Что называется цитокинезом
Цитокинез – это процесс деления эукариотической клетки. Цитокинез был одним из первых событий клеточного цикла, наблюдаемых с помощью простых клеточных биологических методов, однако молекулярная характеристика цитокинеза замедлилась благодаря особой устойчивости к биохимическим подходам in vitro. В то время как результат цитокинеза одинаков во всех делящихся клетках, механизм деления варьируется в разных крупных эукариотических царствах. Например, дрожжи и животные используют сократительное кольцо, которое проникает в середину клетки, чтобы разделить ее, в то время как клетки строят новую клеточную стенку наружу, к коре. Как и следовало ожидать, существуют некоторые сходства в молекулах, участвующих в цитокинезе одноклеточных и клетках животных, но на первый взгляд цитокинез в растительной и животной клетке кажется совершенно разным. Однако в последние годы стало ясно, что базовые процессы совершенно одинаковы у клеток растений, простейших и животных.
Один из видов цитокинеза – это митоз, который подразделяется на несколько стадий: профаза, метафаза, анафаза и телофаза. Ниже приведены особенности каждой фазы цитокинеза.
Профаза
Профаза знаменуется бурными биохимическими изменениями, по результатам которых клетка приходит в состояние, после которого начинается непосредственно деление. Во время профазы хромосомы собираются в центре клетки, а затем удваиваются, предоставляя генетический материал для обеих новообразованных дочерних клеток. Обычно их не видно в микроскоп, однако в тот момент они становятся хорошо различимыми в оптический микроскоп. Также в это время пропадает ядрышко. К середине профазы полностью останавливается транскрипционная активность. Особенности строения клеток таковы, что на ранних стадиях цитокинеза у клеток с крупными хромосомами он замедляется и может растягиваться на несколько часов, в то время как в клетках существ с мелкими хромосомами (например, млекопитающих) она длится около 15 минут. По прошествии этого времени начинается деление тела эукариотической клетки.
Метафаза
Метафаза цитокинеза – это стадия деления клетки, в которой хромосомы выходят в экваториальную плоскость клетки. Микротрубочки на этой фазе обновляются особенно активно. Хромосомы в клетке располагаются так, чтобы ранее упомянутым микротрубочкам было удобно к ним прикрепиться. Сестринские хроматиды разделяются, но не расходятся, останавливаемые центромерами. Из-за особенностей строения клетки метафаза может завершиться только после того, как клетке пошлет сигнал комплекс стимуляции анафазы.
ким образом, если разрушить веретено деления, хромосомы не смогут перейти к анафазе до тех пор, пока вредоносное воздействие не будет полностью устранено. Такой метод исследования часто употребляется генетиками для того, чтобы создать клетки, часами находящиеся в метафазе, которые потом используют для исследований. Молекулярные механизмы этого действия пока остаются загадкой, но на данный момент над раскрытием их тайн успешно работают ученые.
Анафаза
За метафазой следует анафаза. Для цитокинеза это одновременно самая ключевая и самая короткая стадия, во время которой сестринские хроматиды расходятся к краям клетки, образуя дочерние хромосомы. Несмотря на то, что анафаза является самой короткой стадией, она подразделяется на множество фаз. Эти стадии контролирует упомянутый ранее комплекс стимуляции анафазы. Во время анафазы хромосомы расходятся в две новые клетки. Хроматиды каждой хромосомы разгоняются на противоположные стороны клеток, образуя две новые дочерние клетки. Каждая сторона клетки начинает обладать полным набором хромосом. Анафаза важна, потому что именно она помогает ДНК разделить на две части, идя в обе стороны клетки. Она гарантирует, что следующий ген сможет выполнить свою функцию. Если этого не произойдет, тогда не будет двух разных ДНК для следующего процесса
Телофаза
Телофаза – завершающая часть деления клетки. Ее название происходит от латинского слова telos, что означает конец.
этом этапе сестринские хроматиды достигают противоположных полюсов. Маленькие ядерные везикулы в клетке начинают перестраиваться вокруг группы хромосом на каждом конце. По мере того, как ядерная оболочка реформируется путем связывания с хромосомами, в одной клетке создаются два ядра. Телофаза также отмечается растворением микротрубочек кинетохора и продолжением удлинения полярных микротрубочек. По мере преобразования ядерных оболочек хромосомы начинают разлагаться и становиться более рассеянными. После того, как все процессы завершены, две новые клетки начинают функционировать, как ни в чем не бывало.
