, 10 — рибосомы, 11 — разрушающиеся мембранные структуры
Источник: Т. Л. Богданова «Пособие для поступающих в вузы»
Источник: xn--90aeobapscbe.xn--p1ai
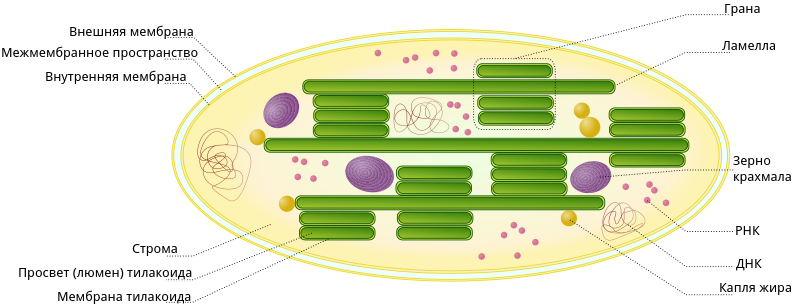
Строение хлоропласта
В строении хлоропластов выделяют внешнюю и внутреннюю мембраны, межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.
Тилакоид представляет собой ограниченное мембраной пространство в форме приплюснутого диска. Тилакоиды в хлоропластах объединяются в стопки, которые называют гранами. Граны связаны между собой удлиненными тилакоидами — ламеллами.
Полужидкое содержимое хлоропласта называется стромой. В ней находятся его ДНК и РНК, рибосомы, обеспечивающие полуавтономность органоида (см. Симбиогенез).
Также в строме находятся зерна крахмала. Они образуются при избытке углеводов, образовавшихся при фотосинтетической активности. Жировые капли обычно формируются из мембран разрушающихся тилакоидов.
Функции хлоропластов
Основная функция хлоропластов — это фотосинтез — синтез глюкозы из углекислого газа и воды за счет солнечной энергии, которая улавливается хлорофиллом. В качестве побочного продукта фотосинтеза выделяется кислород. Однако процесс этот сложный и многоступенчатый, при котором синтезируются и побочные продукты, использующиеся как в самом хлоропласте, так и в остальных частях клетки.
Основным фотосинтетическим пигментом является хлорофилл. Он существует в нескольких разных формах. Кроме хлорофилла в фотосинтезе принимают участие пигменты каротиноиды.
Пигменты локализованы в мембранах тилакоидов, здесь протекают световые реакции фотосинтеза. Кроме пигментов здесь присутствуют ферменты и переносчики электронов. Хлоропласты стараются расположиться в клетке так, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету.
Хлорофилл состоит из длинного углеводного кольца и порфириновой головки. Хвост гидрофобен и погружен в липидный слой мембран тилакоидов. Головка гидрофильна и обращена к строме. Энергия света поглощается именно головкой, что приводит к возбуждению электронов.
Электрон отделяется от молекулы хлорофилла, который после этого становится электроположительным, т. е. оказывается в окисленной форме. Электрон принимается переносчиком, которые передает его на другое вещество.
Разные виды хлорофилла отличаются между собой несколько различным спектром поглощения солнечного света. Больше всего в растениях хлорофилла А.
В строме хлоропласта происходят темновые реакции фотосинтеза. Здесь находятся ферменты цикла Кальвина и другие.
