Гаплоидная стадия преобладает в жизненном цикле
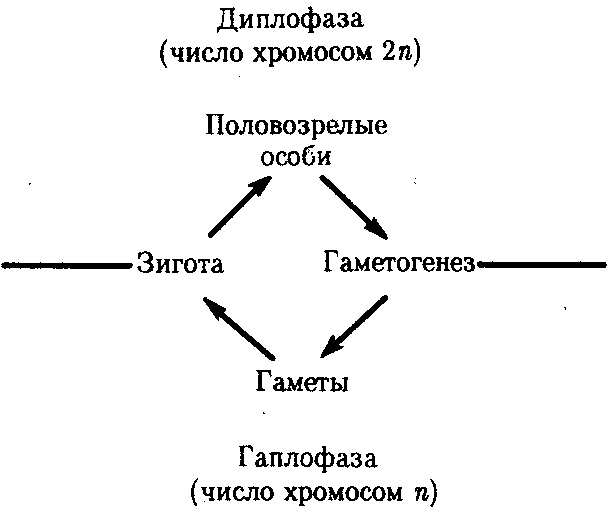
Организмы с половым размножением в результате мейоза образуют гаплоидные гаметы. В момент оплодотворения благодаря слиянию двух таких клеток возникает диплоидная зигота. Многократные митотиче-ские деления зиготы и ее потомков приводят к увеличению числа диплоидных клеток, которые строят тело организма в процессе его развития. По достижении последним половой зрелости возобновляется процесс образования гаплоидных гамет. Таким образом, в жизненных циклах организмов, размножающихся половым способом, выделяются две фазы: гаплоидная и диплоидная (рис. 5.11). Относительная продолжительность этих фаз варьирует у представителей различных групп живых существ: у простейших и грибов преобладает первая, у высших растений и животных — вторая.
Удлинение диплофазы в ходе эволюции объясняется преимуществами диплоидного состояния перед гаплоидным. Благодаря гетерозиготности и рецессивности в диплоидном состоянии укрываются от естественного отбора, сохраняются и накапливаются разнообразные аллели. Это повышает объем генетической информации в генофондах популяций и видов, ведет к образованию резерва наследственной изменчивости, от которого зависят эволюционные перспективы. В то же время у гетерозигот вредные рецессивные аллели, не оказывая влияния на развитие фенотипа, не снижают жизнеспособности.

Рис. 5.11. Фазы жизненного цикла многоклеточных животных
5.5. Пути приобретения организмами биологической информации
Благодаря генетической рекомбинации, которая закономерно происходит в процессе гаметогенеза и при оплодотворении, половое размножение представляет собой эволюционно обусловленный механизм обмена генетической информацией между организмами одного биологического вида. Некоторые факты из области зоологии и особенно вирусологии и микробиологии указывают и на то, что имеются пути приобретения биологической информации и от организмов других видов (см. 3.6.4.5). Эта информация воспроизводится в фенотипе организма и определяет развитие признаков, не закодированных в генетическом материале родителей. Так, в клетках пищеварительного дивертикула брюхоногого моллюска Elysia viridis сохраняются хлоропласты поедаемой водоросли Codium bragile, в результате чего моллюск приобретает способность к фотосинтезу. Стрекательные капсулы гидроидных полипов, которые поедаются некоторыми реснитчатыми червями, не перевариваются, а перемещаются в эпителиальный пласт и используются червем в качестве орудия защиты. В классической зоологии такие примеры получили название клептогенеза или эволюции путем воровства.
Явление трансдукции заключается в том, что в генетический материал клетки-хозяина (бактериальной или эукариотической) встраивается нуклеиновая кислота вируса с фрагментом генома другой клетки. Привносимая таким образом биологическая информация вследствие редупликации чужеродной ДНК может передаваться в ряду клеточных поколений, а также воздействовать на состояние генетической системы клетки-хозяина, изменяя, например, частоту мутирования отдельных генов. Чужеродная ДНК может присутствовать в клетке в виде плазмид и эписом — фрагментов нуклеиновой кислоты, лишенных в отличие от вирусных частиц белковых чехлов. Плазмиды самостоятельны по отношению к хромосомам клетки-хозяина, а эписомы могут встраиваться в них. Биологическая информация плазмид и эписом, проявляясь в фенотипе, дает широкий круг признаков, включая устойчивость к антибиотикам (см. 3.6.3).
Примеры проникновения в организм действующей биологической информации организмов из других таксонов, прежде всего вирусов, описаны у высших животных и человека.