Как мы увидели, цитокинез является сложным, но в то же время понятным и завораживающим процессом. Ученые до сих пор изучают особенности строения клетки.
Источник: fb.ru
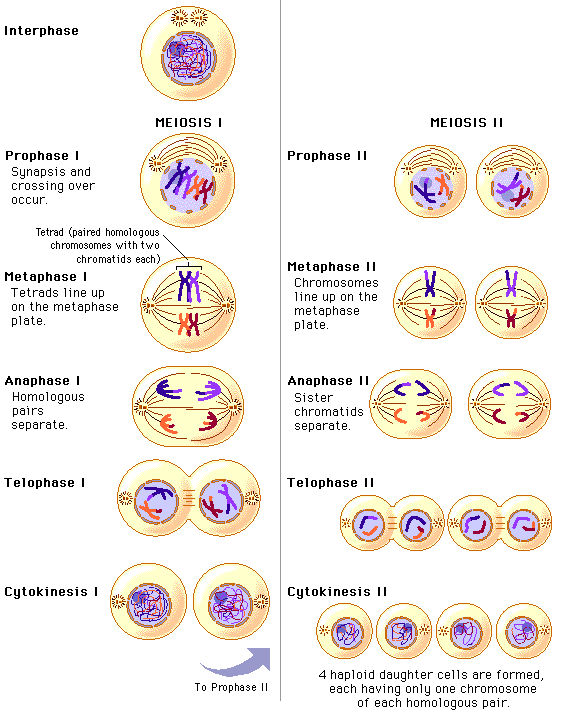
Фазы мейоза
У организмов, размножающихся половым путем, имеется две категории клеток: диплоидные и гаплоидные. К первым относятся соматические и предшественницы половых клеток, ко вторым — зрелые половые (гаметы). Уменьшение количества хромосом в два раза достигается благодаря мейозу. Он включает в себя два последовательных деления. После слияния гамет возникает новый одноклеточный диплоидный организм (зигота), который несет не просто сумму признаков своих родителей, а является индивидуумом с присущими только ему свойствами.
При дальнейшем митотическом делении зиготы образуются диплоидные же клетки, содержащие по два экземпляра каждой хромосомы, которые называются гомологичными. Гомологичные хромосомы, имеющие одинаковую длину и одинаковое расположение центромер, содержат одинаковое количество генов, а эти гены имеют одну и ту же линейную последовательность. Каждая из пары гомологичных хромосом диплоидного организма происходит либо из ядра спермия, либо из ядра яйцеклетки.
При образовании гамет в зрелом организме в результате мейоза в каждую дочернюю клетку от всех пар гомологичных хромосом попадает лишь по одной из них. Это становится возможным потому, что при мейозе происходит лишь одна репликация ДНК, за которой следуют два последовательных деления ядер (мейоз I и II) без повторного синтеза ДНК. В результате из одной диплоидной образуются четыре гаплоидные клетки.
Напомним, что перед началом мейоза в интерфазе клетка прошла обычные фазы Gx, S и G2, так что стала тетраплоидной. Иначе говоря, произошла репликация ДНК и бел-ков-гистонов хромосом, а сестринские хроматиды при этом остались связанными своими центромерами, так что в ядре имеется по четыре набора каждой хромосомы. Увеличена масса клетки и ее органелл.
Каждое из двух делений мейоза (деления I и II) имеет свои отличительные черты.
Особенность деления I состоит в необычном и сложном прохождении профазы (профаза I). Она подразделяется на несколько стадий: пролептонему, леп-тонему, зигонему, пахинему, диплонему и диакинез.
Во время пролептонемы (от греч. pro — период, leptos — тонкий, пета — нить) происходит значительная, но не полная спирализация хромосом. Ядерная оболочка сохраняется, ядрышко не распадается. Поэтому во время профазы мейоза возможны синтезы некоторых РНК и белков. За счет этих синтезов в половых клетках (особенно в женской) создаются запасы веществ, которые будут необходимы для оплодотворения и ранних стадий развития зародыша.
Во время лептонемы хромосомы еще больше спирализуются, и в ядре становятся видными тонкие нитевидные d-хромосомы (их 46, т. е. два набора). Подчеркнем, что каждая гомологичная хромосома уже реплицировна и состоит из двух сестринских хроматид. Каждая хромосома представляет собой тонкую фибриллу, состоящую из осевой белковой нити, к которой прикрепляется хроматин сестринских хроматид (петли ДНК). Хромосомы с помощью белковых скоплений — прикрепительных дисков — закреплены обоими своими концами на внутренней мембране ядерной оболочки (ядерная оболочка сохраняется, ядрышко хорошо видно).