Источник: biology.su
Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных зеленых органеллах клетки. Кроме того, в растительной клетке имеются еще два вида пластид: лейкопласты – бесцветные, хромопласты – оранжевые. В лейкопластах синтезируется и отлагается в запас крахмал, в хромопластах накапливаются каратиноиды. Строение хлоропласта: Строение хлоропласта: 1 — внешняя мембрана; 2 — внутренняя мембрана; 3 — крахмальное зерно; 4 — ДНК; 5 — тилакоиды стромы (фреты); 6 — тилакоид граны; 7 — матрикс (строма); 8 – внутритилакоидное пространство (люмен). Внешняя оболочка хлоропластов отграничивает его внутреннее содержимое от цитоплазмы. Это барьер, осуществляющий контроль обмена веществ между хлоропластом и цитоплазмой. Оболочка состоит из 2-х мембран: Наружная мембрана – проницаемая для большинства органических· и неорганических молекул. Она содержит специальные транслокаторы белков, через которые поступают пептиды из цитоплазмы в хлоропласт. Внутренняя мембрана – избирательно проницаема и осуществляет· контроль над транспортом белков, липидов, органических кислот и углеводов между хлоропластом и цитоплазмой. Участвует в формировании внутренней мембранной системы хлоропластов. Строма – гидрофильный, слабоструктурированный матрикс хлоропластов, содержащий водорастворимые органические соединения, а также неорганические ионы. В строме осуществляются реакции фотосинтетической ассимиляции углерода. В строме находятся: кольцевая ДНК, рибосомы, ферменты матричного синтеза. Внутренняя мембранная система хлоропластов – здесь протекают световые реакции фотосинтеза. Мембраны образуют тилакоиды, которые либо тесно соприкасаются друг с другом и уложены в стопки, или граны (тилакоиды гран – 6), либо пронизывают строму, соединяя граны между собой (тилакоиды стромы (фреты)). Собственно образующие их мембраны называют мембранами (ламеллами) гран и мембранами (ламеллами) стромы. Пространство внутри тилакоидов называется внутритилакоидным пространством, или люменом. Значение сложной организации внутренних мембран хлоропластов: Благодаря значительному мембранному пространству достигается· увеличение числа функциональных единиц, способных осуществлять световые реакции фотосинтеза. Единство внутренней системы хлоропластов позволяет отдельным· компонентам мембраны мигрировать латерально и вступать между собой в структурный и функциональный контакт. Это необходимо для переноса энергии квантов света в реакционные центры, а также для транспорта электронов по электрон-транспортной цепи в ходе световых реакций фотосинтеза. Разделение мембраной всего внутреннего пространства хлоропластов на· два компонента – стромальное и люмен – позволяет создавать электрохимические градиенты ионов между ними. Создание электрохимического градиента Н+ на внутренних мембранах хлоропластов – важный этап в трансформации энергии квантов света в энергию макроэргических связей АТФ. Образование гранальной структуры внутри хлоропластов значительно· повышает общую эффективность фотосинтеза и создает дополнительные возможности для регуляции световых реакций. Сегрегация (разделение) в стромальных или гранальных тилакоидах компонентов мембран с различными функциями позволяет добиться определенной независимости их функционирования. Это итог длительного эволюционного процесса – впервые появилась у зеленых водорослей. Основные этапы образования хлоропластов. Предшественники хлоропластов – пропластиды. Пропластиды образуются из инициальных частиц (зачатков), содержащихся в меристиматических клетках. Формирование хлоропласта может осуществляться двумя путями: I путь – непосредственное преобразование пропластид в хлоропласты. Реализуется при росте растений в условиях нормального соотношения дня и ночи. Пропластиды меристиматических клеток листа превращаются в хлоропласты параллельно с ростом и дифференцировкой клеток листа. Биогенез хлоропластов сопровождается формированием тилакоидных мембран хлоропластов при участии внутренней мембраны оболочки пропластиды. II путь – образование хлоропластов из этиопластов. Этиопласты – органеллы клеток растения, растущего в отсутствие света. Они образуются из пропластид и имеют некоторвые особенности внутреннего строения: содержат