к, сотрудники, длительно работающие в онкологических лабораториях с вирусной опухолью кроликов — папилломой Шопа, имеют, как правило, пониженное содержание в плазме крови аминокислоты аргинина. Объясняется это тем, что вирус папилломы, которым «заражены» такие люди, несет ген синтеза аргиназы, катализирующей обмен аргинина. Будучи активным, этот ген обусловливает образование дополнительных молекул фермента. Описанный факт относится к широкой области так называемого вирусоносительства или латентных вирусных инфекций. Вирус, присутствуя в клетках организма длительное время и не вызывая собственно патологических изменений, приводит к развитию некоторых фенотипических признаков. У человека известен генетический дефект, который проявляется в серьезных нарушениях развития и зависит от недостатка аргиназы. Открытие способа контролируемого «заражения» таких людей вирусом папилломы Шопа могло бы нормализовать их фенотип.
Технологии геномики (см. разд. 3.2), расшифровывающие нуклеотидные последовательности ДНК, создают почву для целенаправленного изменения или введения в человеческий геном нормальных нуклеотидных последовательностей, что в перспективе может стать основой генотерапии.
Половой процесс служит универсальным механизмом обмена генетической информацией между особями в пределах вида. Рассмотренные факты свидетельствуют о том, что количество и содержание информации, используемой различными организмами для развития и жизнедеятельности, не всегда ограничиваются той, которая была ими приобретена от родителей.
ществуют дополнительные пути приобретения биологической информации. Видоспецифическая информация, получаемая в процессе размножения, обеспечивает развитие особей определенного морфофизиологического типа. Дополнительная информация по биологическому содержанию в значительной степени случайна и нередко нарушает реализацию собственной генетической информации хозяина. В связи с этим в эволюции возникли механизмы защиты от проникновения чужеродного наследственного материала. Примером может служить интерферон — белок, вырабатываемый клетками млекопитающих и птиц в ответ на внедрение вирусов и представляющий собой фактор неспецифического противовирусного иммунитета, а также система иммунологического надзора.
Источник: StudFiles.net
Виды плоидности и терминология
- Гаплоидные клетки — содержат одинарный набор непарных хромосом (половые клетки, прокариоты).
- Диплоидные клетки — содержат парное количество хромосом. Большая часть организмов, размножающихся половым путём, диплоидны, т. е. содержат в соматических клетках тела по одному набору хромосом от каждой из гамет (гаплоидных половых клеток).
- Полиплоидные клетки — содержат более чем две пары хромосом (до двенадцати пар). В зависимости от того, сколько раз в ядре клетки повторяется гаплоидный набор, их соответственно называют три-, тетра-, гексаплоидными и т.
Полиплоидия возникает вследствие нарушения хода митоза или мейоза (значительно реже) под воздействием мутагенов: при разрушении веретена деления удвоившиеся хромосомы не расходятся, а остаются внутри неразделившейся клетки (так возникают гаметы с двукратным числом хромосом — 2n). При слиянии такой гаметы с нормальной (n) потомок будет иметь тройной набор хромосом и т.д. Полиплоидия имеет две разновидности:- Автополиплоидия — результат кратного увеличения гаплоидного набора хромосом одного вида.
- Аллополиплоидия — результат объединения наборов хромосом разных видов после образования межвидовых гибридов.
- Анеуплоидные клетки — непропорциональное (не кратное гаплоидному) удвоение или утрата отдельных хромосом. В зависимости от того, произошло уменьшение или увеличение хромосом, используют соответственно приставки гипо- и гипер-. Например, гипердиплоиды — трисомики (2n +1) и тетрасомики (2n + 2), гиподиплоиды — моносомики (2n — 1) и нуллисомики (2n — 2). Анеуплоидия как правило появляется из-за влияния мутагенов.
Иногда термин «плоидность» применяют не только к эукариотам, но и в отношении безядерных прокариотов, которые как правило гаплоидны, однако иногда встречаются диплоидные и полиплоидные бактерии.
Полиплоидию не следует путать с увеличением количества ядер в клетке и увеличением числа молекул ДНК (политенизацией) в хромососоме.
Гаплоидная и диплоидная фазы в жизненном цикле
У раздельнополых организмов в жизненном цикле происходит как правило нормальное чередование гаплоидной и диплоидной фаз. При мейозе образуются гаплоидные клетки в результате разделения диплоидной (у некоторых растений и грибов затем может происходить размножение путём митоза с образованием гаплоидного многоклеточного тела или нескольких поколений гаплоидных клеток-потомков). В результате полового процесса хромосомы двух гаплоидных клеток объединяются в одной диплоидной (зиготе), после чего могут размножаться при помощи митоза (у растений и животных) с образованием диплоидного многоклеточного тела или диплоидных клеток-потомков.
Полиплоидия у растений
Термин полиплоидия был предложен в 1916 году немецким ученым Винклером, изучавшим образцы аномальных (химерных) тканей у паслена.
Естественная полиплоидность в природе распространена достаточно широко. До 75% арктический флоры – полиплоиды, так же велик процент полиплоидов в пустынных и высокогорных регионах, где выживают растения, устойчивые к экстремальным условиям обитания.