Во время зигонемы (от греч. zygon — парный) гомологичные диплоидные хромосомы выстраиваются рядом, обвивают друг друга, укорачиваются и сцепляются между собой (конъюгация). Образуются так называемые тетраплоидные биваленты (от лат. Ы — двойной, valens — сильный). Напомним, что каждая диплоидная хромосома из одного бивалента происходит либо от отца, либо от матери. Половые хромосомы располагаются около внутренней ядерной мембраны. Область, занятая ими, называется половым пузырьком.
В зигонеме гомологичные d-хромосомы выстраиваются рядом, сближаются, между ними образуются специализированные синаптонемальные комплексы (от греч. synapsis — связь, соединение), которые представляют собой белковые структуры. При небольшом электронно-микроскопическом увеличении синаптонемальный комплекс выглядит в виде двух электронно-плотных полос, разделенных светлой полосой. При большом увеличении в комплексе видны две параллельные боковые белковые нити длиной 120—150 нм и толщиной 10 нм каждая, соединенные тонкими поперечными полосами размерами около 7 нм, по обе стороны от них лежат d-хромосомы. Их ДНК формирует множество петель.
В центре комплекса проходит осевой элемент толщиной 20—40 нм. Синаптонемальный комплекс удачно сравнивают с веревочной лестницей, стороны которой образованы гомологичными хромосомами. Именно в результате этого гомологичные хромосомы сцепляются между собой и образуют биваленты, 46 d-хромосом образуют 23 бивалента. Каждый бивалент состоит из двух d-хромосом, т. е. из четырех хроматид.
К концу зиготены каждая пара гомологичных хромосом связана между собой с помощью синаптонемальных комплексов. Лишь половые хромосомы X и Y конъюгируют неполностью, т. к. они неполностью гомологичны.
Пахинема (от греч. pahys — толстый) продолжается не менее, чем несколько суток. Процессы развертываются постепенно. Хромосомы несколько укорачиваются и утолщаются. Между хроматидами материнского и отцовского происхождения в нескольких местах возникают соединения — хиазмы (от греч. chiasma — перекрест), или рекомбинантные узелки. Они представляют собой белковые комплексы размерами около 90 нм. В области каждой хиазмы происходит обмен соответствующих участков гомологичных хромосом — от отцовской к материнской и наоборот. Этот процесс называют кроссинговером (от англ. crossing-over — перекрест). Таким образом, кроссинговер обеспечивает многочисленные генетические рекомбинации. В каждом биваленте человека в профазе I кроссинговер происходит в среднем в двух-трех участках. Количество рекомбинантных узелков равно количеству перекрестков.
По окончании кроссинговера хроматиды разъединяются, но остаются связанными в области хиазм. Наступает стадия диплонемы.
В диплонеме (от греч. diploos — двойной) синаптонемальные комплексы распадаются, конъюгировавшие хромосомы раздвигаются, и гомологичные хромосомы каждого бивалента отодвигаются друг от друга, но связь между ними сохраняется в зонах хиазм.
Между диплонемой и диакинезом нет четкой морфологической границы, равно как и разграничений во времени.
диакинезе продолжается конденсация хромосом, они отделяются от нуклеолеммы, но гомологичные d-хромосомы продолжают еще оставаться связанными между собой хиазмами, а сестринские хроматиды каждой d-хромосомы — центромерами. Благодаря наличию нескольких хиазм биваленты образуют петли. В это время разрушаются ядерная оболочка и ядрышки. Реплицированные центриоли направляются к полюсам, образуется веретено деления.
Вследствие сильно затянутой диплонемы профаза мейоза очень длительна. При развитии спермиев она может длиться несколько суток, а при развитии яйцеклеток — в течение многих лет. Половые клетки в профазе мейоза называются гаметоцитами первого порядка (первичными га-метоцитами, гаметоцитами I).
Метафаза I напоминает аналогичную стадию митоза. Хромосомы устанавливаются в экваториальной плоскости, образуя метафазную пластинку.