проламеллярное тело, сформированное в результате скопления ограниченных мембраной пузырьков и разветвленных трубчатых структур. Мембраны проламеллярного тела содержат небольшие количества каротиноидов и предшественника хлорофилла – протохлорифиллида. Формирование тилакоидных мембран хлоропластов в этиопластах происходит при участии мембран проламелярного тела в ответ на освещение. Выделяют три этапа фотоморфогенеза хлоропластов из этиопластов: 1 этап: Из трубчатых элементов проламелярных тел образуются крупные пузырьки, располагающиеся по радиусу. Этот процесс сопровождается образованием хлорофилла из имеющегося в этиопластах протохлорофиллида. Источник: studopedia.info фотосинтез протекает в специализированных органеллах клеток — хлоропластах. Хлоропласты высших растений имеют форму двояковыпуклой линзы (диска), которая наиболее удобна для поглощения солнечных лучей. Их размеры, количество, расположение полностью отвечают назначению: как можно эффективнее поглощать солнечную энергию, как можно полнее усваивать углерод. Установлено, что количество хлоропластов в клетке измеряется десятками. Это обеспечивает высокое содержание этих органелл на единицу поверхности листа. Так, на 1 мм2 листа фасоли приходится 283 тыс. хлоропластов, у подсолнечника — 465 тыс. Диаметр хлоропластов в среднем 0,5-2 мкм. Строение хлоропласта весьма сложное. Подобно ядру и митохондриям хлоропласт окружен оболочкой, состоящей из двух липопротеидных мембран. Внутреннюю среду представляет относительно однородная субстанция — матрикс или строма, которую пронизывают мембраны — ламеллы (рис.). Ламеллы, соединенные друг с другом, образуют пузырьки — тилакоиды. Плотно прилегая друг к другу, тилакоиды образуют граны, которые различают даже под световым микроскопом. В свою очередь, граны в одном или нескольких местах объединены друг с другом с помощью межгранных тяжей — тилакоидов стромы. Свойства хлоропластов: способны измененять ориентацию и перемещаться. Например, под влиянием яркого света хлоропласты поворачиваются узкой стороной диска к падающим лучам и перемещаются на боковые стенки клеток. Хлоропласты передвигаются в направлении более высокой концентрации СО2 в клетке. Днем они обычно выстраиваются вдоль стенок, ночью опускаются на дно клетки. Химический состав хлоропластов: воды — 75 %; 75-80 % общего количества сухих веществ составляют орг. соединения, 20-25 % -минеральные. Структурной основой хлоропластов являются белки (50-55 % сухой массы), половина из них составляют водорастворимые белки. Такое высокое содержание белков объясняется их многообразными функциями в составе хлоропластов (структурные белки мембран, белки-ферменты, транспортные белки, сократительные белки, реценторные). Важнейшей составной частью хлоропластов являются липиды, (30-40% сух. м.). Липиды хлоропластов представлены тремя группами соединений.
В хлоропластах листьев присутствует значительное количество РНК и ДНК. НК составляют примерно 1 % сухой массы хлоропластов (РНК — 0.75 %, ДНК — 0,01-0,02 %). Геном хлоропластов представлен кольцевой молекулой ДНК длиной 40 мкм с молекулярной массой 108, кодирующей 100-150 белков средних размеров. Рибосомы хлоропластов составляют от 20 до 50 % общей популяции рибосом клетки. Т.о., хлоропласты имеют собственную белоксинтезирующую систему. Однако для нормального функционирования хлоропластов необходимо взаимодеЯствие ядерного и хлоропластного геномов. Ключевой фермент фотосинтеза РДФ-карбоксилаза синтезируется под двойным контролем-ДНК ядра и хлоропласта. Углеводы не являются конституционными веществами хлоропласта. Представлены фосфорными эфирами сахаров и продуктами фотосинтеза. Поэтому содержание углеводов в хлоропластах значительно колеблется (от 5 до 50 %). В активно функционирующих хлоропластах углеводы обычно не накапливаются, происходит их быстрый отток. При уменьшении потребности в продуктах фотосинтеза в хлоропластах образуются крупные крахмальные зерна. В этом случае содержание крахмала может возрасти до 50 % сухой массы и активность хлоропластов снизится. Минеральные вещества. Сами хлоропласты составляют 25-30 % массы листа, но в них сосредоточено до 80 % Fe, 70-72 — Mg и Zn, 50 — Cu, 60 % Ca, содержащихся в тканях