Человеком полиплоидия используется издавна. Сначала просто размножали самые крупные экземпляры, дающие много зерна или же хорошие плоды. С развитием генетики выяснилось, что такие гиганты – отобранные природные полиплоиды. В настоящее время на основе искусственной автополиплоидии синтезированы высокоурожайные формы и сорта пшеницы, ржи, гречихи, кукурузы, картофеля, хлопчатника, сахарной свеклы, сахарного тростника и других культурных растений.
стения-полиплоиды как правило характеризуются более крупными размерами, повышенным содержанием ряда веществ, устойчивостью к неблагоприятным факторам внешней среды, отличными от исходных форм сроками цветения и плодоношения. Искусственная полиплоидия вызывается ядами, разрушающими веретено деления, такими как алкалоид колхицин.
Аллополиплоидия (межвидовое скрещивание) обычно возникает от удвоения хромосом гибрида двух видов, что приводит к его плодовитости (амфиплоидия). Пример природной аллополиплоидии – алыча, гибрид терна и дикой сливы, полученный тысячелетия назад в результате естественной гибридизации. Искусственный гибрид получен в 1928 году русским цитогенетиком Карпеченко, который скрестил редьку с капустой. Полученый «амфидиплоид» получил научное название Paphanobrassica. У этого растения листья были как у редьки, а корни напоминали капустные. Хотя экономической ценностью полученный гибрид не обладает, зато позиционируется эволюционистами в качестве доказательства реальности биологической эволюции. В этом случае стоит отметить, что Paphanobrassica имела признаки обеих видов-прародителей, но не обладало принципиально новыми признаками, которые бы указывали на возможность прогрессивных макроэволюционных изменений.
Полиплоидия у животных
В животном мире полиплоиды встречаются среди нематод, аскарид, пиявок, земноводных. У многих млекопитающих полиплоидные клетки встречаются в отдельных органах (печень, и др.), но пример полной полиплоидии известен лишь один – южноамериканский грызун Tympanoctomys barrerae (вид, родственный морским свинкам и шиншиллам).
Нарушения плоидности у человека
У человека большая часть клеток диплоидны. Гаплоидны только зрелые половые клетки (гаметы). Другие варианты плоидности — несут лишь отрицательное воздействие.
Примеры анеуплоидии у человека: синдром Дауна (21-я хромосома представлена тремя копиями), синдром Кляйнфельтера — избыточная X хромосома (XXY), синдром Тернера — отсутствие одной из половых хромосом (X0). Описаны также примеры утроения X хромосомы и некоторые другие аномалии.
Примерами полиплоидии являются абортивные триплоидные зародыши и триплоидные новорождённые (срок их жизни при этом не превышает нескольких дней), а также диплоидно-триплоидные мозаики.
Полиплодия в теории креационизма
Казалось бы, примеры с удачными гибридами неоспоримо доказывают, что увеличение числа хромосом — путь к эволюционному прогрессу. Однако наблюдение полиплоидии в природе приводит к интересным, а иногда — и к противоположным выводам. В частности Кент Ховант в своих лекциях (1999 г.) любил приводить факты о количестве хромосом в соматических клетках разных организмов. Если бы количество хромосом имело смысл в эволюции, тогда по правилу элементарной логики, чем больше хромосом, тем дальше живое существо взобралось по древу эволюции. Но это не так.
| Царство растений | число хромосом | Животные и человек | число хромосом |
|---|---|---|---|
| Помидор | 12 | Домашняя муха | 12 |
| Горох | 14 | Опоссум | 22 |
| Капуста | 18 | Лягушка | 26 |
| Морковь | 20 | Летучая мышь | 44 |
| Бобы | 22 | Человек | 46 |
| Секвойя | 22 | Шимпанзе | 48 |
| Лук | 32 | Амёба | 50 |
| Соя | 40 | Собака | 78 |
| Табак | 48 | Курица | 78 |
| Папоротник | 480 | Карп | 100 |
Таким образом полиплоидия ещё ждёт своего научного осмысления.
Источник: creationwiki.org
СПОСОБЫ И ФОРМЫ РАЗМНОЖЕНИЯ
В зависимости от характера клеточного материала, используемого в целях размножения, выделяют различные способы и формы последнего. Различают два способа размножения: бесполое и половое Деление надвое приводит к возникновению из одного родительского организма двух дочерних. Оно является преобладающей формой у прокариот и простейших, но встречается и у многоклеточных: продольное у медуз, поперечное у кольчатых червей.