В анафазе I хиазмы распадаются, гомологичные хромосомы отделяются друг от друга и расходятся к полюсам. Центромеры этих хромосом не реплицируются- сестринские хромосомы не расходятся
В телофазе I формируется ядерная оболочка и ядрышко, образуется и углубляется борозда деления, происходит карикенез.
Интерфаза II очень короткая. ДНК не реплицируется.
Профаза II не длительная, и коньюгация хромосом при этом не наступает.
В метафазе II 23 хромосомы выстраиваются в области экватора.
В анафазе II ДНК в области центромеры реплицируется, хромосомы расходятся к полюсам клетки.
В телофазе II образуются две дочерние клетки.
Итак, в результате двух последовательных делений мейоза образуются 4 клетки, каждая из которых несет гаплоидный набор хромосом.
Как известно, клетки не возникают сами по себе, а образуются только при делении других. После деления во вновь образованной клетке не всегда сразу существуют все системы, обеспечивающие ее специфическую функцию. Должно пройти некоторое время, чтобы сформировались все органеллы и были бы синтезированы все необходимые ферменты. Этот отрезок времени называется созреванием. Созревание клетки осуществляется на основе уже сложившейся ее полной детерминации.
Большинство клеток, однако, погибает тогда, когда проявляются особые естественные генетические механизмы. Генетически запрограммированную клеточную гибель называют апоптозом. Механизм возникновения апоптоза весьма сложен. Каждая клетка несет в хромосомах гены, которые могут запускать синтез ферментов, стимулирующих ее к делению. Есть также гены, которые обеспечивают синтез ферментов, препятствующих делению. Пока клетка функционирует, эти синтезы уравновешены.
Для поддержания жизненного равновесия клетка должна также получать сигналы от других клеток, нередко другого вида. Обычно в качестве сигнальных выступают специфические молекулы олигопептидов. Поскольку они поддерживают жизнь клеток, их назвали цитокинами.
В жизненном цикле многих видов клеток наступает момент, когда функциональные их возможности исчерпываются. У таких клеток нарушается чувствительность к цитокинам и изменяется соотношение активности генов, обеспечивающих внутреннее равновесие. Гены, обеспечивающие размножение клетки, блокируются. Напротив, гены, обеспечивающие синтез литических ферментов, стимулируются. Последние поступают в ядро и лизируют хроматин.
Хромосомы распадаются, синтезы в клетке прекращаются. Внешние проявления такой гибели клеток разнообразны и известны давно. Их называли пикнозом (сморщивание ядра), хроматолизисом (снижение окрашиваемости ядра), кариорексисом (распад ядра на части). Лишь недавно было показано, что это лишь частные проявления апоптоза. Вслед за гибелью ядра разрушается и цитоплазма. Остатки фагоцитируются макрофагами. Материал погибших клеток перерабатывается макрофагами и может выводиться ими на поверхность. В таком случае этот материал может опять использоваться другими клетками. Вокруг клеток, подвергшихся апоптозу, воспалительный процесс не возникает, и жизнедеятельность ткани, часть которой составляли погибшие клетки, продолжается без нарушений.

Источник: StudFiles.net
Фазы мейоза
У организмов, размножающихся половым путем, имеется две категории клеток: диплоидные и гаплоидные. К первым относятся соматические и предшественницы половых клеток, ко вторым — зрелые половые (гаметы). Уменьшение количества хромосом в два раза достигается благодаря мейозу. Он включает в себя два последовательных деления. После слияния гамет возникает новый одноклеточный диплоидный организм (зигота), который несет не просто сумму признаков своих родителей, а является индивидуумом с присущими только ему свойствами.
При дальнейшем митотическом делении зиготы образуются диплоидные же клетки, содержащие по два экземпляра каждой хромосомы, которые называются гомологичными. Гомологичные хромосомы, имеющие одинаковую длину и одинаковое расположение центромер, содержат одинаковое количество генов, а эти гены имеют одну и ту же линейную последовательность. Каждая из пары гомологичных хромосом диплоидного организма происходит либо из ядра спермия, либо из ядра яйцеклетки.
При образовании гамет в зрелом организме в результате мейоза в каждую дочернюю клетку от всех пар гомологичных хромосом попадает лишь по одной из них. Это становится возможным потому, что при мейозе происходит лишь одна репликация ДНК, за которой следуют два последовательных деления ядер (мейоз I и II) без повторного синтеза ДНК. В результате из одной диплоидной образуются четыре гаплоидные клетки.