листа. Это объясняется высокой и разнообразной ферментативной активностью хлоропластов (входят с состав простетических групп и кофакторов). Mg входит в состав хлорофилла. Ca стабилизирует мембранные структуры хлоропластов. Возникновение и развитие хлоропластов. Хлоропласты образуются в меристематических клетках из инициальных частиц или зачаточных пластид (рис.). Инициальная частица состоит из амебоидной стремы, окруженной двухмембранной оболочкой. По мере роста клетки инициалььные частицы увеличиваются в размере и приобретают форму двояковыпуклой линзы, в стреме появляются небольшие крахмальные зерна. Одновременно внутренняя мембрана начинает разрастаться, образуя складки (впячивания), от которых отшнуровываются пузырьки и трубочки. Такие образования называют пропластидами. Для дальнейшего их развития необходим свет. В темноте же формируются этиопласты, в которых образуется мембранная решетчатая структура — проламеллярное тело. На свету внутренние мембраны пропластид и этиопластов образуют гранильную систему. Одновременно с этим также на свету в граны встраиваются вновь образованные молекулы хлорофилла и других пигментов. Таким образом, структуры, которые подготавливаются к функционированию на свету, появляются и развиваются только при его наличии. Наряду с хлоропластами имеется ряд других пластид, которые образуются либо непосредственно из пропластид, либо одна из другой путем взаимных превращений (рис.). К ним относятся накапливающие крахмал амилопласты (лейкопласты) и хромопласты, содержащие каротиноиды. В цветках и плодах хромопласты возникают на ранних стадиях развития пропластид. Хромопласты осенней листвы представляют собой продукты деградации хлоропластов, в которых в качестве структур — носителей каротнноидов выступают пластоглобулы. Пигменты хлоропласта, участвующие в улавливании световой энергии, а также ферменты, необходимые для световой фазы фотосинтеза, вмонтированы в мембраны тилакоидов. Ферменты, которые катализируют многочисленные реакции восстановительного цикла углеводов (темповой фазы фотосинтеза), а также разнообразные биосинтезы, в том числе биосинтезы белков, липидов, крахмала, присутствуют главным образом в строме, часть из них является периферическими белками ламелл. Строение зрелых хлоропластов одинаково у всех высших растений, так же как в клетках разных органов одного растения (листьях, зеленеющих корнях, коре, плодах). В зависимости от функциональной нагрузки клеток, физиологического состояния хлоропластов, их возраста различают степень их внутренней структурированности: размеры, количество гран, связь между ними. Так, в замыкающих клетках устьиц основная функция хлоропластов — фоторегуляция устьичных движений. Хлоропласты не имеют строгой гранальной структуры, содержат крупные крахмальные зерна, набухшие тилакоиды, липофильные глобулы. Все это свидетельствует об их низкой энергетической нагрузке (эту функцию выполняют митохондрии). Другая картина наблюдается при изучении хлоропластов зеленых плодов томата. Наличие хорошо развитой гранулярной системы свидетельствует о высокой фукциональной нагрузке этих органелл и, вероятно, существенном вкладе фотосинтеза при формировании плодов. Возрастные изменения: Молодые характеризуются ламеллярнои структурой, в таком состоянии хлоропласты способны размножаться путем деления. В зрелых хорошо выражена система гран. В стареющих происходит разрыв тилакоидов стромы, связь между гранами уменьшается, в дальнейшем наблюдаются распад хлорофилла и деструкция гран. В осенней листве деградация хлоропластов приводит к образованию хромопластов. Структура хлоропластов лабильна и динамична, в ней отражаются все условия жизни растения. Большое влияние оказывает режим минерального питания растений. При недостатке N хлоропласты становятся в 1.5-2 раза мельче, дефицит P и S нарушает нормальную структуру ламелл и гран, одновременная нехватка N и Ca приводит к переполнению хлоропластов крахмалом из-за нарушения нормального оттока ассимилятов. При недостатке Ca нарушается структура наружной мембраны хлоропласта. Для поддержания структуры хлоропласта также необходим свет, в темноте идет постепенное разрушение тилакоидов гран и стремы. Источник: StudFiles.net |