>Множественное деление (шизогония) встречается среди простейших, в том числе паразитов человека (малярийный плазмодий). При размножении почкованием потомок формируется первоначально как вырост на теле родителя с последующей его отшнуровкой (гидра). Фрагментация заключается в распаде тела многоклеточного организма на части, которые далее превращаются в самостоятельных особей (плоские черви, иглокожие). У видов, размножающихся спорами, дочерний организм развивается из специализированной клетки-споры. В зависимости от формы бесполого размножения потомок развивается либо из одной клетки (спорообразование, шизогония, деление), либо из группы клеток родителя. В последнем случае размножение называют вегетативным. Оно распространено среди растений. Бесполое размножение наблюдается у животных с относительно низким уровнем структурно-физиологической организации, к которым принадлежат многие паразиты человека. У паразитов бесполое размножение не только служит увеличению численности особей, но способствует расселению, помогает пережить неблагоприятные условия.
ПОЛОВОЕ РАЗМНОЖЕНИЕ
Хотя в процессе развития жизни бесполое размножение возникло первьш, половое размножение существует на Земле уже более 3 млрд. лет. Оно обнаруживается в жизненных циклах всех основных групп организмов. Распространенность полового размножения объясняется тем, что оно обеспечивает значительное генетическое разнообразие и, следовательно, фенотипическую изменчивость потомства. Этим достигаются большие эволюционные и экологические (расселение в разные среды) возможности. В основе полового размножения лежит половой процесс, суть которого сводится к объединению в наследственном материале для развития потомка генетической информации от двух разных источников — родителей. Представление о половом процессе дает явление конъюгации, например инфузорий. Он заключается во временном соединении двух особей с целью обмена (рекомбинации) наследственным материалом. В результате появляются особи, генетически отличные от родительских организмов. В дальнейшем они осуществляют бесполое размножение. Поскольку количество инфузорий после конъюгации остается неизменным, говорить о размножении в прямом смысле нет оснований. У простейших половой процесс может осуществляться в виде копуляции, которая заключается в слиянии двух особей в одну, объединении и рекомбинации наследственного материала. Далее такая особь размножается делением. На определенном этапе эволюции у многоклеточных организмов половой процесс как способ обмена генетической информацией между особями в пределах вида оказался связанным с размножением. Для участия в половом размножении в родительских организмах вырабатываются гаметы —клетки, специализированные к обеспечению генеративной функции. Слияние материнской и отцовской гамет приводит к возникновению зиготы — клетки, представляющей собой дочернюю особь на первой, наиболее ранней стадии индивидуального развития. У некоторых организмов зигота образуется в результате объединения гамет, неотличимых по строению. В таких случаях говорят об изогамии, У большинства видов по структурным и функциональным признакам половые клетки делятся на материнские (яйцеклетки) и отцовские (сперматозоиды). Как правило, яйцеклетки и сперматозоиды вырабатываются разными организмами — женскими (самки) и мужскими (самцы). В подразделении гамет на яйцеклетки и сперматозоиды, а особей на самок и самцов заключается явление полового диморфизма. Наличие его в природе отражает различия в задачах, решаемых в процессе полового размножения мужской или женской гаметой, самцом или самкой. Образование гамет обоих видов в одном организме, имеющем и мужскую, и женскую половые железы, называют гермафродитизмом
ЧЕРЕДОВАНИЕ ГАПЛОИДНОЙ И ДИПЛОИДНОЙ ФАЗ ЖИЗНЕННОГО ЦИКЛА
Организмы с половым размножением в результате мейоза образуют гаплоидные гаметы. В момент оплодотворения благодаря слиянию двух таких клеток возникает диплоидная зигота. Многократные митотиче-ские деления зиготы и ее потомков приводят к увеличению числа диплоидных клеток, которые строят тело организма в процессе его развития. По достижении последним половой зрелости возобновляется процесс образования гаплоидных гамет. Таким образом, в жизненных циклах организмов, размножающихся половым способом, выделяются две фазы: гаплоидная и диплоидная (рис. 5.11). Относительная продолжительность этих фаз варьирует у представителей различных групп живых существ: у простейших и грибов преобладает первая, у высших растений и животных — вторая. Удлинение диплофазы в ходе эволюции объясняется преимуществами диплоидного состояния перед гаплоидным. Благодаря гетерозиготности и рецессивности в диплоидном состоянии укрываются от естественного отбора, сохраняются и накапливаются разнообразные аллели. Это повышает объем генетической информации в генофондах популяций и видов, ведет к образованию резерва наследственной изменчивости, от которого зависят эволюционные перспективы. В то же время у гетерозигот вредные рецессивные аллели, не оказывая влияния на развитие фенотипа, не снижают жизнеспособности.
Источник: studopedia.ru
В норме у большинства организмов, для которых известен половой процесс, в жизненном циклепроисходит правильное чередование гаплоидной и диплоидной фаз. Гаплоидные клетки образуются в результате мейотического деления диплоидных клеток, после чего у некоторых организмов (растения, водоросли, грибы) могут размножаться при помощи митотических делений с образованием гаплоидного многоклеточного тела или нескольких поколений гаплоидных клеток-потомков. Диплоидные клетки образуются из гаплоидных в результате полового процесса (слияния половых клеток, или гамет) с образованием зиготы, после чего могут размножаться при помощи митотических делений (у растений, водорослей и некоторых других протистов, животных) с образованием диплоидного многоклеточного тела или диплоидных клеток-потомков.