Напомним, что перед началом мейоза в интерфазе клетка прошла обычные фазы Gx, S и G2, так что стала тетраплоидной. Иначе говоря, произошла репликация ДНК и бел-ков-гистонов хромосом, а сестринские хроматиды при этом остались связанными своими центромерами, так что в ядре имеется по четыре набора каждой хромосомы. Увеличена масса клетки и ее органелл.
Каждое из двух делений мейоза (деления I и II) имеет свои отличительные черты.
Особенность деления I состоит в необычном и сложном прохождении профазы (профаза I). Она подразделяется на несколько стадий: пролептонему, леп-тонему, зигонему, пахинему, диплонему и диакинез.
Во время пролептонемы (от греч. pro — период, leptos — тонкий, пета — нить) происходит значительная, но не полная спирализация хромосом. Ядерная оболочка сохраняется, ядрышко не распадается. Поэтому во время профазы мейоза возможны синтезы некоторых РНК и белков. За счет этих синтезов в половых клетках (особенно в женской) создаются запасы веществ, которые будут необходимы для оплодотворения и ранних стадий развития зародыша.
Во время лептонемы хромосомы еще больше спирализуются, и в ядре становятся видными тонкие нитевидные d-хромосомы (их 46, т. е. два набора). Подчеркнем, что каждая гомологичная хромосома уже реплицировна и состоит из двух сестринских хроматид. Каждая хромосома представляет собой тонкую фибриллу, состоящую из осевой белковой нити, к которой прикрепляется хроматин сестринских хроматид (петли ДНК). Хромосомы с помощью белковых скоплений — прикрепительных дисков — закреплены обоими своими концами на внутренней мембране ядерной оболочки (ядерная оболочка сохраняется, ядрышко хорошо видно).
Во время зигонемы (от греч. zygon — парный) гомологичные диплоидные хромосомы выстраиваются рядом, обвивают друг друга, укорачиваются и сцепляются между собой (конъюгация). Образуются так называемые тетраплоидные биваленты (от лат. Ы — двойной, valens — сильный). Напомним, что каждая диплоидная хромосома из одного бивалента происходит либо от отца, либо от матери. Половые хромосомы располагаются около внутренней ядерной мембраны. Область, занятая ими, называется половым пузырьком.
В зигонеме гомологичные d-хромосомы выстраиваются рядом, сближаются, между ними образуются специализированные синаптонемальные комплексы (от греч. synapsis — связь, соединение), которые представляют собой белковые структуры. При небольшом электронно-микроскопическом увеличении синаптонемальный комплекс выглядит в виде двух электронно-плотных полос, разделенных светлой полосой. При большом увеличении в комплексе видны две параллельные боковые белковые нити длиной 120—150 нм и толщиной 10 нм каждая, соединенные тонкими поперечными полосами размерами около 7 нм, по обе стороны от них лежат d-хромосомы. Их ДНК формирует множество петель.
В центре комплекса проходит осевой элемент толщиной 20—40 нм. Синаптонемальный комплекс удачно сравнивают с веревочной лестницей, стороны которой образованы гомологичными хромосомами. Именно в результате этого гомологичные хромосомы сцепляются между собой и образуют биваленты, 46 d-хромосом образуют 23 бивалента. Каждый бивалент состоит из двух d-хромосом, т. е. из четырех хроматид.
К концу зиготены каждая пара гомологичных хромосом связана между собой с помощью синаптонемальных комплексов. Лишь половые хромосомы X и Y конъюгируют неполностью, т. к. они неполностью гомологичны.
Пахинема (от греч. pahys — толстый) продолжается не менее, чем несколько суток. Процессы развертываются постепенно. Хромосомы несколько укорачиваются и утолщаются. Между хроматидами материнского и отцовского происхождения в нескольких местах возникают соединения — хиазмы (от греч. chiasma — перекрест), или рекомбинантные узелки. Они представляют собой белковые комплексы размерами около 90 нм. В области каждой хиазмы происходит обмен соответствующих участков гомологичных хромосом — от отцовской к материнской и наоборот. Этот процесс называют кроссинговером (от англ. crossing-over — перекрест). Таким образом, кроссинговер обеспечивает многочисленные генетические рекомбинации. В каждом биваленте человека в профазе I кроссинговер происходит в среднем в двух-трех участках. Количество рекомбинантных узелков равно количеству перекрестков.
По окончании кроссинговера хроматиды разъединяются, но остаются связанными в области хиазм. Наступает стадия диплонемы.