20. Вероятно, в то время в пресных водах суши уже обитали многочисленные организмы: из растений — зеленые и сине-зеленые водоросли, бактерии, низшие грибы, из животных — одноклеточные корненожки, жгутиковые, реснитчатые инфузории, различные другие беспозвоночные, кишечнополостные и черви. Обитателей пресных вод можно по праву назвать пионерами жизни на суше.
Именно в исчезающих, окраинных участках морей и других водоемов сама природа вынуждала некоторые виды водяных растений (зеленые водоросли и др.) приспосабливаться к жизни вне воды. В периоды мелководий и засух водные растения, у которых лучше были развиты предвестники будущих корней, выживали. В течение тысячелетий растения постепенно расселялись в прибрежной полосе суши, дав начало наземному растительному миру.
21. Иногда скелет растворяется, и от него в породе остается лишь слепок, который называют ядром. Он образован минеральным веществом, принесенным водой. Особенно хорошо такие ядра образуются при растворении различных раковин. Часто от скелета остается в породе только отпечаток, по которому уже трудно судить о строении животного.
Порой даже само образование породы связано с массовым скоплением остатков вымерших организмов. Их можно увидеть под микроскопом в препарате из обыкновенного писчего мела. Известен фузулиновый известняк, образованный похожими на крошечные веретенца простейшими организмами — фузулинами, жившими более 200 млн. лет назад. В Крыму встречается нуммулитовый известняк, образованный крупными монетковидными скелетами одноклеточных организмов — нуммулитов, обитавших в теплых морях более 50 млн. лет назад. Не редкость слои известняка, сложенные скелетами вымерших кораллов, которые в морях далекого прошлого образовывали рифы, подобно их потомкам в современных морях.
22. Эукариотные организмы — простейшие, различные группы растений, грибы и многоклеточные животные — доминируют в современной биосфере. Однако все они несут в своих клетках симбионтов — потомков древних свободноживущих бактерий. Только благодаря им эукариотные организмы способны жить в кислородной атмосфере и использовать энергию солнечного света для синтеза органических веществ. Так может быть, на самом деле эукариоты вовсе не доминируют в биосфере? С Американский биолог Л. Томас сказал: „Обычно на митохондрии смотрят как на порабощённые существа, взятые в плен, чтобы снабжать АТФ клетки, и не способные дышать самостоятельно. Но с точки зрения самих митохондрий они — существа, которые давным-давно нашли для себя лучшее из возможных пристанищ, где можно жить, затрачивая минимум усилий и подвергаясь наименьшему риску“.
Важную роль в эволюции эукариот сыгралсимбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и поглощенными этой клеткой бактериями — предшественниками митохондрийи пластидов.
23. Первым на Земле были древние одноклеточные, от которых произошли современные типы одноклеточных — саркодово-жгутиковые, инфузории и споровики. Наиболее сложное строение из ныне живущих одноклеточных имеют многоядерные инфузории и колониальные жгутиковые. От древних колониальных жгутковых с животным типом питания произошли многоклеточные организмы, тело которых состояло из 2-х групп клеток: наружних — жгутиковых и внутренних — пищеварительных. Образование первых многоклеточных животных было крупным событием в историческом развитии животного мира. Многоклеточные животные получили большие преимущества перед одноклеточными: у них появились широкие возможности к разделению клеток по выполняемым функциям, дальнейшему усложнению строения тела, увеличению размеров и регенерации поврежденных и утраченных частей тела.
Одни из самых примитивных многоклеточных животных кишечнополостные — произошли от древних многоклеточных животных, близких к колониальным жгутиковым. На это указывает наличие в из пищеварительном слое жгутиковых клеток. О родстве кишечнополостных с колониальными жгутиковыми говорит и сходство ранних стадий развития гидры с колониальными жгутиковыми, и развитие зародыша от одной клетки_(оплодотворенной яйцеклетки). Это были двухслойные животные.
Дальнейшие развитие животного мира связано с появлением первых трехслойных животных, похожих на примитивных свободноживущих ресничных червей и произошедших от древних примитивных двухслойных животных.
Трехслойное животные получили в процессе исторического развития прогрессивные особенности строения: мышечную систему и паренхиму. Появление мускулатуры обеспечили более быстрое и совершенное передвижение животных, а благодаря паренхиме сформировалось внутренняя среда организма, обеспечивающая более совершенные обмен веществ. К первым трехслойным животным относятся типы плоских и круглых червей.
Следующий этап в развития животного мира связан с появлением трехслойных животных, имеющих полость тела, в которой располагаются внутренние органы. У этих животных впервые появляется кровеносная система, совершенные органы выделения и более сложная нервная система. От примитивных форм полостных трехслойных животных произошли кольчатые черви, а от древних кольчатых червей — моллюски и членистоногие. От примитивных трехслойных полостных животных ведут свое начало и хордовые животные.
Переходные формы не найдены. Многоклеточные в летописи окаменелостей появляются одномоментно, в большом разнообразии семейств и отрядов. Этот феномен, противоречащий всей теории эволюции, получил в среде эволюционистов наименование «Кембрийский взрыв».
24. Гу́бки (лат. Porifera, Spongia) — тип беcпозвоночных животных. В настоящее время описано около 8000 видов. Хотя подавляющее большинство губок обитает в морях, пресноводные представители (например, бадяги) встречены во внутренних водах всех материков кроме Антарктиды.
Губки относятся к наиболее примитивным многоклеточным организмам, не имеют ясно дифференцированных тканей и обособленных органов. Имеют скелетные образования в виде известковых, кремнезёмных игл (спикул) или волокон белка спонгина. Это древняя группа животных, известная с докембрия.
Губку можно схематично представить в виде двух слоёв клеток — эпителиальных и пищеварительных, а между слоями формируется твёрдый скелет из известковых или кремнезёмных игл.
Почкуясь, губки образуют колонии. Одиночные губки имеют высоту от нескольких миллиметров до 3 сантиметров, колонии — до 1,5 м. Распространены широко: от прибрежной зоны до глубины 8500 м.
Тканевая организация губок развита очень слабо; имеются особые клетки, обеспечивающие циркуляцию воды и питание — хоаноциты.
Если губку измельчить, то её клетки могут снова сформировать полноценное животное.
Различают три основных типа организации тела губок: аскон, сикон и лейкон.
Асконоидная организация — наиболее простая. В центре тела губки располагается атриум (камера), окружённая снаружи стенкой тела, покрытой пинакодермой. Стенка тела пронизана многочисленными отверстиями — порами, пропускающими внутрь воду. Клетки пинакодермы — пинакоциты — могут сокращаться, сжимая губку. Под пинакодермой лежит мезохил — студенистый матрикс, содержащий спикулы и тотипотентные амебоциты.
Сикон — вторая по сложности ирригационная система у губок. Отличается от аскона только тем, что внутри стенок пор находятся полости, что способствует увеличению количества пропускаемой воды и соответственно увеличивается количество поглощаемой пищи.
Лейкон — самая сложноустроенная ирригационная система губок. Количество пищи увеличивается за счет сложной формы полостей в стенках пор, что позволяет пропускать и сохранять большое количество пищи.
Научная классификация
Царство: Животные
Подцарство: Паразои
Тип: Губки
Латинское название
Porifera (Grant in Todd, 1836)
Классы
Известковые губки (Calcarea)
Обыкновенные губки (Demospongiae)
Шестилучевые губки (Hexactinellida)
Своеобразную экологическую группу представляют сверлящие губки (род Cliona). Поселяясь на известковом субстрате (раковины моллюсков, колонии кораллов, известковые породы и т.п.), они образуют в нем ходы, открывающиеся наружу небольшими отверстиями ( рис. 88 ). Через эти отверстия выступают выросты тела губки, несущие оскулумы. Механизм действия сверлящих губок на субстрат еще неясен. В растворении извести, по-видимому, существенную роль играет выделяемая губкой углекислота.
Практическое значение губок невелико. В некоторых южных странах имеется промысел обладающих роговым скелетом туалетных губок, используемых для мытья и различных технических целей. Их ловят в Средиземном и Красном морях, Мексиканском заливе, Карибском море, Индийском океане, у берегов Австралии. Промысел стеклянных губок (преимущественно Euplectella), используемых в качестве украшений и сувениров, существует также у берегов Японии
25. Многоклеточные животные. Тип Кишечнополостные
В отличие от клеток Простейших, способных самостоятельно выполнять все жизненно важные функции, клетки многоклеточных организмов выполняют одну определенную функцию. При этом структура их значительно упрощается, но специфическая функция выполняется клеткой гораздо эффективней.
Многоклеточных животных разделяют на две группы: беспозвоночных и позвоночных. Отличают их по наличию внутреннего осевого скелета — хорды или позвоночника.
Тип Кишечнополостные
Этот тип относится к числу самых древних и примитивных многоклеточных животных. Насчитывается ок. 9 000 видов кишечнополостных. Тип делится на три класса:
* класс Гидроидные, у которых доминирует полип (пресноводная гидра, актинии и колониальные полипы);
* класс Сцифоидные, у которых доминирует стадия медузы (медуза-корнерот и др.);
* класс Коралловые полипы, ведущие колониальный образ жизни (нет стадии медузы).