В диплонеме (от греч. diploos — двойной) синаптонемальные комплексы распадаются, конъюгировавшие хромосомы раздвигаются, и гомологичные хромосомы каждого бивалента отодвигаются друг от друга, но связь между ними сохраняется в зонах хиазм.
Между диплонемой и диакинезом нет четкой морфологической границы, равно как и разграничений во времени. В диакинезе продолжается конденсация хромосом, они отделяются от нуклеолеммы, но гомологичные d-хромосомы продолжают еще оставаться связанными между собой хиазмами, а сестринские хроматиды каждой d-хромосомы — центромерами. Благодаря наличию нескольких хиазм биваленты образуют петли. В это время разрушаются ядерная оболочка и ядрышки. Реплицированные центриоли направляются к полюсам, образуется веретено деления.
Вследствие сильно затянутой диплонемы профаза мейоза очень длительна. При развитии спермиев она может длиться несколько суток, а при развитии яйцеклеток — в течение многих лет. Половые клетки в профазе мейоза называются гаметоцитами первого порядка (первичными га-метоцитами, гаметоцитами I).
Метафаза I напоминает аналогичную стадию митоза. Хромосомы устанавливаются в экваториальной плоскости, образуя метафазную пластинку.
В анафазе I хиазмы распадаются, гомологичные хромосомы отделяются друг от друга и расходятся к полюсам. Центромеры этих хромосом не реплицируются- сестринские хромосомы не расходятся
В телофазе I формируется ядерная оболочка и ядрышко, образуется и углубляется борозда деления, происходит карикенез.
Интерфаза II очень короткая. ДНК не реплицируется.
Профаза II не длительная, и коньюгация хромосом при этом не наступает.
В метафазе II 23 хромосомы выстраиваются в области экватора.
В анафазе II ДНК в области центромеры реплицируется, хромосомы расходятся к полюсам клетки.
В телофазе II образуются две дочерние клетки.
Итак, в результате двух последовательных делений мейоза образуются 4 клетки, каждая из которых несет гаплоидный набор хромосом.
Как известно, клетки не возникают сами по себе, а образуются только при делении других. После деления во вновь образованной клетке не всегда сразу существуют все системы, обеспечивающие ее специфическую функцию. Должно пройти некоторое время, чтобы сформировались все органеллы и были бы синтезированы все необходимые ферменты. Этот отрезок времени называется созреванием. Созревание клетки осуществляется на основе уже сложившейся ее полной детерминации.
Большинство клеток, однако, погибает тогда, когда проявляются особые естественные генетические механизмы. Генетически запрограммированную клеточную гибель называют апоптозом. Механизм возникновения апоптоза весьма сложен. Каждая клетка несет в хромосомах гены, которые могут запускать синтез ферментов, стимулирующих ее к делению. Есть также гены, которые обеспечивают синтез ферментов, препятствующих делению. Пока клетка функционирует, эти синтезы уравновешены.
Для поддержания жизненного равновесия клетка должна также получать сигналы от других клеток, нередко другого вида. Обычно в качестве сигнальных выступают специфические молекулы олигопептидов. Поскольку они поддерживают жизнь клеток, их назвали цитокинами.
В жизненном цикле многих видов клеток наступает момент, когда функциональные их возможности исчерпываются. У таких клеток нарушается чувствительность к цитокинам и изменяется соотношение активности генов, обеспечивающих внутреннее равновесие. Гены, обеспечивающие размножение клетки, блокируются. Напротив, гены, обеспечивающие синтез литических ферментов, стимулируются. Последние поступают в ядро и лизируют хроматин.
Хромосомы распадаются, синтезы в клетке прекращаются. Внешние проявления такой гибели клеток разнообразны и известны давно. Их называли пикнозом (сморщивание ядра), хроматолизисом (снижение окрашиваемости ядра), кариорексисом (распад ядра на части). Лишь недавно было показано, что это лишь частные проявления апоптоза. Вслед за гибелью ядра разрушается и цитоплазма. Остатки фагоцитируются макрофагами. Материал погибших клеток перерабатывается макрофагами и может выводиться ими на поверхность. В таком случае этот материал может опять использоваться другими клетками. Вокруг клеток, подвергшихся апоптозу, воспалительный процесс не возникает, и жизнедеятельность ткани, часть которой составляли погибшие клетки, продолжается без нарушений.
Источник: StudFiles.net