Тело кишечнополостных построено из двух слоев клеток: наружного — эктодермы и внутреннего — энтодермы. Для большинства видов характерна радиальная симметрия.
Питаются кишечнополостные мелкими водными животными. Пищеварительная система имеет одно отверстие — ротовое, через которое пища попадает в кишечник и выбрасываются наружу непереваренные остатки пищи. Кровеносная, дыхательная и выделительные системы отсутствуют. Дыхание и выделение осуществляются через всю поверхность тела. Большинство кишечнополостных обитает в морях и океанах. К ним относятся полипы и медузы. Некоторые животные рода Гидра обитают в пресных водоемах.
Наружный слой тела гидры (эктодерма) образован клетками нескольких типов. Самые многочисленные — это эпителиально-мускульные (кожно-мускульные) клетки, состоящие из тела и мускулистого отростка. Стрекательные клетки способны реагировать на раздражения из внешней среды, они имеют специальный чувствительный волосок и стрекательную капсулу, внутри которой находится ввернутая длинная стрекательная нить.
Стрекательные клетки постоянно пополняются за счет промежуточных клеток, дающих начало новым клеткам. Из эктодермы образуются нервные клетки (нейроны) с длинными отростками, которыми они соединяются друг с другом, образуя нервную систему диффузного (сетчатого) типа.
Под слоем клеток эктодермы располагаются базальные пластинки из межклеточного вещества (мезоглея). Под ним расположен второй слой клеток -энтодерма, выстилающая кишечную полость полипа. Основная масса внутреннего слоя — это эпителиально-мускульные клетки, отростки которых расположены а поперечном направлении. Работа мускульных отростков эктодермальных и эн-тодермальных клеток согласуется нервной системой, и вместе они могут вытягивать иди сокращать тело полипа, наклонять его в разные стороны. Эпителиально-мускульные клетки энтодермы несут несколько длинных жгутиков и способны образовывать псевдоподии для захвата пищевых частиц. Между этими клетками разбросаны железистые клетки, способные вырабатывать пищеварительные секреты.
Пищеварение идет в два этапа. Полостное пищеварение протекает на кишечной полости. Под действием пищеварительных секретов железистых клеток образуются мелкие пищевые частицы. Внутриклеточное пищеварение проходит внутри эпителиально-мускульных клеток. Не переваренная пища выбрасывается через рот.
Размножаются кишечнополостные бесполым путем — почкованием и половым, с формированием половых клеток. Так, в изгибах нижних веточек колоний полипов находятся бластостилии, от которых отпочковываются небольшие подвижные медузы — свободноживущее плавающее поколение. Медузы обеспечивают расселение животных и способны размножаться половым путем. Таким образом, у разных видов кишечнополостных происходит чередование поколений: полипы — бесполое поколение, размножающееся вегетативно, медузы — половое поколение, вырабатывающее половые клетки. Оплодотворение перекрестное. Из оплодотворенных яйцеклеток развиваются маленькие личинки с ресничками, которые оседают на дно и превращаются в полипов.
У рифообразующих кораллов, как и у всех других кишечнополостных животных, нет специальных органов дыхания. Растворенный в воде кислород они «вдыхают» всей поверхностью тела. Известно, что растворимость газов в воде уменьшается по мере возрастания температуры. Теплые воды тропического океана содержат значительно меньше кислорода, чем холодные воды полярных и умеренных областей. Поэтому многие обитатели тропических морей постоянно ощущают кислородное голодание. По сравнению со многими другими водными беспозвоночными рифообразующие кораллы в этом отношении находятся в гораздо более выгодных условиях, так как дефицитный кислород им поставляют все те же симбиодиниумы. Дело в том, что в процессе фотосинтеза как побочный продукт обязательно образуется кислород, который наземные растения отдают атмосфере, свободноживущие водоросли — воде, а симбиотические — своему хозяину. В течение всего светлого времени, т. е. почти половину суток, кораллы непрерывно получают дополнительный кислород, который поступает прямо в толщу их тканей. В сочетании с обилием питательных веществ это дает им возможность быстрее расти и размножаться.
Сожительство с кораллами дает ряд преимуществ и водорослям. В результате дыхания в тканях кораллов образуется двуокись углерода, которая (в светлое время суток) немедленно используется симбиодиниумами для фотосинтеза. Кроме того, водоросли в процессе синтеза белков утилизируют азот и фосфор, находящиеся в метаболитах (продуктах выделения) кораллов. Наконец, находясь в тканях коралла, симбиодиниумы защищены от множества растительноядных животных.
Однако этим взаимовыгодная связь герматипных кораллов и одноклеточных водорослей не ограничивается. Она имеет первостепенное значение также и в процессе построения твердого известкового скелета кораллов.
В течение всего светлого времени суток симбиодиниумы в процессе фотосинтеза быстро и непрерывно изымают из тканей кораллов двуокись углерода — как ту, что проникает в ткани из морской воды, так и образующуюся в процессе дыхания кораллов. Благодаря этому содержание двуокиси углерода во внутренней среде организма коралла снижается, что способствует сдвигу химических реакций в сторону осаждения извести, т. е. кальцификации. Прямыми наблюдениями было показано, что скорость образования скелета герматипных кораллов идет примерно в 10 раз быстрее, чем у агерматипных — нерифостроящих, лишенных симбиотических водорослей. В ночное время, когда фотосинтез невозможен, образование скелета у герматипных кораллов резко замедляется.
Симбиодиниум не встречается в свободном состоянии, т. е. в морской воде, зато он способен вступать в симбиотическиевзаимоотношения со многими беспозвоночными животными. Кроме мадрепоровых кораллов, его обнаружили в тканях гидрокораллов, органчиков, мягких и голубых кораллов, а также в губках, медузах, ресничных червях, моллюсках и даже в теле одноклеточных животных — простейших (инфузориях, радиоляриях и морских раковинных корненожках). Дж. Мак-Лауглин и П. Заль (J. McLaughlin und P. Zahl, 1966) содержали в культуре и исследовали строение симбиодиниумов, полученных от разных животных и из разных регионов Мирового океана. Они пришли к выводу, что все эти симбионты относятся к видам—космополитам.
Отдельная клетка симбиодиниума имеет вид округлого тельца диаметром около 0,005 мм. Снаружи она одета тонкой оболочкой, а внутри нее хорошо заметны ядро, желтоватый хлоропласт и зерна запасных питательных веществ (скорее всего, крахмала). Количество симбионтов в теле коралла невероятно велико: на каждый квадратный сантиметр поверхности полипа приходится около миллиона водорослевых клеток.
Размножается симбиодиниум делением надвое, а периодически клетка распадается на множество спор. Последние лишены оболочки и снабжены парой жгутиков, придающих им подвижность, за что они получили название зооспор. Переместившись на новое место, зооспора теряет жгутики, начинает расти и превращается в типичную клетку симбиодиниума. Половой процесс у этой водоросли неизвестен.
С помощью зооспор симбиодиниум заселяет энтодерму молодых выпочковываюшихся полипов. Посредством деления число клеток симбионта в тканях хозяина постоянно увеличивается. Когда наступает период полового размножения кораллов, зооспоры проникают в развивающиеся яйцеклетки. Плавающая личинка коралла (планула), которая едва достигает в длину 1 мм, несет в энтодерме около 7500 водорослевых клеток.
Способ чередования поколений у кишечнополостных называют метагенезом, а животных с этим типом жизненного цикла— мета-генетическими. Для метагенеза характерно правильное чередование полового поколения с поколением, размножающимся вегетативно, путем деления или почкования.
Но вернемся к типичному метагенезу и проследим за свободноплавающими медузами. Их образ жизни резко отличается от того, который ведут полипы. Медузы— подвижные животные, и потому у них вырабатываются приспособления для движения и ориентировки в пространстве. В первую очередь это касается развития зонтика и мускулатуры, а также органов чувств. У отдельных видов эти приспособления развиты настолько хорошо, что медузы могут далеко уплывать от тех мест, где они начали свою самостоятельную жизнь, отделившись от полипа. Часто они попадают в такие места, где нет подходящих условий для развития полипоидного поколения, например в открытое море, над большими глубинами. Планулы таких медуз, не найдя подходящего места для прикрепления, погибают. Но так обстоит дело далеко не со всеми видами гидроидных и сцифоидных медуз. Часть из них приспособилась к жизни именно в поверхностных слоях воды в открытом море, но при этом изменился и их жизненный цикл.
У гидромедузы бугенвиллии (Bougainvillia platygaster) яйца не выбрасываются в воду, а развиваются в половой железе самки, здесь же происходит и образование маленьких полипчиков, которые выпочковывают новых медуз. Случай этот не единственный, развитие полипов прямо на половой железе медузы характерно также и для одного видакампанулярии (Campanularia maccrady), и для некоторых других медуз. У этих видов полипоидное поколение хотя и имеется, но оно перестало быть донным и не образует больших колоний. У некоторых видов подавление полипоидной стадии пошло еще дальше и полипы вовсе исчезли из жизненного цикла. Из планулы таких медуз возникают не полипы, а такие же медузы. Так протекает жизненный цикл большой группы гидроидных медуз—трахилид, а из сцифоидных у видов рода пелагия (Pelagia), живущих в открытом море.
Как видно из приведенных примеров, метагенетический жизненный цикл весьма пластичен. Пока он является выгодным для процветания вида, он сохраняется, но как только условия меняются и одно из поколений получает некоторое преимущество, другое поколение подавляется и может совсем исчезнуть.
Источник: studopedia.su